

Filling the Silurian gap of solutan echinoderms with the description of new species of Dehmicystis from Spain
SAMUEL ZAMORA and JUAN CARLOS GUTIÉRREZ-MARCO
Zamora, S. and Gutiérrez-Marco, J.C. 2023. Filling the Silurian gap of solutan echinoderms with the description of new species of Dehmicystis from Spain. Acta Palaeontologica Polonica 68 (2): 185–192.
Solutans were among the most enigmatic pre-radial and asymmetric echinoderms. A new species Dehmicystis ariasi sp. nov. is described from the upper part of the Llagarinos Formation, lower Ludlow (Silurian) of Northwest Spain. This is the first solutan formally described from Iberia and the first from the Silurian worldwide. Dehmicystis was previously known based on a small number of poorly preserved specimens from the Emsian, Lower Devonian Hunsrück Slate of Germany. New observations based on the newly studied material suggest that Dehmicystis displayed a feeding appendage facing towards the substrate, and the periproct on the opposite face of the theca. Comparisons with other solutans and new data suggest that Dehmicystis was a detritus feeder that moved over the substrate capturing organic particles from the sediment with a single feeding arm.
Key words: Echinodermata, Soluta, pre-radial, asymmetric, Rhuddanian, Silurian, Spain.
Samuel Zamora [s.zamora@igme.es; ORCID: https://orcid.org/0000-0002-3917-4628 ], Instituto Geológico y Minero de España (IGME-CSIC), Residencia CSIC, Campus Aula Dei, Av. Montañana 1005, E-50059 Zaragoza, Spain.
Juan Carlos Gutiérrez-Marco [jcgrapto@ucm.es; ORCID: https://orcid.org/0000-0003-4213-6144 ], Instituto de Geociencias (CSIC, UCM) and Área de Paleontología, Departamento GEODESPAL, Facultad de Ciencias Geológicas, José Antonio Novais 12, E-28040 Madrid, Spain.
Received 13 January 2023, accepted 17 April 2023, available online 9 May 2023.
Copyright © 2023 S. Zamora and J.C. Gutiérrez-Marco. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Solutans are one of the most enigmatic, exclusively Paleozoic, group of fossil echinoderms whose phylogenetic position is controversial. They had a single ambulacrum and lacked the typical five-fold symmetry of modern echinoderms. Due to their bizarre morphology, the taxonomic position of the group has changed over time since they were first described in the mid-19th century (Noailles et al. 2014 and references therein). They were originally considered as a group of pelmatozoans related with diploporans, eocrinoids, and stylophorans (Bather 1899). Jaekel (1901) considered them as a group of carpoids within pelmatozoans, and grouped them with the asymmetric rhipidocystid eocrinoids. Gill and Caster (1960) were the first to remove rhipidocystids (that they assigned to the subclass Homostelea) from the “order” Soluta providing a better definition of the group. With the publication of the Treatise on Invertebrate Paleontology, Caster (1968) recognized the status of solutans (then included within homoiosteleans) (see further information about the early history of solutans in Lefebvre et al. 2012 and Noailles et al. 2014). Recent contributions consider solutans as a separate class of echinoderms (Lefebvre et al. 2012; Noailles et al. 2014; Nohejlová and Lefebvre 2022), with major disagreements over whether they represent preradial echinoderms before the development of the five-fold symmetry (Smith 2005, 2008; Zamora and Rahman 2014; Rozhnov 2022) or as a distinct group of blastozoans (David et al. 2000; Sprinkle and Guensburg 2004; Noailles et al. 2014).
Solutans ranged from the middle Drumian (Cambrian)to the Emsian (Lower Devonian) (Lefebvre and Lerosey-Aubril 2018). Diversity rapidly increased in the Cambrian Series 3, dropped by the Furongian and started to increase again by the Ordovician (Zamora et al. 2013; Lefebvre and Lerosey-Aubril 2018). No solutan taxa have been described so far from Silurian strata, suggesting that this class was very likely severely impacted by the Hirnantian mass-extinction (Domínguez et al. 2002; Nohejlová and Lefebvre 2022). However, here we fill the Silurian gap of solutans with the description of Dehmicystis ariasi sp. nov. from the ?Gorstian (lower Ludlow, Silurian) of Northwest Spain. This is the first formally described solutan from Iberia, although the Upper Ordovician from Central Spain and Iberian Chains has also provided incomplete specimens of Dendrocystites (Gil Cid et al. 1996; Zamora et al. 2014). This unquestionable record of a Silurian solutan has only been preceded by possible remains of the group in the Wenlock of Wales, which however, have not yet been described (Lefebvre in Nohejlová and Lefebvre 2022: 524). The new material from Spain also helps elucidating the posture of Dehmicystis with respect to the substrate, and provides evidence of feeding behavior in flattened solutans that have the feeding appendage and the periproct on opposite thecal surfaces.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: http://zoobank.org/urn:lsid:zoobank.org:pub:FE359270-26C4-419A- AA05-6F68C88A3173.
Institutional abbreviations.—MGM, Museo Geominero, Instituto Geológico y Minero de España-CSIC, Madrid, Spain; SMF, Naturmuseum Senckenberg, Germany.
Geological setting
Upper Silurian strata from the NW of the Iberian Peninsula crop out in the boundary between the West Asturian-Leonese and the Central Iberian zones of the Iberian Massif (Robardet and Gutiérrez-Marco 2002), where they have provided a diverse shelly fauna with Bohemian affinities including trilobites, crinoids and bivalves (Rábano et al. 1993; Gutiérrez-Marco et al. 2001). Echinoderms are rare and only consist of a few remains of primitive planktonic scyphocrinitid crinoids, being restricted to the Siluro-Devonian formations of the Courel-Peñalba and Sil synclines in the northern Central Iberian Zone (Gutiérrez-Marco et al. 2001). According to the map of Apalategui Isasa et al. (1981), the single specimen of Dehmicystis ariasi sp. nov. described here comes from a Silurian outcrop located in the northern flank of the Sil Syncline near its eastern termination, right below the so-called “cuarcitas azules” (“blue quartzites” = Yeres Quartzite of Zeitz and Nollau 1984). Corresponding beds, containing crinoids, were interpreted by Apalategui Isasa et al. (1981: 190) as probably belonging to the Silurian–Devonian transition, but according to a modern stratigraphical reappraisal for the Silurian of this syncline (Gutiérrez-Marco and Rábano 1997; Gutiérrez-Marco et al. 2001) they can be correlated with Ludlow strata, developed in a slightly chloritoid shaly facies, intercalated at the upper part of the Llagarinos Formation. The latter unit consists of ca. 150‒180 m of graptolite black shale ranging from the Spirograptus guerichi to the Neodiversograptus nilssoni graptolite zones (Telychian [upper Llandovery] to Gorstian [lower Ludlow]; Gutiérrez-Marco et al. 2001 and references therein). In the periclinal termination of the Sil Syncline, the base of the Llagarinos Formation is transgressive over diverse Ordovician formations such as the La Aquiana Limestone (upper Katian) in the northern flank, or the Losadilla Sandstone (Hirnantian) in the southern flank (Rodríguez Fernández et al. 2021). Additionally, the base of the Llagarinos Formation records an earlier development in the core of the adjacent Truchas Syncline, where it starts in the Rhuddanian Parakidograptus acuminatus Zone (Gutiérrez-Marco and Robardet 1991). Graptolites recorded close to the top of the Llagarinos Formation belong to the lower Gorstian (lower Ludlow) Neodiversograptus nilssoni Graptolite Zone, as for instance occur in the Yeres section placed 8 km east-southeast of the present locality (Gutiérrez-Marco and Rábano 1997). Consequently, an early Gorstian (sensu lato) (early Ludlow) may be the most probable age for the studied solutan due to its similar stratigraphic position with regard to the graptolite strata recorded below the Yeres Quartzite from this second nearby section. The position of the fossil locality of the studied specimen is indicated in Fig. 1, and this is situated ca. 960 m northeast of the chapel of Santo Tirso in O Castelo, a village situated to the west of the municipality of Rubiá, province of Ourense (Galicia Region, NW Spain). The point is also 340 m west of the peak of the Poleares hill (946 m high: also known as “cave of La Zorra” in older papers), and lies within an area cleared for a forest repopulation recently destroyed by fire, at the coordinates 42º27′22.5″N, 6º51′57.1″W.

Fig. 1. Localization of the study area in the northern part of the Central Iberian Zone, Spain. A. Sketch of the Iberian Massif showing the different structural and palaeogeographic zones with Neoproterozoic and Palaeozoic rocks. B. Schematic geological map, showing the position of the fossil locality (star) yielding the studied specimen. Base map modified from Villar Alonso et al. (2008) and González Menéndez et al. (2008), with Silurian subdivision adapted from Apalategui Isasa et al. (1981) and Abril Hurtado et al. (1982).
Systematic palaeontology
Phylum Echinodermata Bruguière, 1791 (ex Klein, 1734)
Class Soluta Jaekel, 1901
Remarks.—Soluta is herein considered as a class of echinoderms following Noailles et al. (2014). Previous workers such as David et al. (2000) suggested they possess a feeding appendage in the form of a brachiole, whereas others like Smith (2005) thought they had a true arm. We consider solutans as distinct from any other blastozoan group because of the lack of radial symmetry, distinct oral area and absence of brachioles, and treat them as a plesiomorphic group of early pre-radial echinoderms according with Smith (2005, 2008). Antero-posterior orientation follows Caster (1968) with the homoiostele at the posterior pole. Rahman and Lintz (2012) in the description of Dehmicystis considered the thecal surface bearing the ambulacrum (and associated mouth) as the oral surface, with the opposite face as the aboral surface (as suggested by Kolata et al. 1977). This does not necessarily imply direct homologies with other echinoderms. Descriptive terminology follows a recent review by Noailles et al. (2014).
Order Dendrocystitida Noailles, Lefebvre, and Kašička, 2014
Family Dendrocystitidae Bassler, 1938
Genus Dehmicystis Caster, 1968
Type species: Dendrocystites (Dendrocystoides?) globulus Dehm, 1934, Lower Devonian of western Germany (Fig. 2).
Emended diagnosis.—Solutan with a rounded flattened theca composed of polygonal plates. Medium-size feeding appendage on one of the thecal surfaces, with the hydropore and gonopore on same surface but separated one from each other. Periproct on the opposite face of the theca than the feeding appendage, consisting of a terminal anal pyramid, right to the homoiostele insertion. Large tetramerous proxistele and long dististele composed of paired ossicles.
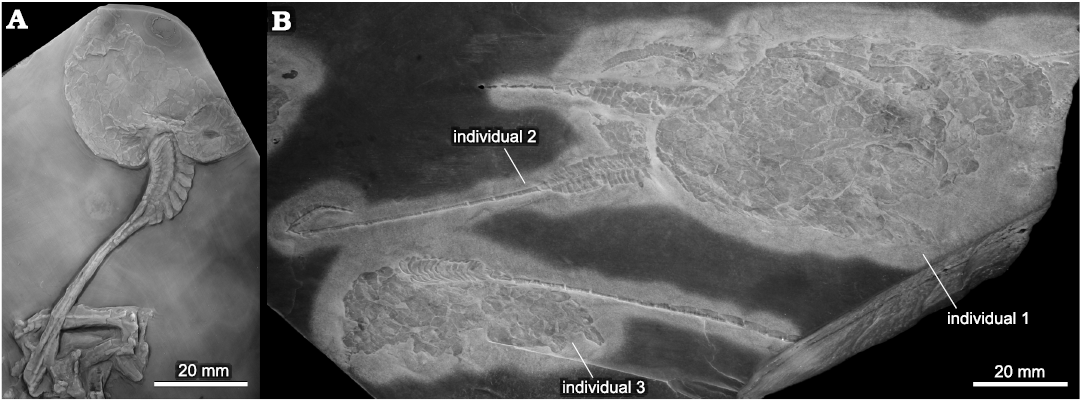
Fig. 2. Solutan echinoderm Dehmicystis globulus (Dehm, 1934) from the the Lower Devonian Hunsrück Slate, Germany. A. Holotype (SMF-HS 432), almost complete specimen showing the theca and posterior appendage. Note anus in on right side. B. SMF-HS 969, slab containing three individuals. Note that individual 1 has the anus on “left” position. Repository number assigned to a replica of the newly prepared slab. Photographs courtesy of Ulrich Jansen.
Remarks.—From all characters listed by Rahman and Lintz (2012) for the genus Dehmicystis the most important is the presence of a feeding appendage (arm) and anus “on the same face of the theca” (Rahman and Lintz 2012: 62). The position of the anal pyramid in Dehmicystis is problematic due in part to the poorly preservation of the material and also because the limited number of specimens available for study. Rahman and Lintz (2012) considered that both structures are on the same face of the theca based on a new studied specimen (Fig. 2B: individual 1). This contradicts the position of the anus in the holotype (Fig. 2A) which appears in the right side and opposite to the face where the feeding appendage is located. Specimens from Germany are extremely flattened and distorted resulting in the superimposition of both faces of the theca. Rahman and Lintz (2012) proposed three hypotheses explaining such position of the anus. The first of them considers this was a result of preservation, in which thecal plates have been displaced to partly reveal the opposing thecal surface where the anus is located, so that it appears to be on the opposite side of the theca. The second as a result of a “situs inversus” where structures located on one side of the theca are transposed to the opposite side, presumably as a result of developmental abnormalities. Finally, they choose a favored scenario in which flattening trajectories during burial affected the position of the anus. Our new material from Spain is less flattened than German specimens because different preservation in a coarser rock. The single specimen shows the arm and anus in the same face (Fig. 3A), but close observations of the anal pyramid (Fig. 3A4) suggest that it is preserved in a different plane than the feeding appendage. This supports the first scenario suggested by Rahman and Lintz (2012) in which some plates have been displaced revealing the opposite side of the theca where the anus was located. We thus consider that in Dehmicystis the arm and anus are on opposite faces. Other solutans (i.e., Dendrocystites Barrande, 1887, Coleicarpus Daley, 1996 and Maennilia Rozhnov and Jefferies, 1996) also have the arm and anus on opposite extremities; or have a terminal arm and the anus on one of the integuments (Girvanicystis) (Noailles et al. 2014). An undescribed Ordovician species from Hope Shale Formation in Shropshire shows the arm and stele on the same thecal face, and the anus on opposite face. A similar situation occurs in “Solutan indet., gen. et sp. nov.”, from the Middle Ordovician of Bohemia (Lefebvre et al. 2012). For contrast Dehmicystis has a terminal homoiostele, offset arm on one of the integuments and anus on the opposite side, right to the homoiostele insertion. Kolata et al. (1977) also consider that Belemnocystites wetherbyi Miller and Gurley, 1894, had the arm located some distance away from the margin. Minervaecystis (see Ubaghs 1970), Pahvanticystis (see Lefebvre and Lerosey-Aubril 2018) and Plasiacystis (see Lefebvre et al. 2012) also have an offset position of the arm.
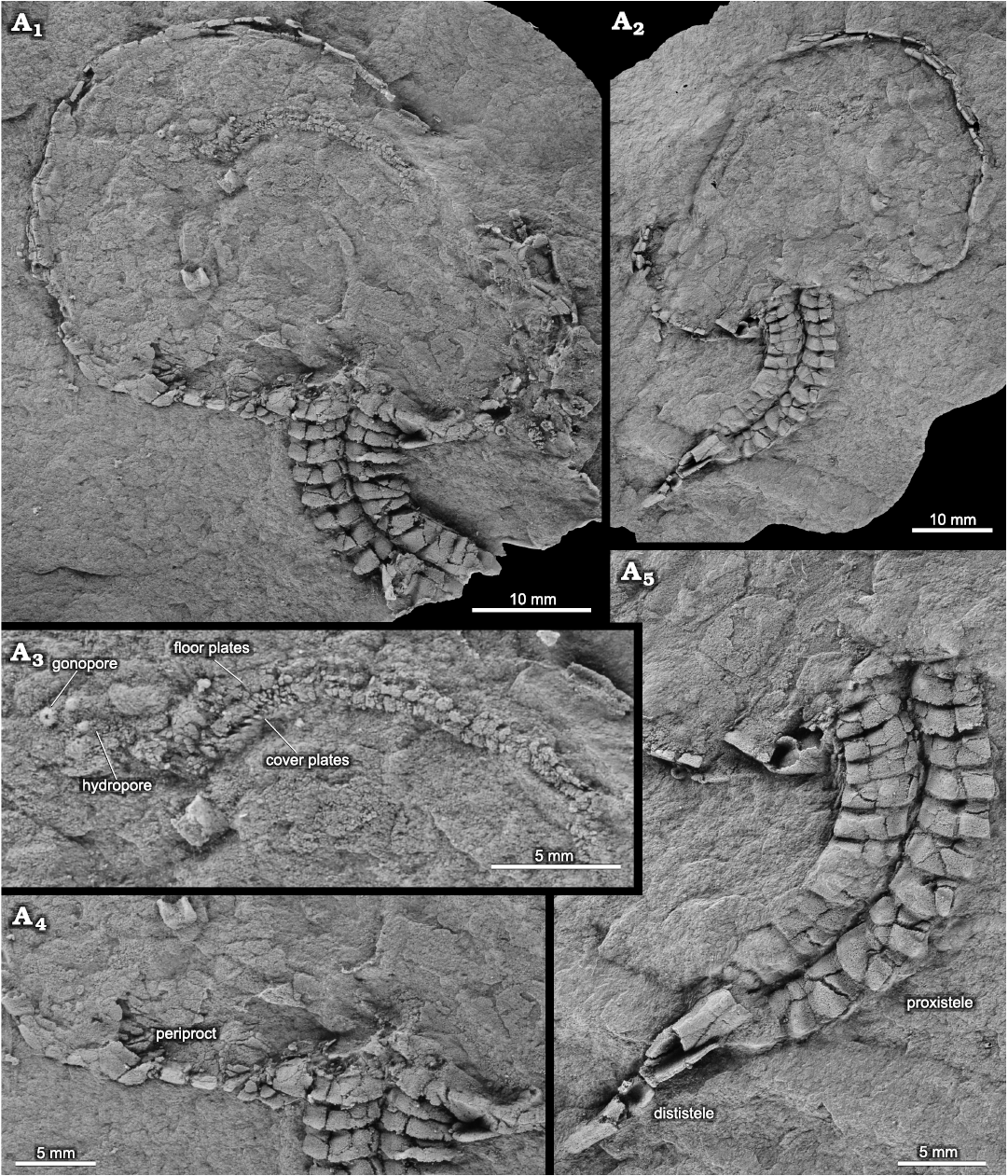
Fig. 3. Solutan echinoderm Dehmicystis ariasi sp. nov. from the Llagarinos Formation, Ludlow, Silurian, northern Central Iberian Zone, Spain. A. MGM-208S. A1, complete specimen in dorsal aspect preserving proxistele and feeding appendage above the body. A2, counterpart of same specimen. A3, detail of feeding appendage consisting on biserial floor plates and cover plates. Gonopore on single plate, and a possible hydropore close to the feeding appendage attachment. A4, detail of the periproct on right position. Note that the periproct is visible from the inside and was located on dorsal right part of the theca. A5, detail of the appendage preserving the complete proxistele and proximal dististele. Photographs are from latex casts whitened with ammonium chloride sublimated.
Dehmicystis ariasi sp. nov.
Figs. 3, 4.
Zoobank LCID: urn:lsid:zoobank.org:act:88604815-85BD-4E73-8FA4- 2381B337484C
Etymology: Dedicated to Francisco Arias Ferrero (Ponferrada, Spain) for collecting the only specimen known and making it available for study.
Holotype: MGM-208S-a (part) and MGM-208S-b (counterpart).
Type locality: 340 m west of Poleares high (946 m), about 960 m northeast of O Castelo village in the Rubiá municipality, Ourense Province, Galicia, NW Spain. GPS coordinates: 42º27′22.5″ N, 6º51′57.1″ W.
Type horizon: Shale bed with a slight chloritoid content in the upper part of the Llagarinos Formation. Bed of a probably Gorstian (early Ludlow) age by lateral correlation with a nearby locality.
Material.—Holotype only.
Diagnosis.—A species of Dehmicystis with a globular theca, proxistele equal in length to the theca. Undifferentiated mesistele.
Description.—Thecal shape rounded, large (45 mm in length, 30 mm in width), nearly bilaterally symmetric with the left side more developed than the right. Theca widening posteriorly with an invagination for the insertion of the homoiostele. Narrow peripheral ring as a result of compression rather than originally biological. Plates very thin, superimposed integuments (taphonomic), polygonal and relatively large. Boundaries between plates are poorly preserved. Feeding appendage off the apex, on an integument with attachment to the theca on left side; relatively large with wider rectangular basal plates (flooring plates) and thinner, cover plates. Feeding appendage thinning distally, with food groove oriented to the substrate (see interpretation below). Small gonopore on a circular tiny plate, associated with some rugose plates interpreted as hydropore. Both separated one another. Periproct large, on the opposite side of the theca, close to the appendage insertion but on the right position. Proxistele very large, consisting of eleven tetramerous rings, two on each side. Posterior side of each ring with a depressed side for the articulation of adjacent rows distally. Distal stele only preserved proximally, composed of two dimeres. Theca slightly longer than the proxistele.
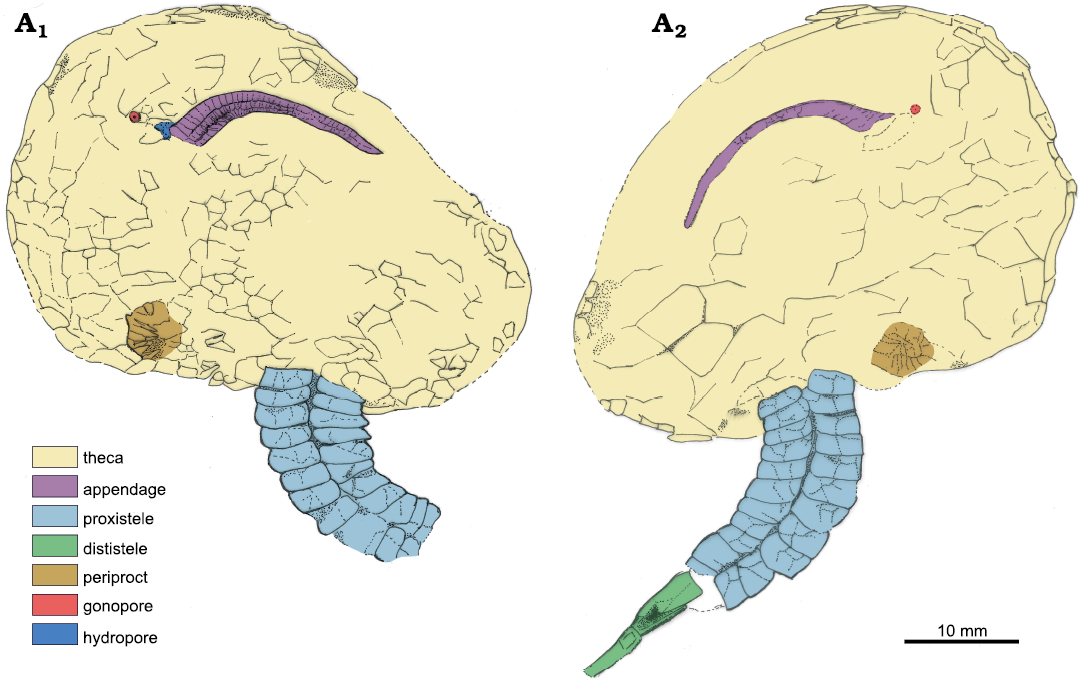
Fig. 4. Camera lucida drawings of solutan echinoderm Dehmicystis ariasi sp. nov. from the Llagarinos Formation, Ludlow, Silurian, northern Central Iberian Zone, Spain; with interpretation of main anatomical parts.
Remarks.—Comparison with the type species Dehmicystis globulus (Dehm, 1934) from Germany (Fig. 2), poses several problems; specially because German specimens are flattened and tectonically distorted, and also because none of the known specimens is totally complete (Fig. 2). Holotype and individual 1 from Rahman and Linz (2012) have the anterior part of the theca incomplete, individual 2 preserves only the homoiostele and individual 3 preserves only the homoiostele and posterior part of the theca. Rahman and Linz (2012) indicated that the dimensions of the holotype are 70 × 67 mm, but based on new observations, this is unlikely (see Table 1). The length of the holotype is unknown because incomplete preservation of the anterior part and its width is about 40 mm. So dimensions of the holotype are comparable to D. ariasi. The other individuals seem relatively larger. Shape of the theca between both species seems different, being more elongate in the German taxon, but this difference can be also due to the strong tectonic distortion affecting the Hunsrück material. Another important difference is the relation between thecal and proxistele lengths. Spanish material seems to have a longer proxistele which is almost equal than the length of the theca. For contrast D. globulus has a shorter proxistele. Mesistele is well preserved in the individual 2 of Rahman and Linz (2012), but it is absent in the Spanish material, where transition from the proxistele to the dististele is more abrupt. Further comparison with the type species is not possible due to incomplete preservation of material from both Spain and Germany, and different preservation styles being German specimens more compressed.
Table 1. Comparative measures (in mm) from Dehmicystis ariasi sp. nov. and Dehmicystis globulus (Dehm, 1934). L, length, W, width; (i), incomplete measure.
|
Specimen |
Repository number |
Thecal L |
Thecal W |
Proxistele |
Nº segments proxistele |
|
Dehmicystis ariasi sp. nov. (holotype) |
MGM-208S |
45 |
30 |
20 |
11 |
|
Dehmicystis globulus (holotype) |
SMF-HS 432 |
30 (i) |
40 |
15 |
?10–11 |
|
Dehmicystis globulus (individual 1) |
SMF-HS 969 |
70 (i) |
40 |
25 |
17 |
|
Dehmicystis globulus (individual 2) |
? |
? |
20 |
15 |
|
|
Dehmicystis globulus (individual 3) |
? |
50 |
18 |
12 |
Stratigraphic and geographic range.—Type horizon and locality only.
Concluding remarks
The earliest solutans in the Cambrian lived either permanently attached (i.e., Coleicarpus) (Ubaghs and Robison 1988; Daley 1996; Zamora et al. 2017) or were free living on muddy substrates (i.e., Castericystis, Pahvanticystis) (Ubaghs and Robison 1985; Lefebvre and Lerosey-Aubril 2018). In these earliest solutans (with the exception of Pahvanticystis), thecal faces are undifferentiated and the feeding appendage is positioned at the adoral pole of the theca on one of the margins, with the periproct on the opposite side, close to the homoiostele insertion (Ubaghs and Robison 1985). This is also the case for most Ordovician solutans like Dendrocystites and Girvanicystis (Noailles et al. 2014; Nohejlová and Lefebvre 2022); however, at least in Girvanicystis the periproct is positioned on one of the thecal faces (Daley 1992). In the case of Coleicarpus, which was permanently fixed on a hard substrate, the most likely interpretation is that it was a suspension feeder (Daley 1996), like some blastozoans. Most free living solutans were probably detritus feeders (Daley 1992; Noailles et al. 2014 and references therein) living on the sediment-water interface, as were other analogous early echinoderms like Dibrachicystis purujoensis (Zamora and Smith 2012) and pleurocystitids (Gorzelak and Zamora 2016). The situation is more complicated in solutes that have the periproct on one thecal surface and the feeding appendage away from the margin, on the opposite surface. Examples include “Solutan indet., gen. et sp. nov.”, from the Middle Ordovician of Bohemia (Lefebvre et al. 2012), an undescribed solutan from the Ordovician Hope Shale Formation in Shropshire, UK (SZ personal observation, 2004), Belemnocystites (Kolata et al. 1977) and species of Dehmicystis. Undescribed solutan from Hope Shale Formation is critic in such respect. Its theca is delimited by a narrow marginal frame and comprises two morphologically distinct surfaces: one is almost flat and hosts the insertions of the two appendages (free ambulacrum and homoiostele), whereas the opposite thecal surface is domed (convex) and bears the periproct (Bertrand Lefebvre, personal communication 2023). Comparison with some other free living, epibenthic echinoderms (e.g., cinctans, pleurocystitids, edrioasteroids) suggests the flat surface was in contact with the substrate, whereas the convex one was facing upwards; this is also supported by the insertion of the stele in the flat surface, in the same face than the feeding arm. Rahman and Lintz (2012) interpreted Dehmicystis as having the feeding appendage and periproct on same surface based on very flattened specimens, but the specimen described herein and comparison with other solutans suggest that Dehmicystis is better interpreted as having feeding appendage and periproct on opposite faces (see discussion above). If correctly interpreted, Dehmicystis probably laid on the substrate with the periproct on the upper surface and the feeding appendage directed to the substrate. Under such interpretation Dehmicystis was probably a detritus feeder capturing organic particles from the muddy substrate.
Acknowledgements
This paper would be not possible without the help of Francisco Arias Ferrero (Ponferrada, Spain) who collected the specimen and made it available for study. We appreciate comments on such enigmatic animals from two referees Imran A. Rahman (Natural History Museum, London, UK) and Bertrand Lefebvre (CNRS-University of Lyon, France) that greatly improved the resulting manuscript. The former also provided as excellent photographs of D. globulus reproduced in Fig. 2 and originally taken by Ulrich Jansen (Senckenberg Naturmuseum, Frankfurt, Germany). This work was supported by the Spanish Ministry of Science and Innovation (PID2021-125585NB-100), co-financed by the European Regional Development Fund, project “Aragosaurus: Recursos Geológicos y Paleoambientales” (E18_17R) funded by the Government of Aragón and grant AECEX2021 “Severo Ochoa extraordinary grants for excellence IGME-CSIC”.
References
Abril Hurtado, J., Pliego Dones, D., and Rubio Navas, J. 1982. Mapa y Memoria Explicativa de la Hoja nº 191 (Silván) del Mapa Geológico de España a escala 1:50.000 (2ª serie MAGNA), 56. Instituto Geológico y Minero de España, Madrid.
Apalategui Isasa, O., Abril Hurtado, J., and Rodríguez Fernández, L.R. 1981. Mapa y Memoria Explicativa de la Hoja nº 190 (Barco de Valdeorras) del Mapa Geológico de España a escala 1:50.000 (2ª serie MAGNA), 44. Instituto Geológico y Minero de España, Madrid.
Barrande, J. 1887. Système silurien du centre de la Bohême. 1ère Partie: Recherches paléontologiques. Volume 7. Classe des Echinodermes. Ordre des Cystidées. 233 pp. Gerhard, Leipzig.
Bassler, R.S. 1938. Pelmatozoa Palaeozoica (Generum et Genotyporum Index et Bibliographia). Fossilium Catalogus I Animalia 83: 1–194.
Bather, F.A. 1899. A phylogenetic classification of the Pelmatozoa. Report of the British Association for the Advancement of Science 1898: 916–923.
Bruguière, J.G. 1791. Tableau Encyclopédique et Méthodique des Trois Règnes de la Nature, contenant l’Helminthologie, ou les vers Infusoires, les vers Intestins, les vers Mollusques, etc., Volume 7. 180 pp. Panckoucke, Paris. Crossref
Caster, K.E. 1968. Homoiostela. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology. Part S. Echinodermata 1(2), S581–S627. Geological Society of America, New York and University of Kansas, Lawrence.
Daley, P.E.J. 1992. The anatomy of the solute Girvanicystis batheri (?Chordata) from the Upper Ordovician of Scotland and a new species of Girvanicystis from the Upper Ordovician of South Wales. Zoological Journal of the Linnean Society 105: 353–375. Crossref
Daley, P.E.J. 1996. The first solute which is attached as an adult: a Mid-Cambrian fossil from Utah with echinoderm and chordate affinities. Zoological Journal of the Linnean Society 117: 405–440. Crossref
David, B., Lefebvre, B., Mooi, R., and Parsley, R. 2000. Are homalozoans echinoderms? An answer from the extraxial-axial theory. Paleobiology 26: 529–555. Crossref
Dehm, R. 1934. Untersuchungen an Cystoideen des rheinischen Unterdevons. Sitzungsberichte der Mathematisch-Naturwissenschaftlichen Abteilung der Bayerischen Akademie der Wissenschaften zu München für 1934: 19–43.
Domínguez, P.D., Jefferies, R.P.S., and Gil Cid, M.D. 2002. An annotated check-list of genera and species of carpoids. Coloquios de Paleontología 53: 33–68.
Gil Cid, M.D., Domínguez Alonso, P., Silván Pobes, E., and Escribano Ródenas, M. 1996. Bohemiaecystis jefferiesi n. sp.; primer Cornuta para el Ordovícico español. Estudios Geológicos 52: 313–326. Crossref
Gill, E.D. and Caster, K.E. 1960. Carpoid echinoderms from the Silurian and Devonian of Australia. Bulletins of American Paleontology 41: 1–71.
González Menéndez, L., Heredia, N., and Marcos, A. 2008. Mapa Geológico Digital continuo E. 1: 50.000, Zona Asturoccidental-Leonesa (Zona-1100). In: GEODE Mapa Geológico Digital continuo de España. Accessed December 16, 2022 at http://info.igme.es/cartografiadigital/geologica/geodezona.aspx?Id=Z1100
Gorzelak, P. and Zamora, S. 2016. Understanding form and function of the stem in early flattened echinoderms (pleurocystitids) using a microstructural approach. PeerJ 4: e1820. Crossref
Gutiérrez-Marco, J.C. and Rábano, I. 1997. Los materiales del Ordovícico y Silúrico de la región limítrofe entre los dominios de Caurel-Peñalba (Zona Asturoccidental-leonesa) y Truchas (flanco norte del Antiforma del Ollo de Sapo: Zona Centroibérica). In: A. Grandal d’Anglade, J.C. Gutiérrez-Marco, and L. Santos Fidalgo (eds.), Comunicaciones XIII Jornadas de Paleontología y V Reunión Internacional PIGC 351, A Coruña, 298–313. Universidad de da Coruña, A Coruña.
Gutiérrez-Marco, J.C. and Robardet, M. 1991. Découverte de la zone à Parakidograptus acuminatus (base du Llandovery) dans le Silurien du Synclinorium de Truchas (Zone Asturo-léonaise, Nord-Ouest de l’Espagne): conséquences stratigraphiques et paléogéographiques au passage Ordovicien–Silurien. Comptes Rendus de l’Acadèmie des Sciences de Paris [IIa] 312: 729–734.
Gutiérrez-Marco, J.C., Sarmiento, G.N., Robardet, M., Rábano, I., and Vaněk, J. 2001. Upper Silurian fossils of Bohemian type from NW Spain and their palaeogeographical interest. Journal of the Czech Geological Society 46: 247–258.
Jaekel, O. 1901. Über Carpoideen; eine neue Klasse von Pelmatozoen. Zeitschrift der Deutschen geologischen Gesellschaft 52: 661–677.
Klein, J.T. 1734. Naturalis dispositio Echinodermatum. Accessit lucubratiuncula de Aculeis Echinorum Marinorum, cum spicilegio de Belemnitis. 278 pp. Schreiber, Gedani. Crossref
Kolata, D.R., Strimple, H.L., and Leverson, C.O. 1977. Revision of the Ordovician carpoid family Iowacystidae. Palaeontology 20: 529–557.
Lefebvre, B. and Lerosey-Aubril, R. 2018. Laurentian origin of solutan echinoderms: new evidence from the Guzhangian (Cambrian Series 3) Weeks Formation of Utah, USA. Geological Magazine 155: 1190–1204. Crossref
Lefebvre, B., Derstler, K., and Sumrall, C. D. 2012. A reinterpretation of the solutan Plasiacystis mobilis (Echinodermata) from the Middle Ordovician of Bohemia. In: A. Kroh and M. Reich (eds.), Echinoderm Research 2010. Proceedings of the Seventh European Conference on Echinoderms, Göttingen, Germany, 2–9 October 2010. Zoosymposia 7: 287–306. Crossref
Miller, S.A. and Gurley, W.F.E. 1894. New genera and species of Echinodermata. Bulletin of the Illinois State Museum of Natural History 5: 5–53. Crossref
Noailles, F., Lefebvre, B., and Kašička, L. 2014. A probable case of heterochrony in the solutan Dendrocystites Barrande, 1887 (Echinodermata: Blastozoa) from the Upper Ordovician of the Prague Basin (Czech Republic) and a revision of the family Dendrocystitidae Bassler, 1938. Bulletin of Geosciences 89: 451–476. Crossref
Nohejlová, M. and Lefebvre, B. 2022. Late Ordovician solutan echinoderms from the Western Tafilalt, Morocco. Geological Society, London, Special Publications 485: 523–539. Crossref
Rábano, I., Gutiérrez-Marco, J.C., and Robardet, M. 1993. Upper Silurian trilobites of Bohemian affinities from the West Asturian-Leonese Zone (NW Spain). Geobios 26: 361–376. Crossref
Rahman, I.A. and Lintz, H. 2012. Dehmicystis globulus, an enigmatic solute (Echinodermata) from the Lower Devonian Hunsrück Slate, Germany. Paläontologische Zeitschrift 86: 59–70. Crossref
Robardet, M. and Gutiérrez-Marco, J.C. 2002. Silurian. In: W. Gibbons and T. Moreno (eds.), The Geology of Spain, 51–66. The Geological Society, London. Crossref
Rodríguez Fernández, L.R., Toyos, J.M., Díez Montes, A., González Menéndez, L., Gallastegui, G., Heredia, N., Martín Parra, L.M., and Rubio Pascual, F.J. 2021. Mapa geológico de la Hoja nº 18 (Ponferrada). Mapa Geológico de España E. 1:200.000. Primera edición en línea. IGME, CSIC, Madrid.
Rozhnov, S.V. 2022. Solutans (Echinoderms): Evolution Frozen between Torsion and Pentaradiality. Paleontological Journal 56: 1306–1321. Crossref
Rozhnov, S.V. and Jefferies, R.P.S. 1996. A new stem-chordate solute from the middle Ordovician of Estonia. Geobios 29: 91–109. Crossref
Smith, A.B. 2005. The pre-radial history of echinoderms. Geological Journal 40: 255–280. Crossref
Smith, A.B. 2008. Deuterostomes in a twist: the origins of a radical new body plan. Evolution & Development 10: 493–503. Crossref
Sprinkle, J. and Guensburg, T.E. 2004. Crinozoan, blastozoan, echinozoan, asterozoan, and homalozoan echinoderms. In: B.D. Webby, F. Paris, M.L. Droser, and I.G. Percival (eds.), The Great Ordovician Biodiversification Event, 266–280. Columbia University Press, New York. Crossref
Ubaghs, G. 1970. Les échinodermes carpoïdes de l’Ordovicien inférieur de la Montagne Noire (France). 112 pp. Editions du CNRS, Paris.
Ubaghs, G. and Robison, R.A. 1985. A new homoiostelean and a new eocrinoid from the Middle Cambrian of Utah. University of Kansas Paleontological Contributions 115: 1–24.
Ubaghs, G. and Robison, R.A. 1988. Homalozoan echinoderms of the Wheeler Formation (Middle Cambrian) of western Utah. University of Kansas Paleontological Contributions 120: 1–17.
Villar Alonso P., Portero Urroz, G., González Cuadra P., García Crespo, J., Nieto García A. B., Rubio Pascual, F. J., Gómez Fernández, F., and Jiménez Benayas, S. 2008. Mapa Geológico Digital continuo E. 1: 50.000, Zona Centroibérica. Domino Ollo de Sapo (Zona-1300). In: GEODE Mapa Geológico Digital continuo de España. Accessed December 16th, 2022 at http://info.igme.es/cartografiadigital/geologica/geodezona.aspx?Id=Z1300
Zamora, S. and Rahman, I. A. 2014. Deciphering the early evolution of echinoderms with Cambrian fossils. Palaeontology 57: 1105–1119. Crossref
Zamora, S. and Smith, A. B. 2012. Cambrian stalked echinoderms show unexpected plasticity of arm construction. Proceedings of The Royal Society B 279: 293–298. Crossref
Zamora, S., Lefebvre, B., Álvaro, J.J., Clausen, S., Elicki, O., Fatka, O., Jell, P., Kouchinsky, A., Lin, J.-P., Nardin, E., Parsley, R., Rozhnov, S., Sprinkle, J., Sumrall, C.D., Vizcaïno, D., and Smith, A.B. 2013. Cambrian echinoderm diversity and palaeobiogeography. In: D.A.T. Harper and T. Servais (eds.), Early Palaeozoic Biogeography and Palaeogeography. Geological Society of London, Memoirs 38: 157–171. Crossref
Zamora, S., Colmenar, J., and Ausich, W.I. 2014. The echinoderm faunas from the Fombuena Formation (Upper Ordovician, Iberian Chains, Spain). In: R. Royo-Torres, F.J. Verdú, and L. Alcalá (coord.), XXX Jornadas de Paleontología de la Sociedad Española de Paleontología. Fundamental 24: 257–259.
Zamora, S., Deline, B., Álvaro, J.J., and Rahman, I.A. 2017. The Cambrian Substrate Revolution and the early evolution of attachment in suspension-feeding echinoderms. Earth-Science Reviews 171: 478–491. Crossref
Zeitz, U. and Nollau, G. 1984. Ordoviz und Silur im Sil-Synclinorium südlich Ponferrada (Provinz León), NW-Spanien. Zeitschrift der Deutschen Geologischen Gessellschaft 135: 211–222. Crossref
Acta Palaeontol. Pol. 68 (2): 185–192, 2023
https://doi.org/10.4202/app.01054.2023