

New Pleistocene bird fossils in Taiwan reveal unexpected seabirds in East Asia
SIAO-MAN WU, TREVOR H. WORTHY, CHIH-KAI CHUANG, and CHIEN-HSIANG LIN
Wu, S.-M., Worthy, T.H., Chuang, C.-K., and Lin, C.-H. 2023. New Pleistocene bird fossils in Taiwan reveal unexpected seabirds in East Asia. Acta Palaeontologica Polonica 68 (4): 613–624.
The island of Taiwan, with its diverse microclimates and key position on the East Asian-Australasian Flyway, attracts numerous bird enthusiasts due to its diverse avian fauna. Nevertheless, due to the scarcity of fossil records, there is a significant knowledge gap between modern and ancient avifaunas in Taiwan. Currently, there is only a single described Pleistocene fossil; it is attributed to Phasianidae. To address this gap, this study describes two new bird fossils, a left humerus and a left tibiotarsus, and discusses them in detail herein. The fossils were collected from the Liuchungchi Formation (Early Pleistocene, 1.95–1.35 Ma) in Niubu, Chiayi, southwestern Taiwan, which represents a neritic environment. The fossils are identified as from species of Gaviidae (loons), with the humerus belonging to an undetermined species of Gavia and the tibiotarsus to Gavia stellata. Loons are seabirds that are primarily distributed in high- and middle latitudes of the Northern Hemisphere. In addition, these birds are extremely rare in modern Taiwan: records are scarce and most are limited to northern and northeastern Taiwan since the 1860s, indicating that the modern Gavia birds only occasionally visit Taiwan. All known Pleistocene fossils of species of Gavia from the northern West Pacific come from Japan. The Taiwan fossils of Gavia provide valuable bird evolutionary and paleobiogeographic information for the subtropical West Pacific and may imply the presence of a distinct avifauna in the region during the Early Pleistocene.
Key words: Aves, Gavia, loon, seabird, Early Pleistocene, Taiwan.
Siao-Man Wu [smallmmmm@gmail.com; ORCID: https://orcid.org/0009-0004-3702-1996 ], Chih-Kai Chuang [ntnupast@gmail.com; ORCID: https://orcid.org/0009-0001-1289-9442 ] and Chien-Hsiang Lin [chlin.otolith@gmail.com; ORCID: https://orcid.org/ 0000-0002-9843-9729 ] (corresponding author), Biodiversity Research Center, Academia Sinica, Taipei, 11529, Taiwan.
Trevor H. Worthy [trevor.worthy@flinders.edu.au; ORCID: https://orcid.org/0000-0001-7047-4680 ], Palaeontology Group, College of Science and Engineering, Flinders University, Adelaide, 5001, Australia.
Received 18 July 2023, accepted 10 November 2023, available online 15 December 2023.
Copyright © 2023 S.-M. Wu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Seabirds inhabit marine environments and forage for fish, squid and krill as their primary diet. They are diverse, with some species also distributed to freshwater environments (BirdLife International 2023; Mayr 2022; Tyrberg 1998). They play a vital role in indicating the health of marine habitats due to their high trophic level and long-distance flights to locate ideal foraging habitats (Croxall et al. 1999, 2012; Piatt et al. 2007; Parsons et al. 2008). Therefore, oceanographic dynamics at geological scales have driven the evolution and distribution of seabirds (Warheit 1992; Block et al. 2011; Clay et al. 2017; Drummond et al. 2021). The Pacific Ocean and surrounding coasts have supported a high diversity of seabirds through the Cenozoic (BirdLife International 2023). Based on fossil records, seabirds appear to have evolved and diversified in the North Pacific region early in the Eocene and Oligocene, and continued to thrive during the Miocene and Pliocene. Examples include Charadriiformes (Miller 1931), Gaviiformes (Olson and Rasmussen 2001), Procellariiformes (Mayr 2015; Mayr and Goedert 2017), and plotopterids, flightless wing-propelled divers in Suliformes (Goedert 1988; Olson and Hasegawa 1996; Kimura et al. 1998; Goedert and Cornish 2002; Sakurai et al. 2008; Dyke et al. 2011; Mayr et al. 2015; Mayr and Goedert 2018, 2022; Ohashi and Hasegawa 2020). During the Pleistocene, the fossil records of seabirds become more diversified with modern lineages, but most are restricted to the Northeast Pacific (Miller 1914, 1925, 1930; Howard 1936, 1949, 1958; Guthrie 1992, 2005; Collins et al. 2018). In the Northwest Pacific, diverse Pleistocene seabird faunas have been reported from Japan (Hasegawa et al. 1988; Watanabe and Matsuoka 2015; Watanabe et al. 2016, 2018a, b, 2020; Matsuoka and Hasegawa 2018; Aotsuka et al. 2022), and a few examples come from Russia (Harington 1978; Tyrberg 1998).
Taiwan is a subtropical island (~20° N) in the Northwest Pacific where many seabirds can be found (Xiao and Li 2022). Though well-known for its diverse modern avifauna, little is known about its ancient birds due to the extreme rarity of bird fossils. The Western Foothills of Taiwan has several fossil localities that sample mainly marine sedimentary environments, making it challenging to discover bird fossils (Lin et al. 2021). The only published bird fossil record to date is a phasianid tarsometatarsus from the Middle Pleistocene of southern Taiwan (Tsai and Mayr 2021).
Recently, two previously undescribed Early Pleistocene bird fossils from southwest Taiwan were found in the Chiayi Municipal Museum. This paper aims to describe these specimens in detail by comparing their morphology with modern specimens and investigate their correspondence with the whole avifauna in the Pleistocene of the Northwest Pacific.
Institutional abbreviations.—CMM, Chiayi Municipal Museum, Chiayi, Taiwan; NHMUK, Natural History Museum, Tring, UK; YIO, Yamashina Institute for Ornithology, Abiko, Chiba, Japan.
Geological setting
The fossils were collected from Niubu, Chiayi, southwest Taiwan (Fig. 1). The Niubu area is renowned for its rich deposits of marine fossils (Hu 1989; Tao 1993; Hu and Tao 1996, 2004; Xue 2004; Buckeridge et al. 2018; Lin et al. 2018, 2022); some terrestrial mammalian fossils and fragments of carbonized woods are also known (Xue 2004). The locality is along the Bazhang River where the Liuchungchi Formation is widely exposed (Lin et al. 2022: figs. 1, 2). The Liuchungchi Formation is composed of light gray muddy siltstone intercalated with dark gray shale or sandy shale, and the depositional environment sequence reflects a shoreface environment from offshore transition zone to inner offshore (Chang 2008; Chen 2016). Because the exact stratigraphic localities of the fossil specimens (Fig. 2) are uncertain, we conducted a pilot nannofossil analysis to constrain the age of the two fossils. Approximately 20 mg of sediments on the surfaces of the fossils and inside each of the shafts were removed. Then two sediment subsamples for each bird fossil were prepared and investigated for nannofossils using polarized light microscopy (Zeiss Axioscope Microscope), following the procedures of Gartner (1969). However, no nannofossils were found in any of the samples. A previous calcareous nannofossil biostratigraphic analysis of the southern Liuchungchi Formation identified the first appearance datum (FAD) Gephyrocapsa oceanica (~1.7 Ma) and the last appearance datum (LAD) Discoaster brouweri (~1.95 Ma) events (Chi 1980; Huang and Ting 1981), but a biostratigraphic study of the northern Liuchungchi Formation, where Niubu is located, has not been conducted. Chen et al. (2011) studied both paleomagnetic and sequence stratigraphic data of the whole Liuchungchi Formation and constrained the age of the formation to 1.90–1.35 Ma. Therefore, we use the gross age 1.95–1.35 Ma for our fossils (Fig. 3).
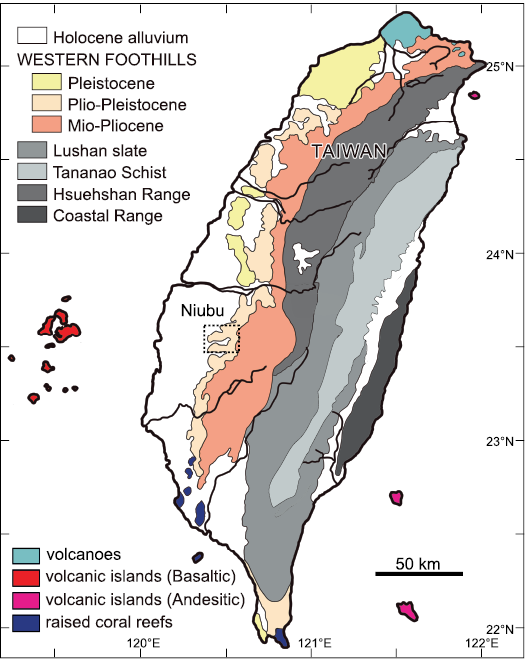
Fig. 1. Geological setting of Taiwan area and Niubu area; the locality of the two bird fossils is marked with rectangle (modified after Lin et al. 2022).
Material and methods
The fossils were collected during the 1980s–2000s by the private collector Wen-Ji Xue, who then donated them to the Chiayi Municipal Museum (CMM), Chiayi, Taiwan. The fossils have been cataloged under CMM 2123 and CMM 2124. The two bird fossils are the well-preserved distal ends of a left humerus and a left tibiotarsus, respectively (Fig. 2).
To uncover the bone morphology covered by the sediments, computed tomography (CT) was applied on the bird fossils using SkyScan 1276 (Bruker microCT, Kontich, Belgium) at the Taiwan Mouse Clinic, Academia Sinica and Taiwan Animal Consortium, Taiwan. The voltage, current, and scan resolution of X-ray source were set to 100 kV, 200 μA, and 20.5 μm, respectively. The filters Al and Cu were used. After scanning was completed, the image processing algorithms following Wang et al. (2015) were applied to the 3D raw images to build the 3D models. The widths and depths of the bird fossils were obtained by using calipers while avoiding broken surfaces. The measurements of CMM 2123 include: (i) the dorsoventral length of the shaft from the cranial view as the shaft width; (ii) the craniocaudal length from the dorsal view of the shaft as the shaft depth; (iii) the length between the ventrodistal face of the condylus ventralis and the dorsodistal face of the condylus dorsalis from the cranial view as the distal width; (iv) the maximal craniocaudal length of the condylus dorsalis from the dorsal view as the distal depth. The measurements of CMM 2124 include: (i) the mediolateral length of the shaft from the cranial view as the shaft width; (ii) the craniocaudal length from the lateral view of the shaft as the shaft depth; (iii) the length between the medial face of the condylus medialis and the lateral face of the condylus lateralis from the cranial view as the distal width; (iv) the maximal craniocaudal length of the condylus lateralis from the lateral view as the distal depth.
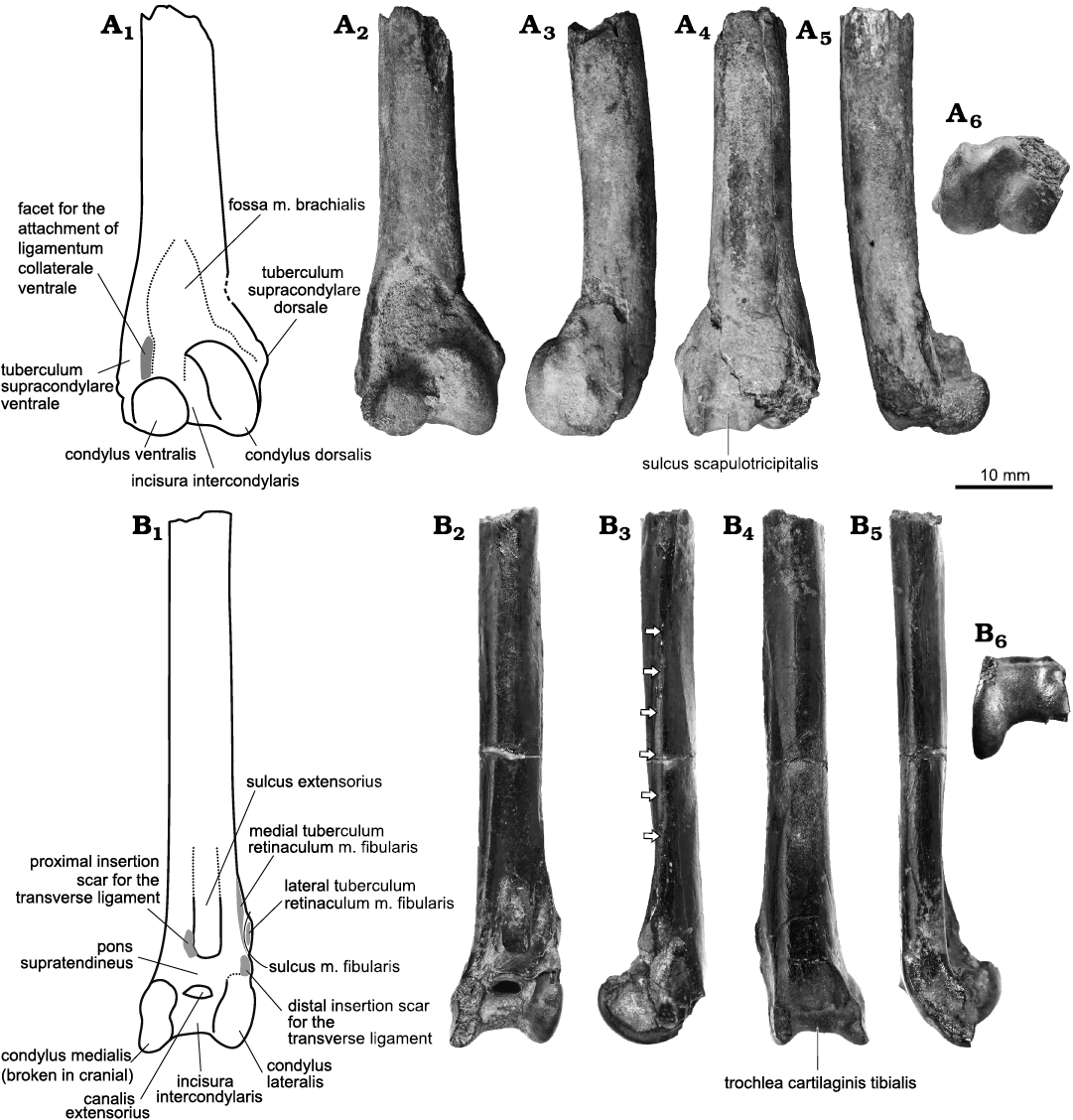
Fig. 2. The fossils of gaviid seabirds from Niubu, Chiayi, Taiwan, Liuchungchi Formation, Lower Pleistocene. A. Gavia sp., CMM 2123, distal left humerus in cranial (A1, A2), dorsal (A3), caudal (A4), ventral (A5), and distal (A6) views. B. Gavia stellata (Pontoppidan, 1763), CMM 2124, distal left tibiotarsus in cranial (B1, B2), lateral (B3), caudal (B4), medial (B5), and distal (B6) views. Arrows highlight the articular scar for the distal fibula.
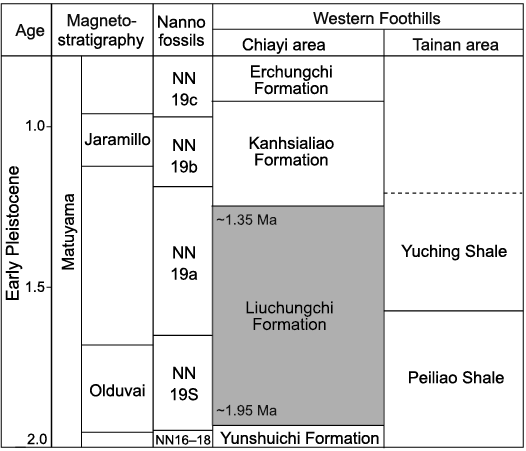
Fig. 3. Stratigraphy of the Western Foothills of southern Taiwan (modified after Chen 2016; Lin et al. 2021). Grey block highlights the Liuchungchi Formation, wherein the bird fossils were found. Only the Early Pleistocene of southern Taiwan is shown.
To compare the size variations, the measurements of modern specimens preserved in the Yamashina Institute for Ornithology, Abiko, Chiba, Japan (YIO) were obtained using the same criteria as those of the fossil specimens (see above). For Gavia immer (Brünnich, 1764), a species not from the West Pacific and not found in YIO, the measurements were obtained from the images of the specimens from the Natural History Museum, Tring, UK (photos courtesy of Junya Watanabe) with a scale bar by using Adobe Photoshop (v. 24.1.1) and its Ruler Tool. Though it was not ideal to obtain measurements from photos, the scale bar was located close to the bone to minimize the measurement error and the obtained data are sufficient to show that the fossils were considerably smaller than the specimens in the imaged sample. The anatomical nomenclature follows Baumel and Witmer (1993).
Systematic palaeontology
Aves Linnaeus, 1758
Order Gaviiformes Wetmore and Miller, 1926
Family Gaviidae Allen, 1897 (Gray, 1840)
Genus Gavia Forster, 1788
Type species: Gavia immer (Brünnich, 1764); Recent, North America.
Gavia sp.
Figs. 2A, 4A.
Material.—CMM 2123 left humerus, Bazhang River, Niubu area, Taiwan; Liuchungchi Formation (Fig. 3); 1.95–1.35 Ma.
Measurements.—See Table 1.
Table 1. Measurements (in mm) of the fossils and modern Gaviidae. Number in the parentheses denotes sample size. The order of modern Gaviidae follows the taxonomy established by Sprengelmeyer (2014). The measurements of Gavia immer were obtained from photos taken by Junya Watanabe. Abbreviations: D, depth; W, width.
|
Measurement |
CMM 2123 |
CMM 2124 |
Gavia stellata |
Gavia arctica |
Gavia pacifica |
Gavia adamsii |
Gavia immer |
|||||
|
Range |
Mean |
Range |
Mean |
Range |
Mean |
Range |
Mean |
Range |
Mean |
|||
|
Humerus |
||||||||||||
|
shaft W |
8.5 |
NA |
6.9–7.9 |
7.4 (4) |
8.1–9.2 |
8.6 (7) |
7.3–9.4 |
8.3 (14) |
11.2–11.2 |
11.2 (1) |
8.8–11.6 |
10.5 (6) |
|
shaft D |
7.9 |
NA |
5.3–6.8 |
5.9 (4) |
5.7–7.4 |
6.4 (7) |
5.6–7.8 |
6.4 (14) |
8.0–8.0 |
8.0 (1) |
7.2–8.7 |
8.3 (6) |
|
distal W |
13.8 |
NA |
11.0–12.0 |
11.6 (4) |
12.3–13.9 |
13.1 (7) |
11.5–13.4 |
12.6 (14) |
17.0–17.0 |
17.0 (1) |
15.0–18.0 |
16.7 (6) |
|
distal D |
11.0 |
NA |
8.2–9.7 |
8.9 (4) |
9.8–11.1 |
10.5 (7) |
9.3–10.6 |
10.0 (14) |
13.9–13.9 |
13.9 (1) |
11.8–15.0 |
13.6 (6) |
|
shaft W/distal W |
0.6 |
NA |
0.6–0.7 |
0.6 (4) |
0.6–0.7 |
0.7 (7) |
0.6–0.7 |
0.7 (14) |
0.7–0.7 |
0.7 (1) |
0.6–0.7 |
0.6 (6) |
|
Tibiotarsus |
||||||||||||
|
shaft W |
NA |
6.9 |
7.9–9.1 |
8.5 (4) |
7.0–8.3 |
7.6 (7) |
6.6–9.1 |
7.5 (11) |
9.4–9.5 |
9.4 (2) |
8.9–11.1 |
9.9 (6) |
|
shaft D |
NA |
4.8 |
4.6–5.5 |
5.2 (4) |
4.4–5.8 |
5.2 (7) |
4.6–6.6 |
5.2 (11) |
6.4–7.4 |
6.9 (2) |
6.0–6.5 |
6.2 (6) |
|
distal W |
NA |
12.0 |
10.6–12.3 |
11.6 (4) |
12.1–13.1 |
12.7 (7) |
11.8–13.8 |
12.3 (11) |
15.3–15.8 |
15.5 (2) |
14.0–17.9 |
16.4 (6) |
|
distal D |
NA |
12.0 |
11.6–13.0 |
12.2 (4) |
12.5–14.1 |
13.1 (7) |
11.2–13.8 |
12.4 (11) |
16.4–16.7 |
16.6 (2) |
14.2–17.3 |
15.5 (6) |
|
shaft W/distal W |
NA |
0.5 |
0.7–0.8 |
0.7 (4) |
0.5–0.6 |
0.6 (7) |
0.6–0.7 |
0.6 (11) |
0.6–0.6 |
0.6 (2) |
0.6–0.6 |
0.6 (6) |
Description.—The shaft of the distal left humerus CMM 2123 is stout and rounded in cross section, and is notched craniodorsally proximal to the tuberculum supracondylare dorsale. Distally, the shaft forms an angle of 150° with respect to the proximal side of the tuberculum supracondylare dorsale. The tuberculum supracondylare dorsale is dorsally prominent and proximodistally overlaps the condylus dorsalis such that the apex is located distal to the most proximal part of the condyle; the sharp and elevated apex extends smoothly to the shaft. The fossa m. brachialis is shallow, proximally elongate, and has its narrow proximal end close to the ventral margin of the fossil, making the fossa remarkably broad. The non-elevated tuberculum supracondylare ventrale is elongated and narrow, and has a narrow proximal margin close to the fossa m. brachialis. The facet where the ligamentum collaterale ventrale attaches to the tuberculum supracondylare ventrale is poorly defined, and slight ventral expansion distally. The proximal end of the facet lies only slightly proximal to the level of the proximal apex of the condylus dorsalis. The condylus ventralis is rounded with a proximodistal length about half of the condylus dorsalis. The two condyli are separated by a narrow incisura intercondylaris. On the caudal side, the sulcus scapulotricipitalis is well-defined and deep, with the two margins slightly converging proximally, and the processus flexorius is missing (Figs. 2, 4A3).
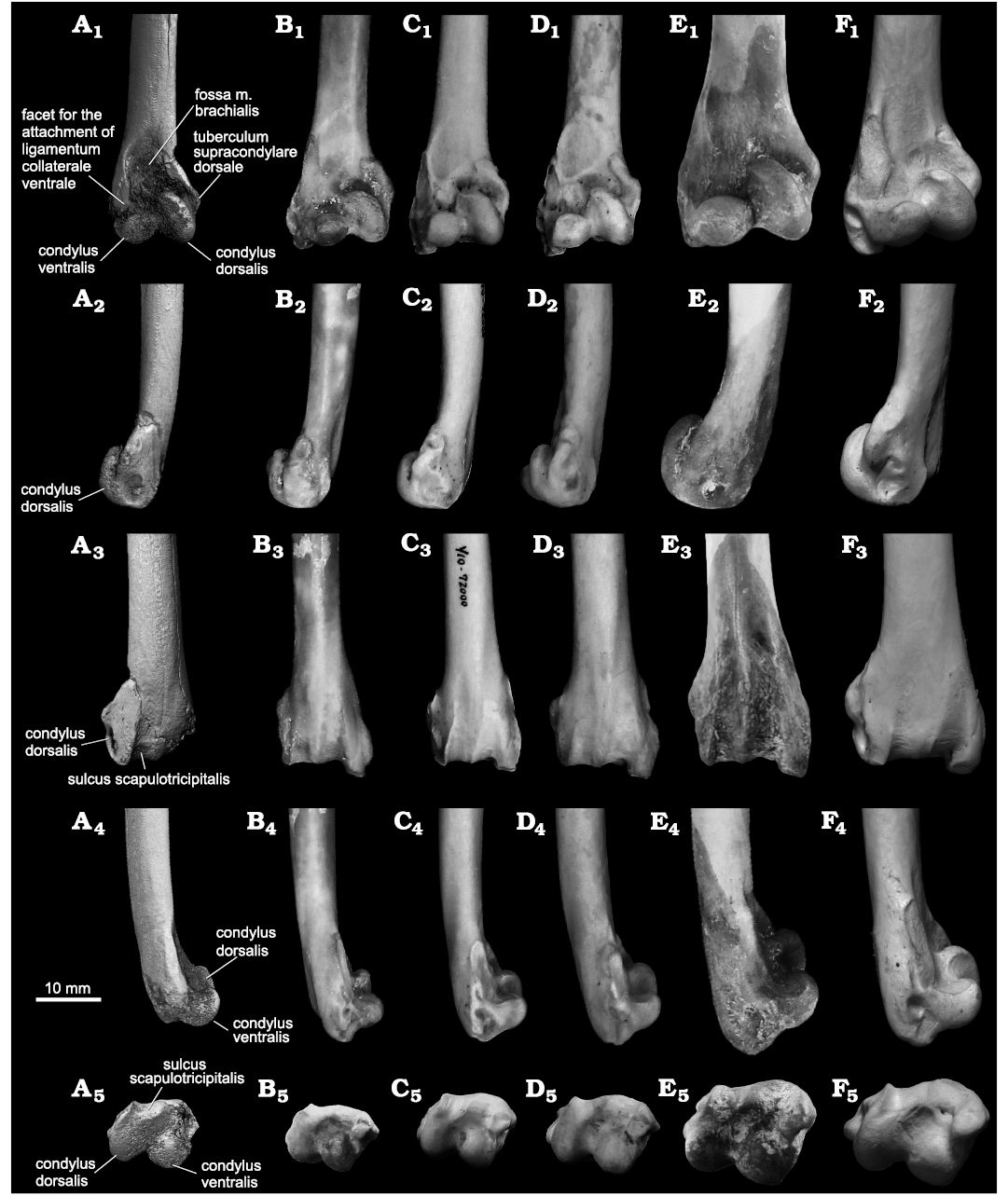
Fig. 4. The humeri of the fossil gaviid seabird Gavia sp. (A) and modern Gaviidae (B–F) in cranial (A1–F1), dorsal (A2–F2), caudal (A3–F3), ventral (A4–F4), and distal (A5–F5) views. A. CMM 2123 from Niubu, Chiayi, Taiwan, Liuchungchi Formation, Lower Pleistocene. B. Gavia stellata (Pontoppidan, 1763) (YIO 79598). C. Gavia arctica (Linnaeus, 1758) (YIO 72000). D. Gavia pacifica (Lawrence, 1858) (YIO 64763). E. Gavia adamsii (G.R. Gray, 1859) (YIO 63205). F. Gavia immer (Brünnich, 1764) (NHMUK S/1987.13.1). The images of CMM 2123 were obtained from CT. The image order of modern Gaviidae follows the taxonomy established by Sprengelmeyer (2014) and the photos of G. immer are courtesy of Junya Watanabe.
Remarks.—CMM 2123 is referred to Gavia based on (i) broad fossa m. brachialis that comes close to the medial margin proximally, (ii) the elongate, narrow, and non-elevated tuberculum supracondylare ventrale, (iii) pronounced dorsally projected tuberculum supracondylare dorsale, and (iv) shaft notched craniodorsally proximally-adjacent to the tuberculum supracondylare dorsale. CMM 2123 represents an adult as displayed by the smooth surface, and the two condyli and processes with clear shapes as displayed in the modern adult specimens (Fig. 4). In addition, CMM 2123 shares the following qualitative features with humeri of modern Gavia adamsii (G.R. Gray, 1859): the relatively sharp (in cranial aspect) apex of tuberculum supracondylare dorsale, the smooth dorsal surface on the proximal side of tuberculum supracondylare dorsale, narrow (less than half width of the tuberculum) and non-elevated proximal end of the facet for the attachment of ligamentum collaterale ventrale on the tuberculum supracondylare ventrale, and the sulcus scapulotricipitalis with deep distal end and slightly proximally converged margins (Figs. 2, 4). However, the fossil humerus is rather smaller than those of the examined G. adamsii specimens, and is also relative larger than the examined G. stellata (Pontoppidan, 1763) specimens (Fig. 4B, E; Table 1). This excludes the possibility of identifying the humerus as either Gavia adamsii or G. stellata. Gavia artica (Linnaeus, 1758) (Fig. 4C) and Gavia pacifica (Lawrence, 1858) (Fig. 4D), while of similar size to the fossil (Table 1), differ by having larger facets for the attachment of ligamentum collaterale ventrale on the tuberculum supracondylare ventrale, having prominent scars for the palmar branch of m. extensor carpi radialis that interrupt the dorsal profile proximal to the apex of the tuberculum supracondylare dorsale, and a much shallower sulcus scapulotricipitalis; G. immer (Fig. 4F) is larger, and has a much more massive facet for the attachment of ligamentum collaterale ventrale on the tuberculum supracondylare ventrale that more greatly separates the fossa brachialis from the ventral shaft facies, a prominent scar for the palmar branch of m. extensor carpi radialis that interrupts the dorsal profile proximal to the apex of the tuberculum supracondylare dorsale, a less rounded condylus ventralis, and a much broader sulcus scapulotricipitalis that is bound ventrally by a lower ridge.
Gavia stellata (Pontoppidan, 1763)
Figs. 2B, 5A.
Material.—CMM 2124 tibiotarsus, Bazhang River, Niubu area, Taiwan; Liuchungchi Formation (Fig. 3); 1.95–1.35 Ma.
Measurements.—See Table 1.
Description.—The shaft of the tibiotarsus CMM 2124 is slightly compressed craniocaudally with an oval cross section, and the distal end expands medially, with and the condylus medialis extending slightly further distally than the condylus lateralis. The articular scar for the distal fibula is, most proximally, on the lateral side of the shaft. It distally locates increasingly cranially and terminates near the proximal end of the medial tuberculum retinaculum m. fibularis. The medial tuberculum retinaculum m. fibularis is more elevated cranially and extends farther proximally than its lateral counterpart, which is prominent laterally, projecting slightly further laterally than the condylus lateralis. Proximally, both tubercula converge cranially, and distally, they converge laterally, which makes the sulcus m. fibularis shallow. The sulcus m. fibularis sits between the tubercula and faces slightly laterally. Distally, the sulcus m. fibularis passes caudally to a small lateral prominence at the distal end of the lateral tuberculum retinaculum m. fibularis. The medial tuberculum extends distally to join a small narrow laterally-facing crest and hence to the condylus lateralis at its proximolateral point. The effect of this is that proximally the sulcus m. fibularis is enclosed by medial and lateral tubercula, but at the level of the proximal margin of the pons supratendineus, the sulcus passes onto the lateral shaft facies. A distinct sulcus sits adjacent to the proximal end of condylus lateralis and is bounded laterally by the distal end of the medial tuberculum retinaculum m. fibularis. This sulcus is interpreted as the distolateral insertion scar of the transverse ligament. The pons supratendineus is wide proximodistally, and its proximomedial side has a rounded and elevated insertion scar, for the proximomedial insertion of the transverse ligament. The canalis extensorius is centered on the shaft and opens widely proximodistally between the condyli, and is close to the condylus medialis.
Remarks.—The fossil tibiotarsus displays several characteristics that are typical of Gavia: (i) parallel condyli that are proximodistally shorter than the distal width of the bone and are medially inclined distally, (ii) proximally-elevated medial and laterally-projected lateral tuberculi retinaculum m. fibularis, with a shallow sulcus m. fibularis in-between, (iii) a proximodistally wide pons supratendineus, (iv) a rounded scar abutting the proximomedial margin of the pons supratendineus for the proximomedial insertion of the transverse ligament, (v) a pit sits beside the end of the thin ridge immediately proximal to the condylus lateralis for the distolateral insertion of the transverse ligament, (vi) a wide opening of the canalis extensorius which is aligned proximodistally between the condyli (not proximal to the condyli) and is medially close to the condylus medialis, and (vii) a proximodistally short incisura intercondylaris with respect to the adjacent condyli. The fossil is similar to G. stellata, especially regarding the fact that the caudomedial margin of the condylus medialis lacks a caudomedially prominent rounded projection as seen in all the other modern gaviids compared (Fig. 5). This characteristic clearly separates the fossil and G. stellata from the other extant species of Gaviidae. While the mediolateral width of the shaft with respect to the distal end of the fossil in the cranial view seems to be smaller than G. stellata (see Table 1), we think this disparity falls within intraspecific scope and, therefore, we prefer to identify the fossil as G. stellata (Table 1).
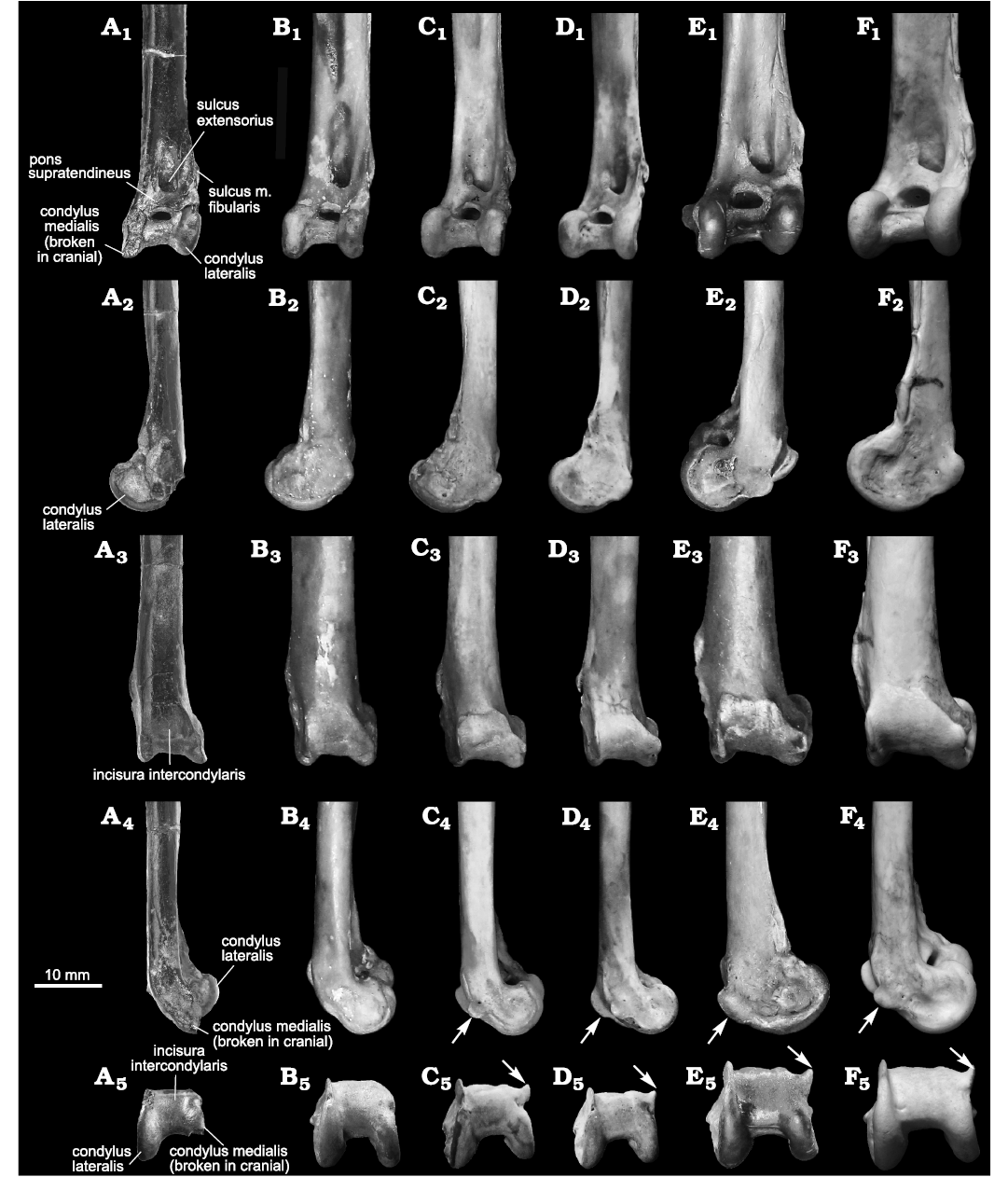
Fig. 5. Tibiotarsi of the fossil gaviid seabird Gavia stellata (Pontoppidan, 1763) (A) and modern Gaviidae (B–F), in cranial (A1–F1), lateral (A2–F2), caudal (A3–F3), medial (A4–F4), and distal (A5–F5) views. A. CMM 2124 from Niubu, Chiayi, Taiwan, Liuchungchi Formation, Lower Pleistocene. B. Gavia stellata (Pontoppidan, 1763) (YIO 79598). C. Gavia arctica (Linnaeus, 1758) (YIO 79597). D. Gavia pacifica (Lawrence, 1858) (YIO 60621). E. Gavia adamsii (G.R. Gray, 1859) (YIO 74354); F. Gavia immer (Brünnich, 1764) (NHMUK S/1987.13.1). Arrows on C4–F4 and C5–F5 indicate the caudomedial process present in these taxa that is lacking in G. stellata. The image order of modern Gaviidae follows the taxonomy established by Sprengelmeyer (2014) and the photos of G. immer are courtesy of Junya Watanabe.
Discussion
Ecology of gaviids and paleoenvironmental interpretation.—The Gaviidae (loons), the modern representatives of Gaviiformes, include five species: Gavia stellata, G. artica, G. pacifica, G. adamsii, and G. immer (HBW and BirdLife International 2022), all of which are migratory seabirds that are entirely limited to the Northern Hemisphere. They mate and breed near mid- and high-latitude lakes during the summer, and migrate south in the winter, when they predominantly stay at sea or near sea coasts. Fishes are their main diet, but crustaceans, mollusks, worms, and plants are also common (BirdLife International 2023). In the Northwest Pacific, all species of Gavia except G. immer are distributed around most islands and along the coastlines of the Asian continent but not in Taiwan, where the subtropical-tropical climate at 20° N is not suitable for them. During the past 160 years, only two Gavia species, G. arctica and G. stellata, have been rare occasional visitors to mainly the northeastern parts of Taiwan (Fig. 6; SOM 1: table S2, Supplementary Online Material available at http://app.pan.pl/SOM/app68-Wu_etal_SOM.pdf) (Swinhoe 1863; Yen 1984; Chu 1996; Chao 2006; National Taiwan Museum 2023; Taiwan Biodiversity Network 2023). Only two seabird fossils have been found in the Liuchungchi Formation and it seems unlikely that a species as rare as modern gaviids are in the region today would be represented in such a meagre fossil record. By this reasoning, the fossils likely represent a greater abundance of gaviids in the region in the Early Pleistocene. This in turn suggests that the two fossil Gavia species were deposited during a cooler climate interval during the oscillation of glacial-interglacial periods (Lisiecki and Raymo 2005; Huybers 2006; Liautaud et al. 2020; Vaucher et al. 2021). If so, this suggests that the Early Pleistocene paleo-avifauna in southwestern Taiwan included regular visitors of species of Gavia reflecting a period when the Gaviidae expanded their distribution south to the subtropical Northwest Pacific during the relatively cold time of the Early Pleistocene. The change in oceanic currents associated with cooler glacial period climate, e.g., the Kuroshio Current in the Northwest Pacific, may have brought upwellings that supported both marine and coastal ecosystems in the ancient Taiwan Strait. This hypothesis is supported by a highly diverse fossil record of rays, sharks and bony fishes during the Early Pleistocene in the Liuchungchi Formation (Lin et al. 2018, 2021, 2022).
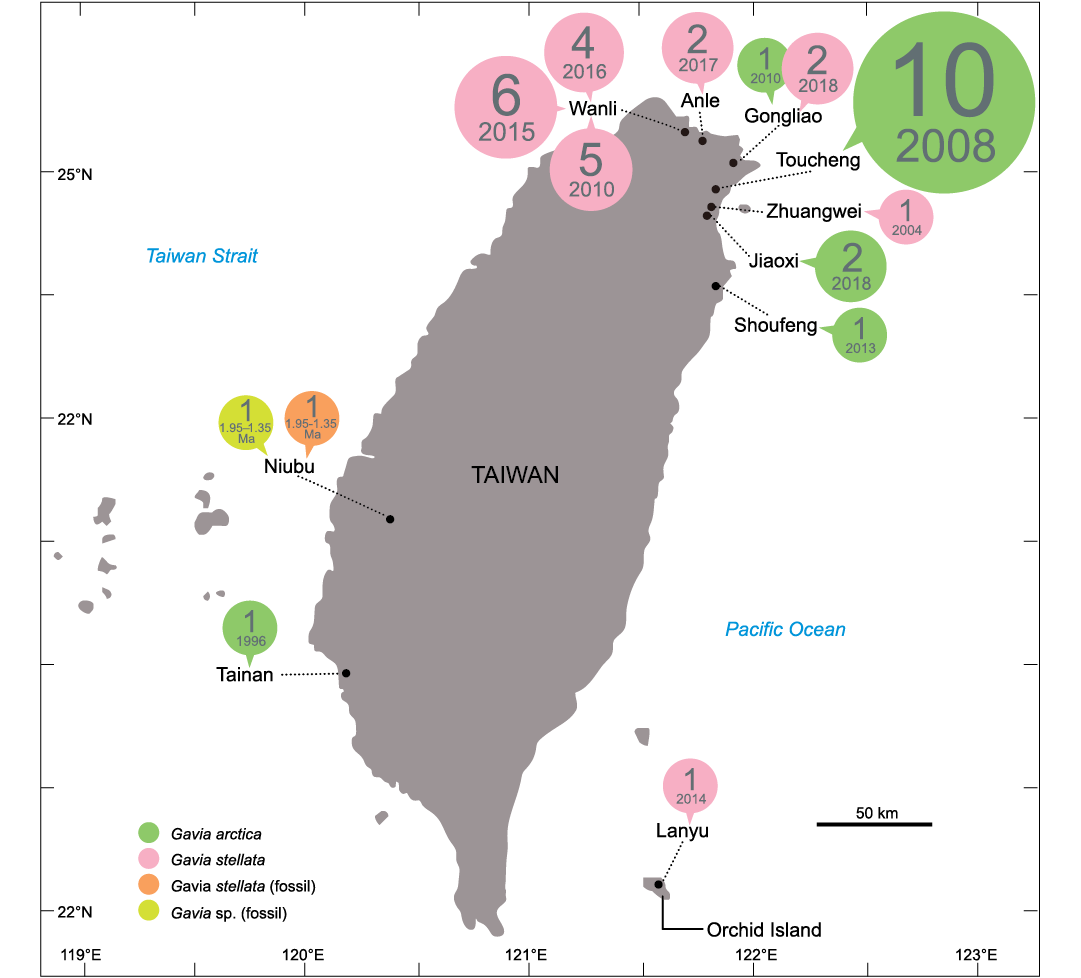
Fig. 6. Records of Gaviidae in Taiwan. The records are displayed as the number of occurrences (1–10) and dates (year or Ma). Records between 1861–1862 and between 1970–1971 are not included because the accurate localities are not accessible. See SOM 1: table S2 for details.
Fossil gaviids in Taiwan and their significance to the gaviid records in the Pleistocene.—The humerus CMM 2123 we identify as an indeterminate species of Gavia is smaller than the few modern specimens of G. adamsii we compared it to, but is more similar in qualitative features to this modern species than it is to any other modern Gavia species. These observations may be explained by three possible reasons. First, the specimen represent the small end of a larger size range than our samples of modern G. adamsii represents; however modern measurements of birds do not support this (Uher-Koch et al. 2020; US Fish & Wildlife Service 2023). Second, the fossil derives from birds in the Early Pleistocene population that consisted of smaller individuals than their modern counterparts. Third, this fossil represents an extinct sister-species to G. adamsii, characterized by the same qualitative features and smaller size. Resolution of this issue will require further fossils. The fossil records of G. adamsii are quite scarce, restricted to the North Pacific, and all come from the Holocene (Friedmann 1934a, b, 1935, 1941). In contrast, G. stellata has numerous Pleistocene fossil records from Europe, Asia and the Northeast Pacific (Fig. 7, SOM 1: table S3). The relative abundance of fossil records for G. stellata and G. adamsii is consistent with their modern distribution, as the former is widely distributed in mid- and high-latitude areas, whereas the latter is mainly found in a narrower zone in coastal regions of high-latitude areas (BirdLife International 2023). Most Pleistocene gaviid records are restricted to mid- and high-latitude regions of the Northern Hemisphere, and a few records from low-latitudes are from Mexico (Corona Martínez 2009) and Florida (Brodkorb 1953, 1963; Woolfenden 1959; Emslie 1995, 1998) (Fig. 7). The gaviid fossils from Taiwan provide another low-latitude occurrence, the first for the Pacific.
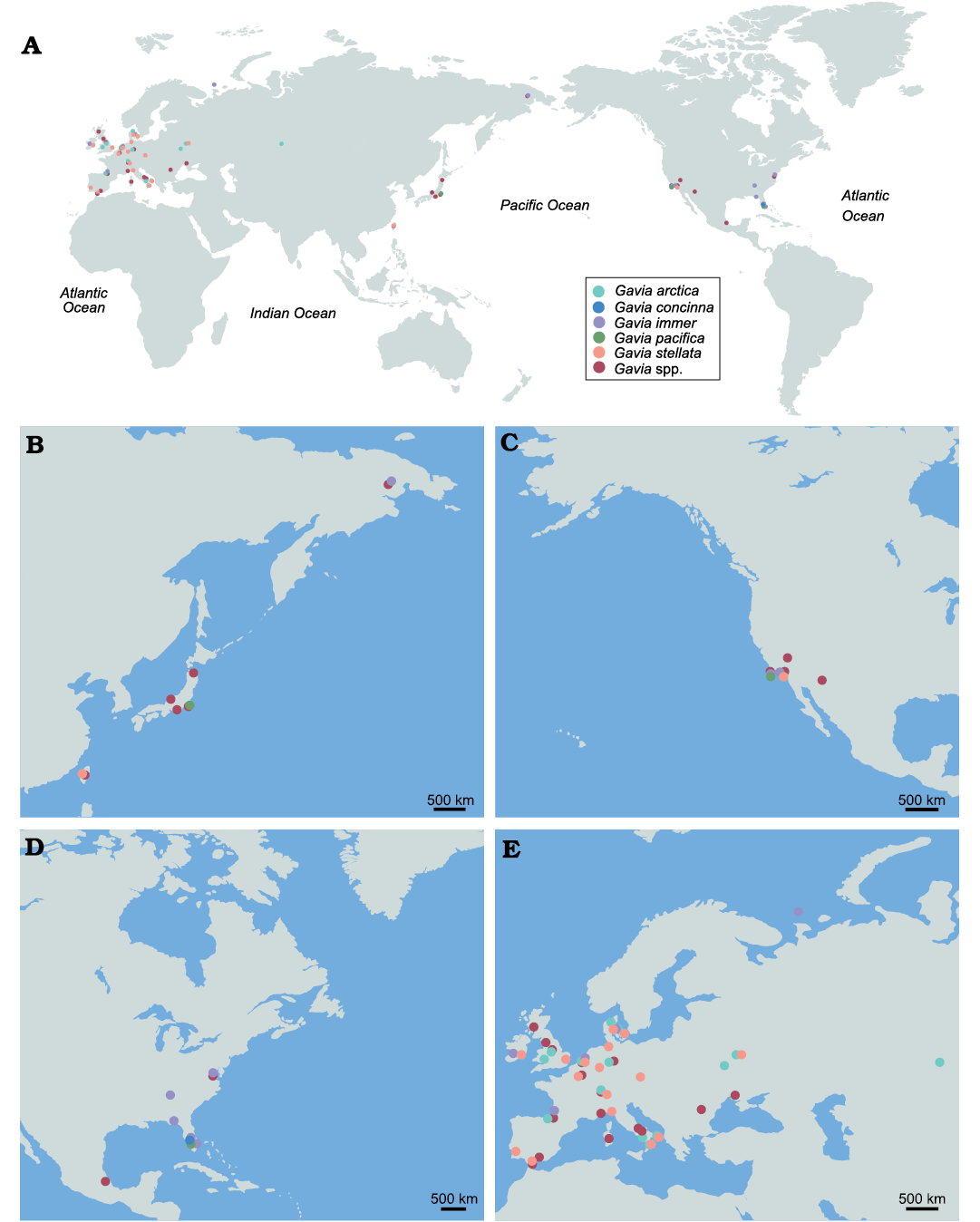
Fig. 7. Distribution of Pleistocene records of Gaviidae. A. The distribution around the world. Regional distribution in the Northwest Pacific (B), the Northeast Pacific (C), the Northwest Atlantic (D), and the Northeast Atlantic (E). Taxa with uncertainty are grouped into Gavia spp. See SOM 1: table S3 for details.
Conclusions
The two fossil species of Gavia from Taiwan, G. stellata, and an indeterminate species, are the first Pleistocene records of Gaviidae in the subtropical Northwest Pacific. They not only suggest a southern expansion of Pleistocene Gaviidae populations in the Northwest Pacific, but also hint at a distinct Pleistocene paleoavifauna in southern Taiwan, where the current subtropical climate is not suitable for Gaviidae. Therefore, the climate may have been colder when the two birds were alive in the Early Pleistocene, and the ancient Taiwan Strait may have been more productive and supported seabirds like Gaviidae and the greater diversity of fish evidenced by fossils. One of the two fossils is a species of Gavia which is much smaller than G. adamsii, yet similar in morphological features. To solve the taxonomic implications of the size disparity between it and G. adamsii requires more fossil remains.
Authors’ contributions
SMW and CHL conceived and designed the project and wrote the paper. SMW collected the data. THW and SMW worked on the taxonomic identification. CKC conducted the biostratigraphy study. All authors edited the manuscript.
Acknowledgements
We would like to express our sincere gratitude to Junya Watanabe (University of Cambridge, UK) who helped us at the initial stage of the study and generously provided us with a wealth of relevant data, as well as supplying photographs of Gavia immer. We also thank Taku Mizuta, Takeshi Yamasaki, Yasuko Iwami, Sayaka Kobayashi, and Tomoko Imamura (YIO) for access to the collections in Japan, and Yen-Jean Chen (National Museum of Natural Science, Taichung, Taiwan; NMNS), Meng-Min Hsueh (Biodiversity Research Museum, Academia Sinica, Taipei, Taiwan) and Jun-Tsong Lin and Fu-Chun Yang (National Taiwan Museum, Taipei, Taiwan) for access to the bird collections in Taiwan. We thank Guan-Shuo Mai (Biodiversity Research Center, Academia Sinica, Taipei, Taiwan; BRCAS), Chie-Jen Ko and Da-Li Lin (Taiwan Biodiversity Research Institute, Nantou, Taiwan) for general data of migratory birds in Taiwan. We thank the Taiwan Mouse Clinic, Academia Sinica and Taiwan Animal Consortium for the technical support in computed tomography (CT), and Hao-Chun Hsu (BRCAS), for the postprocess of CT slices. We thank Sheng-Tzung Wang (Central Geological Survey, Ministry of Economic Affairs, New Taipei City, Taiwan) for nannofossil analysis, and Chih-Wei Chien (CPC Corporation Taiwan, Mioli, Taiwan) and Tzu-Ruei Yang (NMNS) for geological background. This study was also helped by initial discussion with Ursula Göhlich (Natural History Museum, Vienna, Austria) and Genway Gao (California, USA). We thank the assistance of I-Ju Hsiao (CMM) and Hsin-Wei Liu (BRCAS) who helped in dealing with the loan process of the bird fossils. Finally, we thank the editor Daniel E. Barta (Oklahoma State University, Oklahoma, USA) and reviewers Gerald Mayr (Senckenberg Research Institute and Natural History Museum Frankfurt, Frankfurt, Germany) and Hanneke Meijer (University of Bergen, Bergen, Norway) for their comments on the manuscript. This study was supported by the Intramural funding of Academia Sinica, Taipei, Taiwan and Ministry of Science and Technology, Taiwan (Grant No. 110-2116-M-001-009, 111-2116-M-001-033, and 112-2116-M-001-017-MY3) to CHL.
References
Aotsuka, K., Isaji, S., and Endo, H. 2022. An avian sternum (Aves: Procellariidae) from the Pleistocene Ichijiku Formation in Chiba, Japan. Paleontological Research 26: 74–86. Crossref
Baumel, J.J. and Witmer, L.M. 1993. Osteologia. In: Baumel, J.J., King, A.S., Breazile, J.E., Evans, H.E. and Vanden Berge, J.C. (eds.), Handbook of Avian Anatomy: Nomina Anatomica Avium (2nd Ed.). Publications of the Nuttall Ornithological Club 23, 45–132. Nuttall Ornithological Club, Cambridge, Massachusetts.
BirdLife International 2023. IUCN Red List for Birds [downloaded from http://datazone.birdlife.org on 10 March 2023].
Block, B.A., Jonsen, I.D., Jorgensen, S.J., Winship, A.J., Shaffer, S.A., Bograd, S.J., Hazen, E.L., Foley, D.G., Breed, G.A., Harrison, A.L., Ganong, J.E., Swithenbank, A., Castleton, M., Dewar, H., Mate, B.R., Shillinger, G.L., Schaefer, K.M., Benson, S.R., Weise, M.J., Henry, R.W., and Costa, D.P. 2011. Tracking apex marine predator movements in a dynamic ocean. Nature 475: 86–90. Crossref
Brodkorb, P. 1953. A review of the Pliocene loons. The Condor 55 (4): 211–214. Crossref
Brodkorb, P. 1963. Catalogue of fossil birds. Part 1 (Archaeopterygiformes through Ardeiformes). Bulletin of the Florida State Museum. Biological sciences 7 (4): 179–293.
Brünnich, M.T. 1764. Ornithologia borealis, sistens collectionem avium ex omnibus, Imperio Danico subjectis, provinciis insulisqve borealibus Hafniae factam, cum descriptionibus novarum, nominibus incolarum, locis natalium et icone. 80 pp. J.C. Kall, Copenhagen. Crossref
Buckeridge, J.S., Chan, B.K.K., and Lee, S.-W. 2018. Accumulations of fossils of the whale barnacle Coronula bifida Bronn, 1831 (Thoracica: Coronulidae) provides evidence of a Late Pliocene cetacean migration route through the Straits of Taiwan. Zoological Studies 57: (54): 1–12.
Chang, H.-C. 2008. Geological Map of Taiwan. Scale 1:50 000. Chiayi. Vol. 44 [in Chinese]. Central Geological Survey, Ministry of Economic Affairs, New Taipei City.
Chao, W.-K. 2006. Rare bird: red-throated loon [in Chinese]. Feather 215: 4.
Chen, W.-S. 2016. An Introduction to the Geology of Taiwan [in Chinese]. 204 pp. Geological Society of Taiwan, Taipei.
Chen, W.-S., Huang, N.-W., and Yang, C.-C. 2011. Pleistocene sequence stratigraphic characteristics and foreland basin evolution, southwestern Taiwan [in Chinese, with English abstract]. Special Publication of the Central Geological Survey (MOEA) 25: 1–38.
Chi, W.-R. 1980. Calcareous nannoplankton biostratigraphic study and correlation of the Late Neogene sequence in the Chiayi and Hsinying foothills, southern Taiwan [in Chinese, with English abstract]. Proceedings of the Geological Society of China 23: 16–28.
Chu, T.-T. 1996. Injured birds frequently found on the coast of Tainan: another Arctica loon rescued [in Chinese]. Min Sheng Pao [downloaded from udndata.com (https://udndata.com/ndapp/Index?cp=udn), Dec. 15].
Clay, T.A., Phillips, R.A., Manica, A., Jackson, H.A., and Brooke, M.D.L. 2017. Escaping the oligotrophic gyre? The year-round movements, foraging behaviour and habitat preferences of Murphy’s petrels. Marine Ecology Progress Series 579: 139–155. Crossref
Collins, P.W., Guthrie, D.A., Whistler, E.L., Vellanoweth, R.L. and Erlandson, J.M. 2018. Terminal Pleistocene–Holocene avifauna of San Miguel and Santa Rosa islands: identifications of previously unidentified avian remains recovered from fossil sites and prehistoric cave deposits. Western North American Naturalist 78: 370–403. Crossref
Corona Martínez, E. 2009. Las aves en el Cenozoico tardío de México. 300 pp. Universidad Autónoma de Madrid, Madrid.
Croxall, J.P., Butchart, S.H.M., Lascelles, B., Stattersfield, A.J., Sullivan, B., Symes, A. and Taylor, P. 2012. Seabird conservation status, threats and priority actions: a global assessment. Bird Conservation International 22: 1–34. Crossref
Croxall, J.P., Reid, K., and Prince, P.A. 1999. Diet, provisioning and productivity responses of marine predators to differences in availability of Antarctic krill. Marine Ecology Progress Series 177: 115–131. Crossref
Drummond, B.A., Orben, R.A., Christ, A.M., Fleishman, A.B., Renner, H.M., Rojek, N.A., and Romano, M.D. 2021. Comparing non-breeding distribution and behavior of red-legged kittiwakes from two geographically distant colonies. PLOS ONE 16 (7): e0254686. Crossref
Dyke, G.J., Wang, X., and
Habib, M.B. 2011. Fossil plotopterid seabirds from the Eo-Oligocene of
the Olympic Peninsula (Washington State, USA): descriptions and
functional morphology. PLOS ONE
6 (10): e25672. Crossref
Emslie, S.D. 1995. An early Irvingtonian avifauna from Leisey Shell Pit, Florida. Bulletin of the Florida Museum of Natural History 37: 299–344.
Emslie, S.D. 1998. Avian community, climate, and sea-level changes in the Plio-Pleistocene of the Florida Peninsula. Ornithological Monographs 50: 1–113. Crossref
Friedmann, H. 1934a. Bird bones from Eskimo ruins on St. Lawrence Island, Bering Sea. Journal of the Washington Academy of Sciences 24 (2): 83–96.
Friedmann, H. 1934b. Bird bones from old Eskimo ruins in Alaska. Journal of the Washington Academy of Sciences 24 (5): 230–237.
Friedmann, H. 1935. Avian bones from prehistoric ruins on Kodiak Island, Alaska. Journal of the Washington Academy of Sciences 25 (1): 44–51.
Friedmann, H. 1941. Bird bones from Eskimo ruins at Cape Prince of Wales, Alaska. Journal of the Washington Academy of Sciences 31 (9): 404–409.
Gartner, S. Jr. 1969. Correlation of Neogene planktonic foraminifer and calcareous nannofossil zones. AAPG Bulletin 53 (9): 2036. Crossref
Goedert, J.L. 1988. A new late Eocene species of Plotopteridae (Aves: Pelecaniformes) from northwestern Oregon. Proceedings of the California Academy of Sciences 45 (4): 97–102.
Goedert, J.L. and Cornish, J. 2002. A preliminary report on the diversity and stratigraphic distribution of Plotopteridae (Pelecaniformes) in Paleogene rocks of Washington State, USA. In: Z. Zhou and F. Zhang (eds.), Proceedings of the 5th Symposium of the Society of Avian Paleontology and Evolution, 63–76. Science Press, Beijing.
Gray, G.R. 1859. Description of a new species of diver (Colymbus). Proceedings of the Zoological Society of London 27 (2): 167.
Guthrie, D.A. 1992. A late Pleistocene avifauna from San Miguel Island, California. In: K.E. Campbell Jr. (ed.), Papers in Avian Paleontology Honoring Pierce Brodkorb. Los Angeles County Natural History Museum Science Series 36: 319–327.
Guthrie, D.A. 2005. Distribution and provenance of fossil avifauna on San Miguel Island. In: D.K. Garcelon and C.D. Schwemm (eds.), Proceedings of the 6th California Islands Symposium. NPS Technical Publications CHIS-05-01, 35–42. Institute for Wildlife Studies, Arcata.
Harington, C.R. 1978. Quaternary vertebrate faunas of Canada and Alaska and their suggested chronological sequence. Syllogeus 15: 105.
Hasegawa, Y., Tomida, Y., Kohno, N., Ono, K., Nokariya, H., and Uyeno, T. 1988. Quaternary vertebrates from Shiriya area, Shimokita Peninsula, northeastern Japan [in Japanese, with English summary]. Memoirs of the National Science Museum (Tokyo) 21: 17–33.
HBW and BirdLife International 2022. Handbook of the Birds of the World and BirdLife International digital checklist of the birds of the world. Version 6b [available online, http://datazone.birdlife.org/userfiles/file/Species/Taxonomy/HBW-BirdLife_Checklist_v6b_Jul22.zip].
Howard, H. 1936. A new fossil bird locality near Playa del Rey, California, with description of a new species of sulid. The Condor 38 (5): 211–214. Crossref
Howard, H. 1949. Avian fossils from the marine Pleistocene of Southern California. The Condor 51 (1): 20–28. Crossref
Howard, H. 1958. Further records from the Pleistocene of Newport Bay Mesa, California. The Condor 60 (2): 136. Crossref
Hu, C.-H. 1989. Manual for ten geological routes in central Taiwan, route 7: geology along the Nanheng Highway, Tsailiao, Peiliao, Chiahsien [in Chinese]. In: Faculty Members of the Department of Earth Sciences (ed.), Field Manual of the Geology of Taiwan (II), 105–163. Department of Earth Sciences, National Taiwan Normal University, Taipei.
Hu, C.-H. and Tao, H.-J. 1996. Crustacean Fossils of Taiwan [in Chinese]. 228 pp. Ta-Jen Printers, Taipei.
Hu, C.-H. and Tao, H.-J. 2004. Studies on the Neogene crabs from south-western foothills of Taiwan. Acta Palaeontologica Sinica 43: 537–555.
Huang, T.-C. and Ting, J.-S. 1981. Calcareous nannofossil biostratigraphy of the late Neogene sallow marine deposits in Taiwan [in Chinese, with English abstract]. Ti-Chin 3: 105–109.
Huybers, P. 2006. Early
Pleistocene glacial cycles and the integrated summer insolation
forcing. Science 313:
508–511. Crossref
Kimura, M., Sakurai, K., and Katoh, T. 1998. An extinct fossil bird (Plotopteridae) from the Tokoro Formation (late Oligocene) in Abashiri City, northeastern Hokkaido, Japan. Journal of Hokkaido University of Education (Section IIB) 48: 29–34.
Lawrence, G.N. 1858. Report of Explorations and Surveys to Ascertain the Most Practical and Economical Route For a Railroad from the Mississiippi River to the Pacific Ocean. Birds. Vol. 9. lvi + 1055 pp. A.O.P. Nicholson, Washington.
Liautaud, P.R., Hodell, D.A.
and Huybers, P.J. 2020. Detection of significant climatic precession
variability in early Pleistocene glacial cycles. Earth
and Planetary Science Letters 536: 116137. Crossref
Lin, C.-H., Chien, C.-W., Lee, S.-W., and Chang, C.-W. 2021. Fish fossils of Taiwan: a review and prospection. Historical Biology 33: 1362–1372. Crossref
Lin, C.-Y., Lin, C.-H., and Shimada, K. 2022. A previously overlooked, highly diverse early Pleistocene elasmobranch assemblage from southern Taiwan. PeerJ 10: e14190. Crossref
Lin, C.-H., Wang, L.-C., Wang, C.-H., and Chang, C.-W. 2018. Common early Pleistocene fish otoliths from Niubu in Chia-Yi County, southwestern Taiwan. Journal of the National Taiwan Museum 71 (3): 47–68.
Linnaeus, C. 1758. Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. 824 pp. Laurentius Salvius, Stockholm. Crossref
Lisiecki, L.E. and Raymo, M.E. 2005. A Pliocene–Pleistocene stack of 57 globally distributed benthic δ18O records. Paleoceanography 20: 1–17. Crossref
Matsuoka, H. and Hasegawa, Y. 2018. Birds around the Minatogawa man: the late Pleistocene avian fossil assemblage of the Minatogawa Fissure, southern part of Okinawa Island, central Ryukyu Islands, Japan. Bulletin of Gunma Museum of Natural History 22: 1–21.
Mayr, G. 2015. A new Paleogene procellariiform bird from western North America. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 275: 11–17. Crossref
Mayr, G. 2022. Paleogene Fossil Birds. 251 pp. Springer, Cham. Crossref
Mayr, G. and Goedert, J.L. 2017. Oligocene and Miocene albatross fossils from Washington State (USA) and the evolutionary history of North Pacific Diomedeidae. The Auk 134: 659–671. Crossref
Mayr, G. and Goedert, J.L. 2018. First record of a tarsometatarsus of Tonsala hildegardae (Plotopteridae) and other avian remains from the late Eocene/early Oligocene of Washington State (USA). Geobios 51: 51–59. Crossref
Mayr, G. and Goedert, J.L. 2022. New late Eocene and Oligocene plotopterid fossils from Washington State (USA), with a revision of “Tonsala” buchanani (Aves, Plotopteridae). Journal of Paleontology 96: 224–236. Crossref
Mayr, G., Goedert, J.L., and Vogel, O. 2015. Oligocene plotopterid skulls from western North America and their bearing on the phylogenetic affinities of these penguin-like seabirds. Journal of Vertebrate Paleontology 35 (4): 1–14. Crossref
Miller, A.H. 1931. An auklet from the Eocene of Oregon. University of California Publications, Bulletin of the Department of Geological Sciences 20: 23–26.
Miller, L. 1914. Bird remains from the Pleistocene of San Pedro, California. University of California Publications, Bulletin of the Department of Geological Sciences 8 (4): 31–38.
Miller, L. 1925. Chendytes, a diving goose from the California Pleistocene. The Condor 27 (4): 145–147. Crossref
Miller, L. 1930. Further bird remains from the upper San Pedro Pleistocene. The Condor 32 (2): 116–118. Crossref
National Taiwan Museum 2023. National Taiwan Museum Digital Archive Information System. Downloaded from https://collections.culture.tw/ntm_cms/ on 12 July 2023.
Ohashi, T. and Hasegawa, Y. 2020. New species of Plotopteridae (Aves) from the Oligocene Ashiya Group of Northern Kyushu, Japan. Paleontological Research 24: 285–297. Crossref
Olson, S.L. and Hasegawa, Y. 1996. A new genus and two new species of gigantic Plotopteridae from Japan (Aves: Pelecaniformes). Journal of Vertebrate Paleontology 16: 742–751. Crossref
Olson, S.L. and Rasmussen, P. C. 2001. Miocene and Pliocene birds from the Lee Creek Mine, North Carolina. Smithsonian Contributions to Paleobiology 90: 233–365. Crossref
Parsons, M., Mitchell, I., Butler, A., Ratcliffe, N., Frederiksen, M., Foster, S., and Reid, J.B. 2008. Seabirds as indicators of the marine environment. ICES Journal of Marine Science 65 (8): 1520–1526. Crossref
Piatt, J.F., Sydeman, W.J., and
Wiese, F. 2007. Introduction: a modern role for seabirds as
indicators. Marine Ecology Progress
Series 352: 199–204. Crossref
Pontoppidan, E. 1763. Den Danske Atlas eller Konge-Riget Dannemark. Vol. 1, 723 pp., Godiche, Copenhagen.
Sakurai, K., Kimura, M., and Katoh, T. 2008. A new penguin-like bird (Pelecaniformes: Plotopteridae) from the late Oligocene Tokoro Formation, northeastern Hokkaido, Japan. Oryctos 7: 83–94.
Sprengelmeyer, Q.D. 2014. A Phylogenetic Reevaluation of the Genus Gavia Using Next-Generation Sequencing. 43 pp. Master’s Thesis, Northern Michigan University, Marquette.
Swinhoe, R. 1863. The ornithology of Formosa or Taiwan. Ibis 5: 377–435. Crossref
Taiwan Biodiversity Network. 2023. TBN Home Page. Taiwan Endemic Species Research Institute. Downloaded from https://www.tbn.org.tw/ on 2 March 2023.
Tao, H.-J. 1993. A new Miocene fossil species Priacanthus liui (Pisces: Perciformes) from the Nanchung Formation in Chiayi Hsien, Taiwan. Bulletin of the National Museum of Nature and Science 4: 91–100.
Tsai, C.-H. and Mayr, G. 2021. A phasianid bird from the Pleistocene of Tainan: the very first avian fossil from Taiwan. Journal of Ornithology 162: 919–923. Crossref
Tyrberg, T. 1998. Pleistocene Birds of the Palearctic: A Catalogue. 720 pp. Nuttall Ornithological Club, Cambridge.
Uher-Koch, B.D., North, M.R., and Schmutz, J.A. 2020. Yellow-billed Loon. In: S.M. Billerman (ed.), Birds of the World. Vol. 1. Cornell Laboratory of Ornithology, Ithaca [downloaded from https://doi.org/10.2173/bow.yebloo.01 on 8 May 2023]. Crossref
US Fish & Wildlife Service. 2023. Yellow-billed Loon Gavia adamsii. Downloaded from https://www.fws.gov/species/yellow-billed-loon-gavia-adamsii on 12 April 2023.
Vaucher, R., Dashtgard, S.E., Horng, C.S., Zeeden, C., Dillinger, A., Pan, Y.Y., Setiaji, R.A., Chi, W.R., and Löwemark, L. 2021. Insolation-paced sea level and sediment flux during the early Pleistocene in Southeast Asia. Scientific Reports 11 (1): 16707. Crossref
Wang, C.-N., Hsu, H.-C., Wang, C.-C., Lee, T.-K., and Kuo, Y.-F. 2015. Quantifying floral shape variation in 3D using microcomputed tomography: a case study of a hybrid line between actinomorphic and zygomorphic flowers. Frontiers in Plant Science 6: 724. Crossref
Warheit, K.I. 1992. A review of the fossil seabirds from the Tertiary of the North Pacific: plate tectonics, paleoceanography, and faunal change. Paleobiology 18: 401–424. Crossref
Watanabe, J. and Matsuoka, H. 2015. Flightless diving duck (Aves, Anatidae) from the Pleistocene of Shiriya, northeast Japan. Journal of Vertebrate Paleontology 35 (6): e994745. Crossref
Watanabe, J., Koizumi, A., Nakagawa, R., Takahashi, K., Tanaka, T., and Matsuoka, H. 2020. Seabirds (Aves) from the Pleistocene Kazusa and Shimosa groups, central Japan. Journal of Vertebrate Paleontology 39 (5): 1–20. Crossref
Watanabe, J., Matsuoka, H., and Hasegawa, Y. 2016. Two species of Uria (Aves: Alcidae) from the Pleistocene of Shiriya, northeast Japan, with description and body mass estimation of a new species. Bulletin of the Gunma Museum of Natural History 20: 59–72.
Watanabe, J., Matsuoka, H., and Hasegawa, Y. 2018a. Pleistocene fossils from Japan show that the recently extinct Spectacled Cormorant (Phalacrocorax perspicillatus) was a relict. The Auk 135: 895–907. Crossref
Watanabe, J., Matsuoka, H., and Hasegawa, Y. 2018b. Pleistocene seabirds from Shiriya, northeast Japan: systematics and oceanographic context. Historical Biology 32: 671–729. Crossref
Woolfenden, G.E. 1959. A Pleistocene avifauna from Rock Spring, Florida. Wilson Bulletin 71: 183–187.
Xiao, M.-J. and Li, Z.-L. 2022. A Field Guide to the Birds of Taiwan [in Chinese]. 437 pp. Wild Bird Society of Taipei, Taipei.
Xue, W.-J. 2004. Chiayi Area Fossil Map [in Chinese]. 211 pp. Chiayi City Cultural Bureau, Chiayi.
Yen, C. 1984. Wild Birds in Taiwan: Migratory Birds. Vol. 2 [in Chinese]. 197 pp. Holiday, New Taipei City.
Acta Palaeontol. Pol. 68 (4): 613–624, 2023
https://doi.org/10.4202/app.01091.2023