
Floian, Early Ordovician, trilobites from the Olongbluk Terrane, northwest China
XIN WEI and ZHIQIANG ZHOU
Wei, X. and Zhou, Z.Q. 2023. Floian, Early Ordovician, trilobites from the Olongbluk Terrane, northwest China. Acta Palaeontologica Polonica 68 (4): 683–693.
Floian, Early Ordovician trilobites are systematically described and revised based on new material from the middle part of the Duoquanshan Formation of the Shihuigou area, northern Qinghai Province, northwest China. The fauna that lived on the shallow-water carbonate platform comprises three species belonging to two families, i.e., Tsaidamaspis diarmatus, Zhiyia tsinghaiensis, and Liexiaspis sp. indeterminate. It exhibits a strong endemicity to the Olongbluk terrane. The new isoteline genus Zhiyia is established on the basis of the material from the Olongbluk terrane and South China palaeoplate, and is characterized by its: (i) almost obsolete cephalic and pygidial axial furrows; (ii) flattened anterior border and narrow (sag., exsag.) occipital ring; (iii) bilobed hypostome with a shallow median notch and a small triangular median projection; (iv) subsemicircular pygidium with wide pygidial axis and border. Faunal evidence indicates that the palaeogeographic position of the Olongbluk terrane may have been situated closer to the South China palaeoplate rather than the North China palaeoplate during the Floian.
Key words: Trilobita, palaeogeography, Floian, Ordovician, Olongbluk terrane, northwest China.
Xin Wei [xwei@nigpas.ac.cn; ORCID: https://orcid.org/0000-0003-4095-280X ], State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Science, Nanjing 210008, China.
Zhiqiang Zhou [zsy1940@163.com; ORCID:https://orcid.org/0009-0003-5113-0913 ], Xi’an Institute of Geology and Mineral Resources, Chinese Academy of Geological Sciences, Xi’an 710054, China.
Received 21 August 2023, accepted 30 October 2023, available online 15 December 2023.
Copyright © 2023 X. Wei and Z. Zhou. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Qilian Orogenic Belt is located between the three major palaeoplates in China (i.e., North China, South China, and Tarim palaeoplates), and composed of seven nearly NW-SE trending subparallel stratigraphic subregions (Song et al. 2013; Fig. 1A). The Olongbluk area is a rather small and isolated terrane located between the Qaidam terrane and the South Qilian Mountains area (Fig. 1A), and bounded on the north and south by the Zongwulong Mountain and the Chaibei faults, respectively (Zhou and Lin 1995; Zhu et al. in press). The Ordovician succession crops out in the Dachaidan, Huaitoutala and Wulan areas, consisting of the Duoquanshan (Tremadocian–lower Darriwilian, Lower–Middle Ordovician), Shihuigou (lower–middle Darriwilian, Middle Ordovician), and Datouyanggou (Sandbian–Katian, Upper Ordovician) formations in ascending order (Mu et al. 1963). During the past few decades, the Ordovician fossils of this area have been reported and systematically studied, including trilobites (Chang and Fan 1960), nautiloids (Zhang 1965; Lai et al. 1984; Lai 1985), graptolites (Mu et al. 1962) and chitinozoans (Wang et al. 2018). However, the position of the Olongbluk terrane during the Ordovician is uncertain, with palaeogeographic reconstructions placing it either near South China or near North China palaeoplates (e.g., Cocks and Torsvik 2013; Wang et al. 2018). In this study, we provide a systematic description and revision of the trilobites from the middle part of the Floian Duoquanshan Formation in the Shihuigou section, and discuss the palaeogeographic relationships between the Olongbluk terrane and other palaeoplates during the Early Ordovician through analysis of their trilobite faunas.
Institutional abbreviation.—NIGPAS, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, China; NIGP, Nanjing Institute of Geology and Palaeontology, China.
Other abbreviations.—SHG, Shihuigou; exsag., exsagittal; sag., sagittal; tr., transverse; S1–5, glabellar furrows 1–5.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: lsid:zoobank.org:pub:AB6230A3-5AB9-466E-B272-6FD 74A89358F.
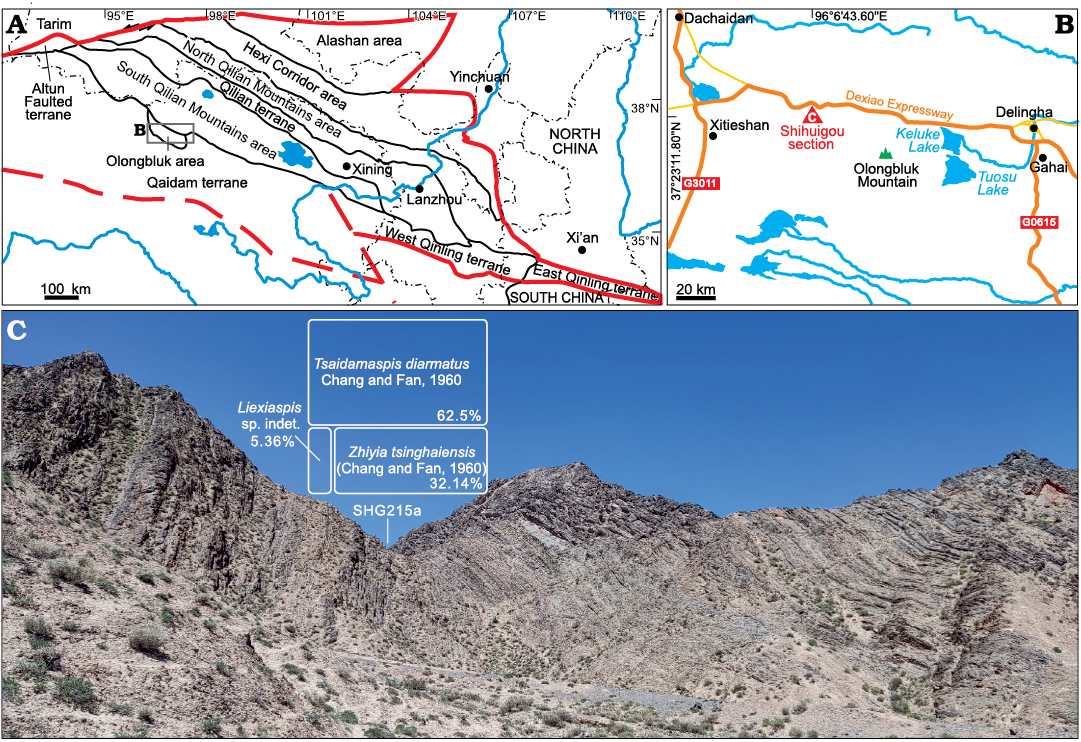
Fig. 1. The Ordovician palaeogeographic units of the Qilian and Qaidam regions (modified from Zhu et al. in press) (A), location of the Shihuigou section of Delingha, north Qinghai Province (B), and photograph of the middle part of the Duoquanshan Formation at the Shihuigou section and percentage data of trilobites from the sample SHG215a (C).
Geological setting, material and methods
The studied section is located at the Shihuigou (GPS: N 37°23′11.80′′, E 96°6′43.60′′), about 110 km west of Delingha, northern Qinghai Province (Fig. 1B). In this area, the Duoquanshan Formation is about 390 m thick and composed of bioclastic, argillaceous and nodular limestone beds. It is conformably overlain by the Shihuigou Formation (lower–middle Darriwilian) and underlain by the Olongbluk Group (Miaolingian–Furongian).
A total of 112 trilobite specimens were collected from the middle part of the Duoquanshan Formation (SHG215a, bioclastic limestone) at the Shihuigou section (Fig. 1C). Of these, three genera and three species of two families were identified, comprising Tsaidamaspis diarmatus Chang and Fan, 1960 (62.5%, N = 70), Zhiyia tsinghaiensis (Chang and Fan, 1960) (32.14%, N = 36) and Liexiaspis sp. indet. (5.36%, N = 6). The cranidia, librigenae, thoracic segments and pygidia of the three species are almost always disarticulated and buried separately due to high-energy depositional environment on a shallow-water carbonate platform. Figured specimens were lightly coated with magnesium oxide, and then photographed using a Sony a7RM4A digital camera. They were processed using Adobe Photoshop 2021 and CorelDRAW 2020. Specimens described in this paper are housed in the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (NIGP).
Sample SHG215a, from the middle part of the Duoquanshan Formation, yielded abundant trilobites and gastropods, together with a few conodonts and nautiloids. Some conodonts are identifiable in the sample and suggest that the layer is late Floian (Early Ordovician) in age (Chen Zhongyang, personal communication 2023). Morphological terminology used herein follows that of the Treatise on Invertebrate Palaeontology (Whittington and Kelly 1997).
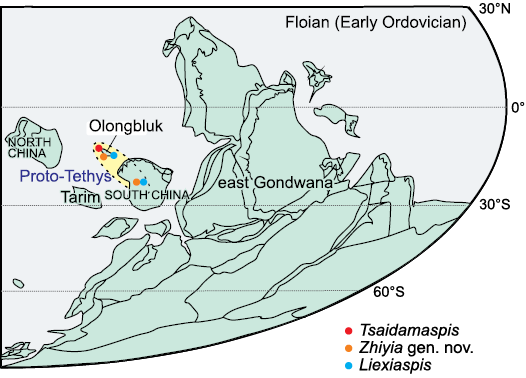
Fig. 2. Palaeogeographic map of the east Gondwana during the Floian (Early Ordovician) (modified from Torsvik and Cocks 2013, 2016; Cocks and Torsvik 2021), showing known occurrences of three genera reported from the Duoquanshan Formation.
Systematic palaeontology
Class Trilobita Walch, 1771
Order Asaphida Salter, 1864
Superfamily Asaphoidea Burmeister, 1843
Family Asaphidae Burmeister, 1843
Subfamily Isotelinae Angelin, 1854
Genus Tsaidamaspis Chang and Fan, 1960
Type species: Plesiomegalaspis (Tsaidamaspis) diarmatus Chang and Fan, 1960; Floian, Lower Ordovician, Duoquanshan Formation, northwestern Qinghai Province, China.
Emended diagnosis.—Cephalon is semicircular with genal spines. Glabella is subrectangular in outline, defined by deep axial furrows. There are five pairs of glabellar furrows and a V-shaped median furrow on the frontal glabellar lobe. Anterior border is broad (sag.) and flattened. Palpebral lobe is small. Anterior branches of facial sutures are divergent forward, curving inward at a rounded angle, then running rapidly forward and meeting at a pointed angle; posterior branches are long, extending posterolaterally. Hypostome is forked, with a broadly rounded median notch. Pygidium is triangular with a long (sag.) and narrow (tr.) axis. Pygidial border is declined laterally and extended posteriorly as a pair of short and stout posterior border spines (modified from Chang and Fan 1960: 123).
Remarks.—Tsaidamaspis was established by Chang and Fan (1960: 123) on the basis of the type species Tsaidamaspis diarmatus from the Duoquanshan Formation of Shihuigou, northwestern Qinghai Province, and regarded as a subgenus of Plesiomegalaspis Thoral, 1946. However, the latter was considered by Fortey and Owens (1987) to be a junior synonym of Asaphellus Callaway, 1877, which is distinctly different from Tsaidamaspis in having a much shorter preglabellar field, the hypostome with slightly concave and rounded posterior margin, and a semicircular pygidium without a pair of posterior border spines. Therefore, herein, we follow Lu et al. (1963: 39) and Zhou and Zhou (2008: 239) in giving Tsaidamaspis generic status. In addition, as noted by Laurie (2006) and Fortey and Bruton (2013), asaphids are difficult to classify due to major problem in the phylogeny. In this paper, we temporarily use the traditional classification and place Tsaidamaspis to subfamily Isotelinae.
The taxonomy of the Baltoscandian genus Megistaspis Jaanusson, 1956, and its allies is much complicated (see Nielsen 1995; Hoel 1999). Of them, Megistaspis (Paramegistaspis) Balashova, 1976, and Megistaspis (Ekeraspis) Tjernvik, 1956, are rather similar to Tsaidamaspis in cranidial morphology. Some species of M. (Paramegistaspis), such as M. (P.) planilimbata (Angelin, 1851) from the lower Floian and M. (P.) estonica (Tjernvik, 1956) from the upper Floian to lower Dapingian (Lower–Middle Ordovician), show the cranidium that has an obvious anterior border, the posterior margin of hypostome with a tongue-shaped median process or a shallow median notch, and the much shorter and subsemicircular pygidium without a pair of posterior border spines (see Tjernvik 1956: pl. 6), all of these features are clearly different from those of Tsaidamaspis species. Megistaspis (Ekeraspis) [see type species Plesiomegalaspis (Ekeraspis) armata Tjernvik, 1956: 242, pl. 7: 7–13] of the lower part of Latorp Formation (upper Tremadocian, Lower Ordovician) of Västergötland, Sweden differs from Tsaidamaspis in having less divergent forwardly anterior branches of the facial suture, rounded posterior margin of hypostome, and the pygidium with only one median spine.
Tsaidamaspis diarmatus Chang and Fan, 1960
Figs. 3, 4.
1960 Plesiomegalaspis (Tsaidamaspis) diarmatus subgen. nov. sp. nov.; Chang and Fan 1960: 123, pl. 6: 1–5, 8, non 6, 7.
1960 Isotelus tsinghaiensis sp. nov.; Chang and Fan 1960: 124, pl. 6: 12.
1963 Tsaidamaspis diarmatus Chang and Fan; Lu et al. 1963: 39, pl. 9: 3, 5, 6, non 4.
1965 Megalaspidella (Tsaidamaspis) diarmatus Chang and Fan; Lu et al. 1965: 497, pl. 100: 10, 11, 13, non 12.
1979 Megalaspidella (Tsaidamaspis) diarmatus Chang and Fan; Zhu et al. 1979: 111, pl. 45: 13–16.
2008 Tsaidamaspis diarmatus Chang and Fan; Zhou and Zhou 2008: 239.
Holotype: NIGP 10109, pygidium, see Chang and Fan (1960: 123, pl. 6: 4).
Type locality: Shihuigou, northwestern Qinghai Province, China.
Type horizon: Duoquanshan Formation (Floian, Lower Ordovician).
Material.—An incomplete cranidium, two librigenae, one hypostome, and four pygidia (NIGP 203156–203163), all from the type locality and horizon.
Description.—Cranidium is gently convex, with flattened anterior border, the width (tr.) of which is equal to about 70% of the cranidial length (sag.), and the length (sag.) of which occupies 30% of the cranidial length (sag.). Glabella is gently convex, and slightly narrowed at the level of S2, rounded anteriorly, 58% as wide as long. Preglabellar furrow is deepened laterally, medially shallow into the V-shaped median furrow on the frontal glabellar lobe. A prominent median tubercle is adjacent to occipital furrow. Bacculae are low, elongately oval in outline, and isolated by S1 and deep axial furrows. There are five pairs of shallow glabellar furrows; S1 extends rearwards and inward from the rear of constriction of glabella to connect at about 10% of occipital furrow; S2 is short (tr.) and suboval in outline, running backwards directed slightly; S3 is longer than S2 and almost transversely directed; S4 is long, suboval in outline, and forwards directed at 40° to the sagittal line; S5 is short and faint, extending forwards at 70° to the sagittal line. Axial furrow is deep, becoming relatively shallower forwards. Occipital ring is poorly preserved, about 10% of the glabellar length, defined by a broad (sag. and exsag.) and shallow occipital furrow. Palpebral lobe is small and semicircular, occupying about 10% of cranidial length (sag.). Anterior branches of facial sutures are divergent forward, curving inward at a rounded angle, then running rapidly forward and meeting at a pointed angle; posterior branches are long, extending posterolaterally. Palpebral area of fixigena is narrow (tr.) and strongly convex. Posterior area of fixigena is broad (tr.). Librigena is broad (tr.) and gently convex, with broad-based genal spine directed rearward and outward. Lateral border of librigena is flattened, narrowing rearwards. Posterior border of librigena is faintly defined. Eye is raised above the level of the rest of the librigena.
Hypostome is forked, with a broadly rounded median notch, about 80% as wide as long. Median body is oval, convex (tr.), and defined by shallow and broad lateral border furrows. Middle furrow is shallow and broad with a pair of ridge-like maculae. Lateral border widening posteriorly. Posterior border is broadly based, bluntly pointed posteriorly. Anterior wing is rounded triangular and sloping downwards laterally. Surface is covered with terrace lines subparallel to lateral and posterior borders.
Pygidium is triangular, about 93–96% as long as wide. Axis is long and conical, narrow and convex (tr.), tapering gradually rearwards, and defined by shallow axial furrows, occupying about 73–77% of the pygidial length and 24% of the anterior pygidial width. There are at least 20 axial rings and a rounded terminal piece, which are separated by shallow ring furrows. Articulating half ring is narrow (sag.), and rounded anteriorly. Pleural field is moderately convex, curving downwards laterally and posteriorly to border. Pleural field has at least 14 pleural furrows, which extend rearward and outward to border furrow. Triangular facets present at anterolateral corners of pygidium. Pygidial border is flattened and defined by faint border furrow. Pygidial border extended rearwards as a pair of short and stout posterior border spines, which occupy about 9% of the pygidial length. Surface of pygidium is covered with irregular lines.
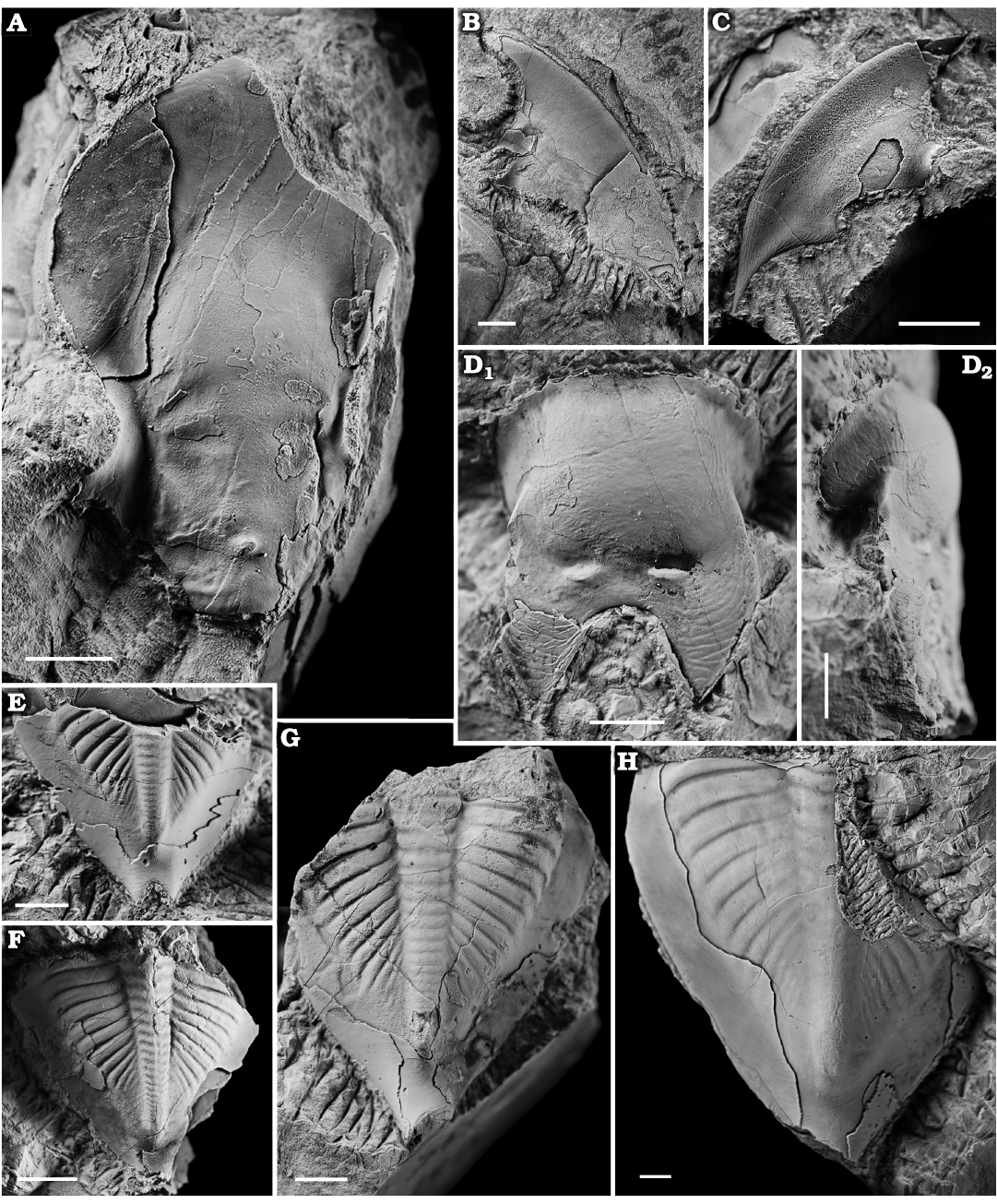
Fig. 3. Asaphid trilobite Tsaidamaspis diarmatus Chang and Fan, 1960, from the middle part of the Duoquanshan Formation (upper Floian, Lower Ordovician) of Shihuigou, Delingha, northern Qinghai Province, China. A. NIGP 203156, cranidium. B. NIGP 203157, librigena. C. NIGP 203158, librigena. D. NIGP 203159, hypostome in dorsal (D1) and lateral (D2) views. E. NIGP 203160, pygidium. F. NIGP 203161, pygidium. G. NIGP 203162, pygidium. H. NIGP 203163, pygidium. Scale bars 5 mm.
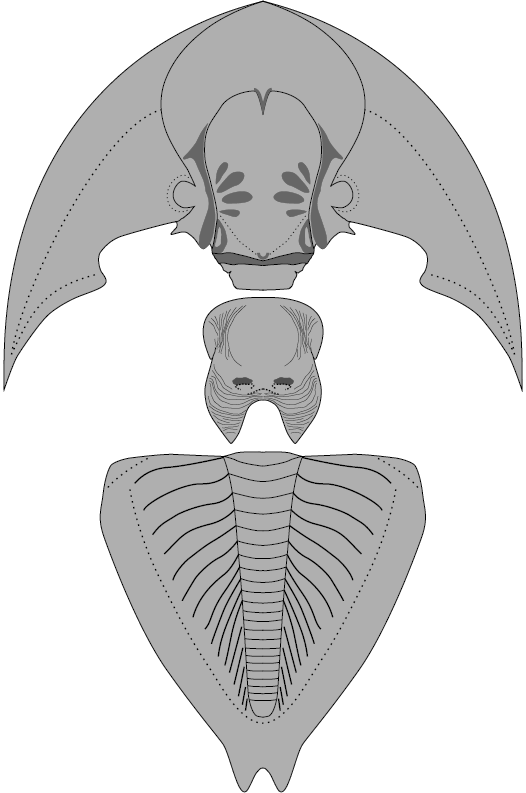
Fig. 4. Reconstruction of the asaphid trilobite Tsaidamaspis diarmatus Chang and Fan, 1960. Not to scale.
Remarks.—An incomplete hypostome figured by Chang and Fan (1960: 123, pl. 6: 6, 7) from the Duoquanshan Formation of Shihuigou, northwestern Qinghai Province, was assigned to Tsaidamaspis diarmatus by Chang and Fan (1960), and this view was followed by Lu et al. (1963: 39, pl. 9: 4; 1965: 497, pl. 100: 12). However, this hypostome exhibits a shallow median notch and a pair of rounded posterior borders, which agree well with species of the new genus Zhiyia (see below), and should be excluded from T. diarmatus. Another hypostome collected from the same horizon by Chang and Fan (1960: 124, pl. 6: 12, as Isotelus tsinghaiensis) is reassigned herein to T. diarmatus based on a deep median notch and forked posterior border.
Tsaidamaspis? sp. was recorded by Chang and Fan (1960: 124, pl. 5: 13, 14) based on one poorly preserved pygidium from the Floian strata (Lower Ordovician) from Yumen of western Gansu Province, showing the outline of triangular pygidium with a narrow (tr.) pygidial axis and a great number of axial rings and pleural furrows. As the specimen is poorly preserved, further materials are required to ascertain whether it ought to be assigned to Tsaidamaspis.
Stratigraphic and geographic range.—Floian, Lower Ordovician; Olongbluk terrane, NW China.
Genus Zhiyia nov.
Zoobank LSID: urn:lsid:zoobank.org:act:BEBEF30D-A413-4684-A27 E-62FCA94B7BBF
Etymology: In memory of the late Professor Zhou Zhiyi (1939–2022), of the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (NIGPAS), who have made great contributions to the research of trilobites in China.
Type species: Isotelus tsinghaiensis Chang and Fan, 1960; Floian, Lower Ordovician, Duoquanshan Formation, northwestern Qinghai Province, China.
Species included: Type species, Niobe (Niobella) obscura Zhou and Zhou, 2019. Zhiyia sp. nov. (i.e., Asaphid gen. et sp. indet. of Chen et al. 2023).
Diagnosis.—Isoteline genus with almost obsolete cephalic and pygidial axial furrows. Anterior border of cranidium is flattened and pointed forward, defined by obvious preglabellar furrow. Occipital ring is very narrow (sag. and exsag.). Hypostome is oval in outline, with a shallow median notch and a small triangular median projection between the bilobed posterior projections. Pygidium is subsemicircular in outline, with a broad (tr.) pygidial axis. Pygidial border is wide (sag.), defined by a broad and shallow border furrow. Surface of cephalic and pygidial borders is smooth or covered with transverse terrace lines.
Remarks.—The diagnostic features listed above are based mainly on the specimens of the late Floian (Early Ordovician) species Isotelus tsinghaiensis Chang and Fan, 1960, and Niobe (Niobella) obscura Zhou and Zhou, 2019, from middle Floian (Lower Ordovician), Madaoyu Formation, Liexi, western Hunan Province. The middle Floian Asaphid gen. et sp. indet. of Chen et al. (2023: 107–109, figs. 6–12) from the Madaoyu Formation of western Hunan Province is also considered to belong to the new genus on the basis of its almost obsolete cephalic and pygidial axial furrows, a flattened anterior border of the cranidium, a narrow occipital ring, an oval hypostome with a shallow median notch and bilobed posterior projections, and a subsemicircular pygidium with a wide border.
Two species belonging to the new genus were previously assigned to the genera Isotelus, Homotelus, and Niobe (Niobella) (see Chang and Fan 1960: 124; Zhou and Zhou 2008: 237; 2019: 156). However, Zhiyia differs from Isotelus Dekay, 1824, in having the cephalon more rounded forwardly, a shorter and subsemicircular pygidium with a shorter axis and wider border, and an oval hypostome with a longer anterior lobe and a shallower median notch with a small triangular median projection. Compared with Homotelus Raymond, 1920, the new genus has a flattened anterior border defined by obvious preglabellar furrow and a larger palpebral lobe, while the pygidium has almost obsolete axial furrows and a wide border defined by shallow border furrow. Isoteline Zhiyia is distinguished from Niobe (Niobella) Reed, 1931, and its allies of the Niobinae mainly by its almost obsolete cephalic and pygidial axial furrows, narrower (sag. and exsag.) occipital ring, shorter hypostome and the pygidium with wider axis.
With regard to the general outline and structure of the hypostome and pygidium, Zhiyia is very similar to Protopresbynileus Hintze, 1954 (= Pseudonileus Hintze, 1953) from the Fillmore Formation (Lower Ordovician) of western USA (see type species Pseudonileus willdeni Hintze, 1953: 224, pl. 15: 14–17), but is distinguished by the flattened anterior border of the cranidium and wider librigena having a flattened border, by the obvious pygidial border and lacking a median node on the pygidial doublure. In addition to Protopresbynileus, Ptyocephalus Whittington, 1948, of the Lower Ordovician from the Pogonip of Nevada, USA (see Ptyocephalus vigilans Whittington, 1948: 567, pl. 82: 1–7, pl. 83: 1–11) has a border to the cranidium as in Zhiyia, but the pygidium is pentagonal in outline with a longer axis.
Similar morphology of the hypostome and pygidium is also present in questionable species of Lachnostoma Ross, 1951, i.e., Lachnostoma platypyga Fortey and Bruton (2013: 101, fig. 39) from the Kirtonryggen Formation (Lower Ordovician) of Spitsbergen, which is easily distinguished from the new genus as it has a strongly waisted and longer glabella, a larger palpebral lobe, shorter (exsag.) posterior area of the fixigena, and a narrower (tr.) pygidial axis. However, L. platypyga is different from the type species Lachnostoma latucelsum in having a much larger, more anteriorly positioned palpebral lobe, a much less divergent anterior section of the facial sutures and a hypostome without marginal spines. Thus, this species may not be correctly assigned to Lachnostoma.
Fitzroyaspis irritans Fortey and Shergold, 1984, was established by Fortey and Shergold (1984) from the Nora Formation (Lower Ordovician) of central Australia, and then transferred by Laurie (2006: 309) to genus Eisarkaspis Laurie, 2006. Its cranidium as the holotype (see Fortey and Shergold 1984: 338, pl. 42: 2) is rather similar to that of Zhiyia in having almost obsolete cephalic axial furrows and the flattened anterior border, but other materials assigned to this species may be questionable because they are collected from different sections and horizons. Therefore, it is difficult to determine whether this species might be assigned to Zhiyia. In addition, Eisarkaspis from the Horn Valley Siltstone and basal Stairway Sandstone (upper Floian–middle Darriwilian) of Amadeus Basin (see type species E. etheridgei Laurie, 2006: 309, fig. 17–19) resembles Zhiyia in the cranidial and hypostomal morphology, but the former has a deeper lateral border furrow of the librigena, a hypostome without a small triangular median projection and a pygidium with narrower and longer axis as well as well-defined border furrow.
Stratigraphic and geographic range.—Floian, Lower Ordovician; Olongbluk terrane, South China palaeoplate.
Zhiyia tsinghaiensis (Chang and Fan, 1960)
Figs. 5, 6.
1960 Isotelus tsinghaiensis sp. nov.; Chang and Fan 1960: 124, pl. 6: 9–11, 13–15, non 12.
1960 Plesiomegalaspis (Tsaidamaspis) diarmatus subgen. nov. sp. nov.; Chang and Fan 1960: 123, pl. 6: 6, 7.
1963 Tsaidamaspis diarmatus Chang and Fan; Lu et al. 1963: 39, pl. 9: 4.
1965 Isotelus tsinghaiensis Chang and Fan; Lu et al. 1965: 488, pl. 97: 12–16, non 17.
1965 Megalaspidella (Tsaidamaspis) diarmatus Chang and Fan; Lu et al. 1965: 497, pl. 100: 12.
2008 Homalopyge tsinghaiensis (Chang and Fan); Zhou and Zhou 2008: 237.
Holotype: NIGP 10115, cranidium, Chang and Fan (1960: 124, pl. 6: 10); see Fig. 5A.
Type locality: Shihuigou, northwestern Qinghai Province, China.
Type horizon: Duoquanshan Formation, Floian (Lower Ordovician).
Material.—Three cranidia, one librigena, two hypostomes and seven pygidia (NIGP 203164–203176). All from the type locality and horizon.
Description.—Cranidium is gently convex (tr.), and its width between outer margins of palpebral lobes is about 87–91% of the sagittal length. Anterior border is flattened, bluntly pointed medially, and about 10% of cranidial length at the sagittal line, with transverse terrace lines. Elongated glabella is convex, broadly rounded and narrowing forwards, defined by obvious preglabellar furrow and shallower posteriorly axial furrows, about 78–88% as long as the width across the posterior ends of palpebral lobes. A prominent median tubercle is located at about 26% of the glabellar length (sag.) from the posterior. Axial furrows become shallower rearwards, and then almost totally effaced after the posterior ends of palpebral lobes. Four pairs of glabellar furrows are oval in shape, and shown as faint impressions on the exfoliated surface. S1 and S2 extend rearwards and inwards, and the latter opposite the level of the mid-length of the palpebral lobes; S3 and S4 directed forwards and inwards level with the anterior part of the palpebral lobes. Occipital ring is very narrow (sag. and exsag.) and slightly arched posteriorly, well separated from the rest of the glabella by a deep and broad (sag.) occipital furrow, and occupying about 5% of glabellar length (sag.). Palpebral lobe is gently convex, semicircular in outline, length (exsag.) equal to about 23–26% of the cranidial length (sag.), with the mid-length situated 40–42% of the cranidial length (sag.) from the posterior. Anterior branches of facial suture are divergent forward, curving inward at a rounded angle and meeting at a blunt angle in front; posterior branches extend posterolaterally. Posterior area of fixigena is long (exsag.), narrowing adaxially. Librigena is wide (tr.), gently convex (tr.), with rounded genal angle. Lateral border is flattened, defined by shallow lateral border furrow, with terrace lines parallel to margin. Posterior border furrow is effaced. Eye is semicircular and raised above the level of the librigena.
Hypostome is oval in outline, with a shallow median notch, about 95% as long as wide. Median body is oval, convex (tr.), and defined by shallow and broad (tr.) lateral border furrows and deep posterior furrow. Middle furrow is shallow and broad (sag.), with a pair of oval maculae situated at about 31% of the hypostomal length (sag.) from the posterior. Anterior lobe is subrectangular; posterior lobe is ridge-like, slightly arched anteriorly. Posterior border furrow is narrow (sag.), and slightly deeper than middle furrow. Lateral border occupies about 28% of hypostomal width. Posterior border has bilobed posterior projections and broadly based, rounded posteriorly. A small triangular median projection is situated between the bilobed posterior projections. Anterior wing is small and rounded. Surface of hypostome is covered with transverse lines.
Pygidium is subsemicircular in outline, about 58–67% as long as wide, with forwardly arched anterior margin. Axis is moderately convex (tr.), faintly defined by effaced axial furrows on exfoliated surface, occupying 46–53% of pygidial width at anterior margin, and 72–78% of pygidial length, tapering rapidly rearwards. Eight or nine rings are divided by axial ring furrows on exfoliated surface. Pleural field is gently sloping down towards border, with shallow anterior pleural furrow. Articulating facet is small and triangular in outline. Border is wide (sag.), about 21–28% of sagittal length of pygidium, sloping down towards margin, defined by a broad and shallow border furrow. Surface of pygidium is smooth, except for facet and border that are covered with approximately transverse terrace lines.
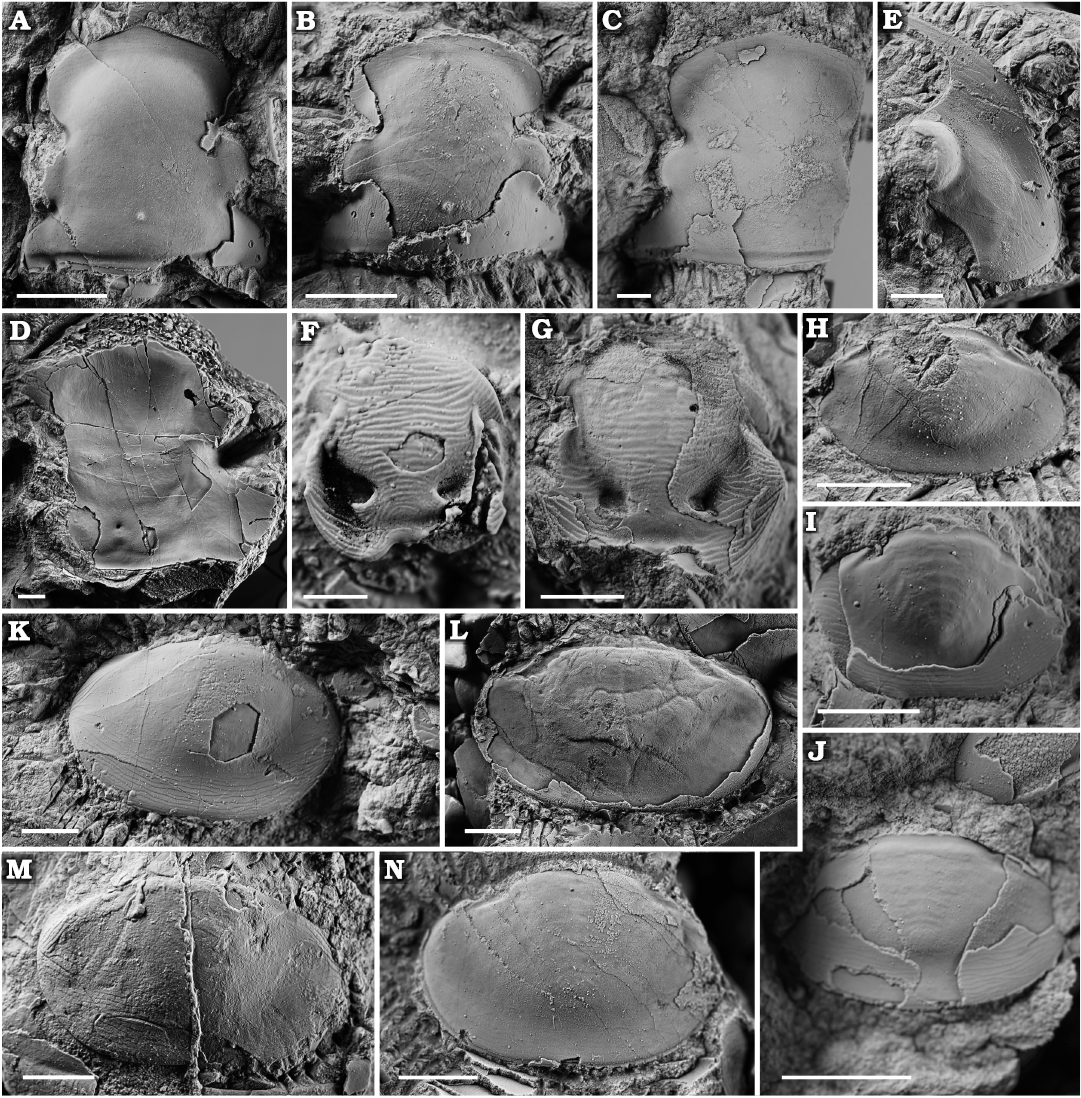
Fig. 5. Asaphid trilobite Zhiyia tsinghaiensis (Chang and Fan, 1960), from the middle part of the Duoquanshan Formation (upper Floian, Lower Ordovician) of Shihuigou, Delingha, northern Qinghai Province, China. A. NIGP10115, holotype, cranidium (refigured from Chang and Fan 1960: 124, pl. 6: 10). B. NIGP 203164, cranidium. C. NIGP 203165, cranidium. D. NIGP 203166, cranidium. E. NIGP 203167, librigena. F. NIGP 203168, juvenile hypostome. G. NIGP 203169, hypostome. H. NIGP 203170, pygidium. I. NIGP 203171, pygidium. J. NIGP 203172, pygidium. K. NIGP 203173, pygidium. L. NIGP 203174, pygidium. M. NIGP 203175, pygidium. N. NIGP 203176, pygidium. Scale bars: A–E, G–N 5 mm; F 2 mm.
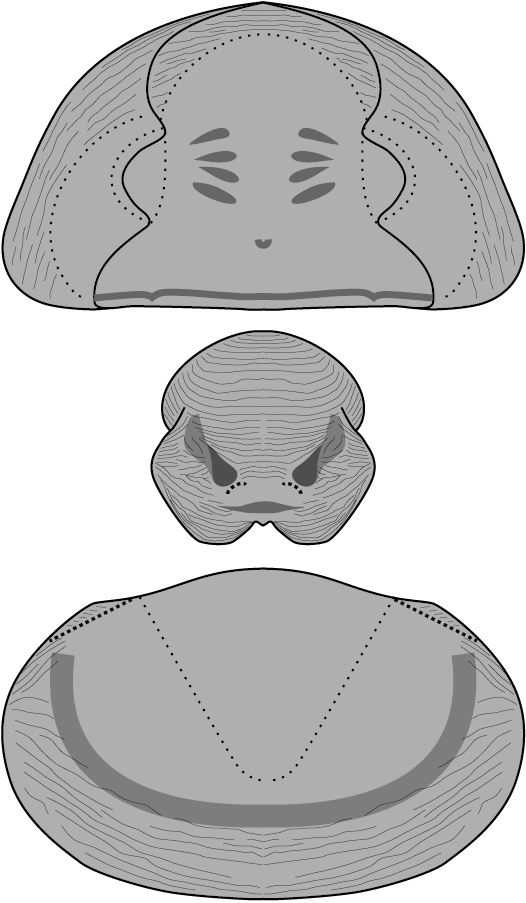
Fig. 6. Reconstruction of the asaphid trilobite Zhiyia tsinghaiensis (Chang and Fan, 1960). Not to scale.
Remarks.—Our materials are identical to specimens of Zhiyia tsinghaiensis (as Isotelus tsinghaiensis by Chang and Fan 1960: 124, pl. 6: 9–11, 13–15, non 12; see also Fig. 5A) from the same horizon and locality. It is very similar to Z. obscura (as Niobe (Niobella) obscura by Zhou and Zhou 2019: 156, fig. 5A–F) from the lower part of the Dawan Formation (upper Floian) of Yichang, western Hubei Province, but differs in having a pair of the more forwardly situated S1, smaller palpebral lobes, a relatively shorter pygidium, and obvious transverse terrace lines on the anterior border of the cranidium, the lateral border of the librigena and the pygidial border.
Of the Early Ordovician (middle Floian) species from Liexi, western Hunan Province, Asaphid gen. et sp. indet. collected from the Madaoyu Formation (see Chen et al. 2023: 107–109, figs. 6–12) is comparable with Z. tsinghaiensis particularly in its almost obsolete cephalic and pygidial axial furrows, but the former has a narrower cranidium, shallower preglabellar and occipital furrows, a longer pygidium, and smooth surface of the cranidium and pygidium. As is noted above it should be assigned to the new genus Zhiyia, and distinguished from all other described species, but until additional specimens become available, a new taxon is not formally introduced.
Stratigraphic and geographic range.—Floian, Lower Ordovician; Olongbluk terrane, NW China.
Order Phacopida Salter, 1864
Suborder Cheirurina Harrington and Leanza, 1957
Family Pliomeridae Raymond, 1913
Subfamily Protopliomeropinae Hupé, 1953
Genus Liexiaspis Liu, 1982
Type species: Liexiaspis zhengjiaensis Liu, 1982; upper Tremadocian, Lower Ordovician, Madaoyu Formation; Liexi, western Hunan Province, China.
Species included: Type species and Liexiaspis sp. of Fang et al. (2022).
Liexiaspis sp.
Fig. 7.
Material.—Two pygidia (NIGP 203177–203178) from the middle part of the Duoquanshan Formation (upper Floian, Lower Ordovician), Shihuigou section, northwestern Qinghai Province, China.
Description.—Pygidium is subhexagonal in outline, roughly as wide as long. Axis tapers rapidly rearwards, occupying about 64% and 45% of the pygidial length and the maximum width respectively; composed of four rings with articulating half-ring and terminal piece, which are well defined by axial ring furrows forwardly arched. Pleural region has four pairs of ribs separated by deep interpleural furrows, and becoming slightly narrower (tr.) and more posteriorly directed rearwards. Terminal piece is triangular, laterally enclosed by the fourth pair of ribs.
Remarks.—The present specimens agree well with the pygidium of the type species Liexiaspis zhengjiaensis Liu (1982: 339, pl. 238: 13) from the Madaoyu Formation (upper Tremadocian, Lower Ordovician) of western Hunan Province especially in the shape of the pygidial axis, and the number of the pleural ribs and axial rings. Herein, we prefer to regard our specimens as species indeterminate because of the lack of associated cranidia from the same horizon. Another species, Liexiaspis sp., figured by Fang et al. (2022: fig. S1-h) from the Madaoyu Formation (middle Floian, Lower Ordovician) of western Hunan Province differs clearly from the present form in having five axial rings and five pairs of pleural ribs.
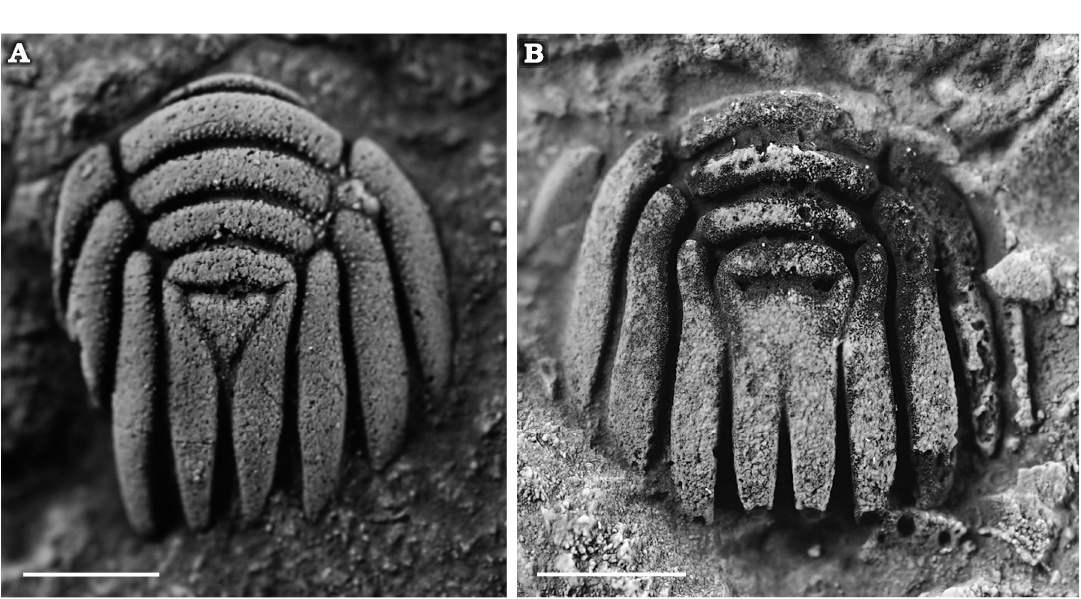
Fig. 7. Pliomerid trilobite Liexiaspis sp., from the middle part of the Duoquanshan Formation (upper Floian, Lower Ordovician) of Shihuigou, Delingha, northern Qinghai Province, China. A. NIGP 203177, pygidium. B. NIGP 203178, pygidium. Scale bars 2 mm.
Discussion
A low-diversity trilobite fauna with strong endemicity occurs on the shallow-water carbonate platform of the Olongbluk terrane during the Early Ordovician. As noted by Fortey (1975), Fortey and Owens (1978), Zhou et al. (1989, 2011, 2014) and Fortey and Cocks (2003), trilobites usually display decreasing endemicity from the shallow-water towards deep-water environments in the palaeoplate, while those shallow-water groups that are endemic to a particular palaeoplate have a higher value on defining biogeographic relationships.
The known palaeogeographical occurrences of the three genera are shown in Fig. 2. Tsaidamaspis Chang and Fan, 1960, as an endemic genus, only occurs in the Floian (Lower Ordovician) of Olongbluk terrane. Zhiyia gen. nov. has been reported from the Floian of the South China palaeoplate, i.e., the lower part of the Dawan Formation (upper Floian) of Yichang (Zhou and Zhou 2019, as Niobe (Niobella) obscura) and the Madaoyu Formation (middle Floian) of Liexi (Chen et al. 2023, as Asaphid gen. et sp. indet.). Liexiaspis Liu, 1982 ranges from late Tremadocian to Floian and occurs in South China palaeoplate (in Liu 1982 as L. zhengjiaensis and in Fang et al. 2022 as Liexiaspis sp.) and Olongbluk terrane (this paper).
Abundant nautiloid fossils of the Duoquanshan Formation were collected and described by Zhang (1965), Lai et al. (1984) and Lai (1985), those faunal evidences indicate that the Olongbluk terrane was near the North China palaeoplate during the Early–Middle Ordovician. However, the deep-water chitinozoan and graptolite faunas of the Shihuigou Formation suggest that the Olongbluk terrane was at low latitude and close to South China palaeoplate during the Middle Ordovician (Wang et al. 2018). Trilobite evidence is consistent with that of chitinozoan and graptolite faunas, suggesting that the Olongbluk terrane may have had a faunal link with the South China palaeoplate during the Floian (Early Ordovician) (Fig. 2). During the Middle–Late Ordovician, both the Olongbluk terrane and the South China palaeoplate would be interpreted as part of the Proto-Tethys Archipelagic Ocean (PTAO) (see Wei et al. 2023).
Conclusions
In this paper, we revise the late Floian, Early Ordovician trilobite fauna (three species of three genera) from the Olongbluk terrane, northern Qinghai Province, northwest China. The new isoteline genus Zhiyia is established based on new material from the Olongbluk terrane and South China palaeoplate. The fauna shows a strong endemicity to the Olongbluk terrane, and a close palaeogeographic relationship to coeval faunas of the South China palaeoplate during the Floian, Early Ordovician.
Acknowledgements
We thank Zhang Yuandong, Luo Hui, Zhu Youhua, Chen Zhongyang, Li Wenjie, Wu Xuejin and Hu Yunfeng (all NIGPAS) for help in the field, Chen Zhongyang (NIGPAS) for identifying conodonts, and Fang Xiang (NIGPAS) for valuable discussions. Special thanks are due to Alan W. Owen (University of Glasgow, Scotland, UK) and another anonymous reviewer for their constructive suggestions and critical reading of the manuscript. The research was financially supported by the Second Tibetan Plateau Scientific Expedition and Research Program (2019QZKK0706), the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB 26000000), the National Science Foundation of China (41672008, 41972011, 41972015, 42072005, 42072007 and 42030510) and the State Key Laboratory of Palaeobiology and Stratigraphy (Nanjing Institute of Geology and Palaeontology, CAS) (213123). This paper is a contribution to the IGCP 735 project “Rocks and the Rise of Ordovician Life”.
References
Angelin, N.P. 1851. Palaeontologia Scandinavica 1. Crustacea Formations Transitionis, Fasc 1. Palaeontologia Suecica 1851: 1–24.
Angelin, N.P. 1854. Palaeontologia Scandinavica 1. Crustacea Formations Transitionis, Fasc 2. i–ix + 21–92 pp. Samson and Wallin, Lund.
Balashova, E.A. [Balašova, E.A.] 1976. Sistematika Trilobitov Asaphina i ih Predstaviteli v SSSR. 215 pp. Nedra, Leningrad.
Burmeister, H. 1843. Die Organisation der Trilobiten aus ihren lebenden Verwandten entwickelt; nebst einer systematische Uebersicht aller zeither beschriebenen Arten. 147 pp. Reimer, Berlin. Crossref
Callaway, C. 1877. On a new area of Upper Cambrian rocks in South Shropshire, with a description of a new fauna. Quarterly Journal of the Geological Society of London 33: 652–672. Crossref
Chang, W.T. and Fan, J.S. 1960. Ordovician and Silurian trilobites of the Qilian Mountains [in Chinese]. Geological Gazetter of the Qilian Mountains 4 (1): 83–148.
Chen, H.M., Zhou, J., and Ding, L. 2023. The Liexi Fauna of the Early Ordovician in western Hunan Province [in Chinese]. Treasure 1: 99–113.
Cocks, L.R.M. and Torsvik, T.H. 2013. The dynamic evolution of the Palaeozoic geography of eastern Asia. Earth-Science Reviews 117: 40–79. Crossref
Cocks, L.R.M. and Torsvik, T.H. 2021. Ordovician palaeogeography and climate change. Gondwana Research 100: 53–72. Crossref
Dekay, J.E. 1824. Observations on the structure of trilobites, and description of an apparently new genus. Annales of the Lyceum of Natural History, New York 1: 174–189. Crossref
Fang, X., Mao, Y.Y., Liu, Q., Yuan, W.W., Chen, Z.Y., Wu, R.C., Li, L.X., Zhang, Y.C., Ma, J.Y., Wang, W.H., Zhan, R.B., Peng, S.C., Zhang, Y.D., and Huang, D.Y. 2022. The Liexi fauna: a new Lagerstätte from the Lower Ordovician of South China. Proceedings of the Royal Society B: Biological Sciences 289: 20221027. Crossref
Fortey, R.A. 1975. Early Ordovician trilobite communities. Fossils and Strata 4: 339–360. Crossref
Fortey, R.A. and Bruton, D.L. 2013. Lower Ordovician trilobites of the Kirtonryggen Formation, Spitsbergen. Fossils and Strata 59: 1–116. Crossref
Fortey, R.A. and Cocks, L.R.M. 2003. Palaeontological evidence bearing on global Ordovician–Silurian continental reconstructions. Earth Science Reviews 61: 245–307. Crossref
Fortey, R.A. and Owens, R.M. 1978. Early Ordovician (Arenig) stratigraphy and faunas of the Carmarthern district, south-west Wales. Bulletin of the British Museum (Natural History), Geology Series 30: 225–294.
Fortey, R.A. and Owens, R.M. 1987. The Arenig Series in South Wales. Bulletin of the British Museum (Natural History), Geology Series 41: 69–305.
Fortey, R.A. and Shergold, J.H. 1984. Early Ordovician trilobites from the Nora Formation, central Australia. Palaeontology 27: 315–366.
Harrington, H.J. and Leanza, A.F. 1957. Ordovician trilobites of Argentina. Special Publications, Department of Geology, University of Kansas 1: 1–276.
Hintze, L.F. 1953. Lower Ordovician Trilobites from Western Utah and Eastern Nevada. Utah Geological and Mineralogical Survey Bulletin 48: 1–249.
Hintze, L.F. 1954. Presbynileus and Protopresbynileus, new generic names proposed for Pseudonileus and Paranileus Hintze, preoccupied. Journal of Paleontology 28: 16–20.
Hoel, O.A. 1999. Trilobites of the Hagastrand Member (Tøyen Formation, lowermost Arenig) from the Oslo Region, Norway, Part I: Asaphidae. Norsk Geologisk Tidsskrift 79: 179–204. Crossref
Hupé, P. 1953. Classification des trilobites. Annals de Paléontologie 39: 61–168.
Jaanusson, V. 1956. Untersuchungen über baltoskandische Asaphiden. III. Über die Gattungen Megistaspis n. nom. und Homalopyge n. gen. Bulletin of the Geological Institution of Uppsala 36: 59–77.
Lai, C.G. 1985. Cephalopoda of the Duoquanshan Formation from Shihuigou, Ulan, Qinghai Province [in Chinese with English abstract]. Bulletin of the Institute of Geology Chinese Academy of Geological Sciences 12: 109–117.
Lai, C.G., Zhao, G.Z., and Xiang, L.W. 1984. New data from the Cambrian and Ordovician in the Shihuigou area, Ulan county, Qinghai Province [in Chinese with English abstract]. Geological Review 30: 311–318.
Laurie, J.R. 2006. Ordovician trilobites from the Horn Valley Siltstone and basal Stairway Sandstone, Amadeus Basin, Northern Territory. Memoirs of the Association of Australasian Palaeontologists 32: 287–345.
Liu, Y.R. 1982. Trilobita [in Chinese]. In: S.Q. Li (ed.), Palaeontological Atlas of Hunan. Ministry of Geology and Mineral Resources, Geological Memoir, Series 2. Stratigraphy and Palaeontology 1: 290–347.
Lu, Y.H., Chang, W.T., and Zhu, Z.L. 1963. Trilobita [in Chinese]. In: Nanjing Institute of Geology and Palaeontology, Academia Sinica (ed.), A Handbook of Index Fossils of Northwest China, 24–32, 35–39, 51, 57, 107. Science Press, Beijing.
Lu, Y.H., Chang, W.T., Zhu, Z.L., Chien, Y.Y., and Xiang, L.W. 1965. Trilobites of China [in Chinese]. 766 pp. Science Press, Beijing.
Mu, E.Z., Li, J.J., Ge, M.Y. and Lin, Y.K. 1962. Graptolites of Chilianshan [in Chinese]. 127 pp. Science Press, Beijing.
Mu, E.Z., Yu, C.M., Zhu, Z.L., Shi, C.G., Ge, M.Y., Yin, J.X. and Luo, J.D. 1963. The Ordovician of South Qilian Mountains [in Chinese]. Geological Record of the Qilian Mountains 2 (1): 115–128.
Nielsen, A.T. 1995. Trilobite systematics, biostratigraphy and palaeoecology of the Lower Ordovician Komstad Limestone and Huk Formations, southern Scandinavia. Fossils and Strata 38: 1–374. Crossref
Raymond, P.E. 1913. Notes on some new and old trilobites in the Victoria Memorial Museum, Canada Geological Survey (Ottawa). Bulletin of the Victoria Memorial Museum 1: 33–39. Crossref
Raymond, P.E. 1920. Some new Ordovician trilobites. Bulletin of the Museum of Comparative Zoology, Harvard University 64: 273–296.
Reed, F.R.C. 1931. A review of the British species of the Asaphidae. Annals and Magazine of Natural History 10: 441–472. Crossref
Ross, R.J. 1951. Stratigraphy of the Garden City Formation in northeastern Utah and its trilobite fauna. Bulletin of the Peabody Museum of Natural History 6: 1–161.
Salter, J.W. 1864. The British trilobites from the Cambrian, Silurian and Devonian formations, Part 1. Monograph of the Palaeontographical Society 16: 1–80. Crossref
Song, S.G., Niu, Y.L., Su, L., and Xia, X.H. 2013. Tectonics of the North Qilian orogen, NW China. Gondwana Research 23: 1378–1401. Crossref
Thoral, M. 1946. Conocoryphidae languedociens. Annales del’Université de Lyon, Sciences Naturelles 4: 5–74.
Tjernvik, T.E. 1956. On the Early Ordovician of Sweden. Stratigraphy and fauna. Bulletin of the Geological Institutions of Uppsala 36: 107–284.
Torsvik, T.H. and Cocks, L.R.M. 2013. New global palaeogeographical reconstructions for the Early Palaeozoic and their generation. In: D.A.T. Harper and T. Servais (eds.), Early Palaeozoic Palaeobiogeography and Palaeogeography. Geological Society, London, Memoirs 38: 5–24. Crossref
Torsvik, T.H. and Cocks, L.R.M. 2016. Ordovician. In: T.H. Torsvik and L.R.M. Cocks (eds.), Earth History and Palaeogeography, 101–123. Cambridge University Press, London. Crossref
Walch, J.E.I. 1771. Die naturgeschichte der verteinerungen, Dritter Theil. Zur erläuterung der Knorrischen Sammlung von Merkwürdigkeiten der Natur. 235 pp. P.J. Felstecker, Nürnberg.
Wang, W.H., Zhao, R., Muir, Lucy A., Li, M., and Tan, J.Q. 2018. Darriwilian (Middle Ordovician) chitinozoans from the Qaidam Paleoplate, northwest China. Review of Palaeobotany and Palynology 259: 123–133. Crossref
Wei, X., Luan, X.C., Zhang, Y.C., Yan, G.Z., and Zhan, R.B. 2023. Katian (Late Ordovician) trilobites of the North Qilian Mountains and their palaeogeographical implications for the Proto-Tethys Archipelagic Ocean (PTAO). Papers in Palaeontology 9 (6): e1532. Crossref
Whittington, H.B. 1948. A new Lower Ordovician trilobite. Journal of Paleontology 22: 567–572.
Whittington, H.B. and Kelly, S.R.A. 1997. Morphological terms applied to Trilobita. In: R.L. Kaesler, (ed.), Treatise on Invertebrate Paleontology, Part O, Arthropoda 1, Trilobita (revised). Volume 1, O313–O329. Geological Society of America, Boulder and University of Kansas, Lawrence.
Zhang, R.D. 1965. On some Lower Ordovician nautiloids from Qilianshan, northwestern China [in Chinese with English summary]. Acta Palaeontologica Sinica 13: 343–362.
Zhou, Z.Q. and Zhou, Z.Y. 2019. Ordovician trilobites from the lower part of the Dawan Formation (Floian–Dapingian) at Huanghuachang, Yichang, Hubei, southern China. Palaeoworld 28: 145–165. Crossref
Zhou, Z.Y. and Lin, H.L. 1995. Stratigraphy, Palaeogeography and Plate Tectonics of Northwest China [in Chinese]. 299 pp. Nanjing University Press, Nanjing.
Zhou, Z.Y. and Zhou, Z.Q. 2008. Latest Cambrian and Ordovician. In: Z.Y. Zhou and Y.Y. Zhen (eds.), Trilobite Record of China, 208–274. Science Press, Beijing.
Zhou, Z.Y., Bergström, J., Zhou, Z.Q., Yuan, W.W., and Zhang, Y.B. 2011. Trilobite biofacies and palaeogeographic development in the Arenig (Ordovician) of the Yangtze Block, China. Palaeoworld 20: 15–45. Crossref
Zhou, Z.Y., Yin, G.Z., and Zhou, Z.Q. 2014. Ordovician (Darriwilian–early Katian) trilobite faunas of northwestern Tarim, Xinjiang, China. Memoirs of the Association of Australasian Palaeontologists 46: 1–142.
Zhou, Z.Y., Zhou, Z.Q., and Zhang, J.L. 1989. Ordovician trilobite biofacies of North China Platform and its western marginal area [in Chinese with English summary]. Acta Palaeontologica Sinica 28: 296–313.
Zhu, X.H., Wang, H.L. and Xu, X.Y. (in press). Report on the Qilian Mountains, Regional Geology of China [in Chinese]. Geological Publishing House, Beijing.
Zhu, Z.L., Lin, H.L., and Zhang, Z.H. 1979. Trilobites [in Chinese]. In: Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, and Qinghai Institute of Geological Sciences (eds.), Palaeontological Atlas of Northwest China, Qinghai, 2, 81–116. Geological Publishing House, Beijing.
Acta Palaeontol. Pol. 68 (4): 683–693, 2023
https://doi.org/10.4202/app.01102.2023