
Novel pneumatic features in the ribs of the sauropod dinosaur Brachiosaurus altithorax
MICHAEL P. TAYLOR and MATHEW J. WEDEL
Taylor, M.P. and Wedel, M.J. 2023. Novel pneumatic features in the ribs of the sauropod dinosaur Brachiosaurus altithorax. Acta Palaeontologica Polonica 68 (4): 709–718.
Pneumatic dorsal ribs are known for many sauropods, but to date costal pneumaticity has received relatively little attention. In particular, the pneumatic ribs of the holotype specimen of Brachiosaurus altithorax have been largely overlooked, although they present a unique configuration of pneumatic features. One rib, with a pneumatic foramen some distance down the shaft, was briefly described and illustrated in the early 20th century by Elmer S. Riggs. A second rib with a pneumatic foramen in the tuberculum of the rib has not previously been described or illustrated. This previously undescribed foramen is similar in location to those in some dorsal ribs of Brontosaurus excelsus and Giraffatitan brancai, but differs from them in both size and shape. The contrasting sites of costal pneumaticity in the holotype individual of Brachiosaurus altithorax emphasize the generally opportunistic mode of postcranial pneumatization, in both sauropods and other ornithodirans, but conform to models of pneumatization following vascularization.
Key words: Sauropoda, Dinosauria, Brachiosauridae, pneumaticity, costal pneumaticity.
Michael P. Taylor [dino@miketaylor.org.uk; ORCID: https://orcid.org/0000-0002-1003-5675 ], Department of Earth Sciences, University of Bristol, Bristol BS8 1RJ, UK.
Mathew J. Wedel [mathew.wedel@gmail.com; ORCID: https://orcid.org/0000-0001-6082-3103 ], College of Osteopathic Medicine of the Pacific and College of Podiatric Medicine, Western University of Health Sciences, Pomona, California, USA.
Received 1 September 2023, accepted 13 November 2023, available online 29 December 2023.
Copyright © 2023 M.P. Taylor and M.J. Wedel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
In descriptions and analyses of fossil vertebrates, ribs have generally not been considered anatomically or phylogenetically important, and are often given only cursory treatment in even otherwise comprehensive descriptive work. In his three increasingly detailed descriptions of the giant sauropod dinosaur Brachiosaurus altithorax, Elmer S. Riggs (1901, 1903, 1904) had little to say about the ribs. Two of them, however, preserve interestingly different pneumatic features, which we describe, illustrate and discuss.
Anatomical nomenclature.—Some older authors, including Marsh and Riggs (e.g., Marsh 1896; Riggs 1903, 1904), refer to the head and tubercle of the rib. We use the now conventional terms capitulum and tuberculum respectively for these structures. Because the term “head” is also sometimes used informally to indicate the entire proximal portion of a rib, including both capitulum and tuberculum and the area in between them, we avoid this ambiguous term entirely and refer to the “proximal portion” when this is what we mean.
In life, the position and orientation of sauropod ribs were complex, and are still not fully understood. Broadly speaking, in all tetrapods with bicipital (two-headed) ribs, the ribs move during respiration as though rotating about an axis along the line from diapophysis to parapophysis (i.e., from capitulum to tuberculum on the rib itself). But because the extent and shape of articular cartilage on both the ribs and vertebrae of sauropods is unknown, it is not possible to make a precise assessment either of neutral position or range of motion.
Rather than attempt to discuss directions on ribs according to life position, then, we adopt the descriptive convention of a rib being oriented vertically. The proximal face of the rib that would in life have been oriented in some posteroventromedial direction is here considered to be posterior, so that the directions up and down the rib are proximal (towards the vertebra) and distal (towards the belly); and the directions across the shaft of the rib are medial and lateral (Fig. 1).
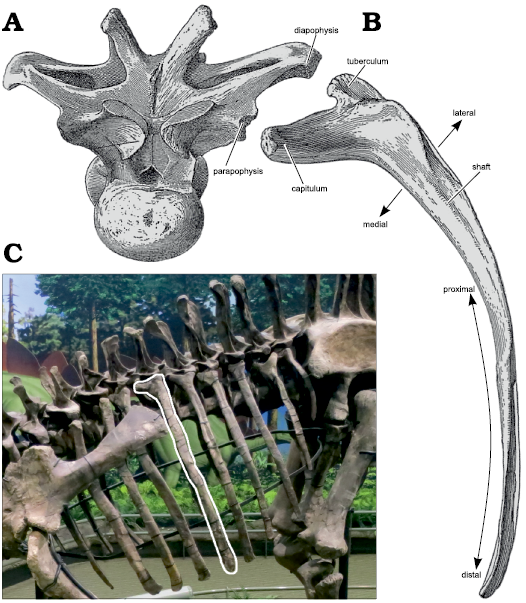
Fig. 1. Schematic illustration of a sauropod dorsal rib. A. Representative dorsal vertebra, in anterior view, with diapophysis and parapophysis labeled: these are the part of the vertebra that the rib articulates with; modified from dorsal vertebra 4 of Camarasaurus supremus Cope 1877, AMNH 5760’/D-X-131 in anterior view (Osborn and Mook 1921: pl. 70). B. Representative dorsal rib, shown in “anterior view” as described in the Anatomical Nomenclature section, with the capitulum, tuberculum and shaft labeled. The principal directions are illustrated: proximal towards the articulation with the vertebra and distal away from it along the shaft; medial towards the body core and lateral towards skin; modified from left rib 4 of Camarasaurus supremus AMNH 5761/R-A-24 in anterior view (Osborn and Mook 1921: fig. 71). C. The articulated rib cage of a mounted sauropod in left dorsolateral view with a single dorsal rib highlighted to emphasize that, due to the parapophyses being located more anteriorly than the diapophyses, the ribs do not lie in a plane perpendicular to the longitudinal axis of the torso; photograph by MPT of the mounted skeleton of the Apatosaurus louisae Holland 1915, holotype CM 3018 in the public gallery of the Carnegie Museum, in right dorsolateral view, reversed.
Institutional abbreviations.—CM, Carnegie Museum of Natural History, Pittsburgh, USA; FMNH, Field Museum of Natural History, Chicago, USA; Mal, Malawi Department of Antiquities Collection, Lilongwe and Nguludi, Malawi; MB, Humboldt Museum für Naturkunde Berlin, Berlin, Germany; OMNH, Sam Noble Oklahoma Museum of Natural History, Norman, USA; RRBP, Rukwa Rift Basin Project, Tanzanian Antiquities Unit, Dar es Salaam, Tanzania; SMM, Science Museum of Minnesota, Saint Paul, USA; PVL, Instituto Miguel Lillo, Collection of Vertebrate Paleontology, Tucumán, Argentina; USNM, United States National Museum of Natural History, Washington D.C., USA; YPM, Yale Peabody Museum, New Haven, USA.
Historical background
Brachiosaurus altithorax.—The giant sauropod dinosaur B. altithorax was very briefly described, with no name, in a preliminary report by Riggs (1901). It was then formally named, and described in slightly more detail by Riggs (1903). Riggs did not indicate a specimen number, but the holotype is FMNH PR 25107, and is held at the Field Museum of Natural History in Chicago, Illinois, USA.
The name that Riggs chose describes the animal’s morphology: “Brachiosaurus” means arm-lizard, in reference to the proportionally long forelimbs, and “altithorax” means high-torso, in reference to the “great size … of this specimen” (Riggs 1903: 299), presumably the length of the dorsal ribs in particular in light of “the immense size of the thorax” (Riggs 1903: 300). However, Riggs said little about the ribs in either of these publications. He followed these initial descriptions with a full descriptive monograph (Riggs 1904), but this too gives the ribs rather short shrift, describing them in a single paragraph (Riggs 1904: 239) of only 17 lines, which does not even specify how many were recovered, or from which side of the animal.
Riggs’s preliminary paper says only that “A complete rib, presumably from about the sixth presacral vertebra, measures more than nine feet in length. Some of the thoracic ribs have a secondary tubercle, and also a foramen leading to a cavity in the shaft.” (Riggs 1901: 549).
The formal description of the ribs is not much more informative (Riggs 1903: 303–304): “The unusual length of the ribs bears evidence of the immense thorax of this animal. In the mid-thoracic region they measure fully nine feet (2.745 m) in length. The capitulum and tuberculum are almost equally developed and widely separated, to give the firm anchorage necessary to the great length of the ribs. In some instances the attachment is strengthened by a second tubercle on the posterior surface of the head similar to that figured by Marsh [1896: 167] in the cervical ribs of Apatosaurus. The anterior surface of the shaft below the head is perforated by a large foramen which leads to an internal cavity in the shaft.”
Finally, the monographic description provides a little more detail (Riggs 1904: 239), along with some repetition. Here, we reproduce it in full: “The unusual length of the ribs, as well as the breadth of the head and tubercle and the strength of the shaft, bears evidence of the immense thorax of this animal. One of the more slender ribs from the mid-thoracic region measures fully nine feet (2.745 m) in length, Another has a shaft eight inches (0.204 m) in breadth. The head and tubercle are almost equally developed and widely separated to give the firm attachments rendered necessary by the great length of the ribs. In some instances the attachment is strengthened by a second tubercle on the inferior surface of the head similar to that figured by Marsh on the cervical vertebrae of Apatosaurus. The anterior surface of the shaft below the head is perforated by a large foramen which leads to an internal cavity. On account of the elevation of the capitular facet on the vertebra, the head and tubercle are borne almost on a level. By reason of this the flattened surface of the proximal end passes insensibly into the lateral surface of the shaft without that twist common to the ribs in animals of this group.”
The reference to “a second tubercle on the posterior surface of the head” (Riggs 1903) or “inferior surface” (Riggs 1904) is puzzling. We have not been able to identify any structure on any of the preserved ribs that persuasively matches the designation “second tubercle”, but we highlight in Figs. 2 and 3 candidate structures which Riggs could conceivably have been referring to.
But the illustration by Marsh (1896: 167, reproduced here as Fig. 4) is puzzling in its own right. It consists of his figs. 7 and 8, captioned as “Cervical rib of Apatosaurus ajax Marsh” with “outer” (i.e., lateral or anterior) view on the left and “inner” (i.e., medial or posterior) view on the right. But the three prongs identified in the figure are labeled “anterior extremity”, “head” (i.e., capitulum) and “tubercle” (i.e., tuberculum), with no rib shaft shown. The structure is extremely difficult to interpret as a cervical rib. A “posterior process” is shown in lateral view, which could possibly be construed as a “second tubercle”, but if it was on the lateral aspect of the rib it could not have served as an additional articulation. Furthermore, a third articulation for a rib would restrict the rib’s movement: possible in a cervical rib, but surely not in a dorsal rib, the purpose of which is to move in order to ventilate the respiratory system.
In summary, Riggs’s (1901, 1903, 1904) “second tubercle” is difficult to find on the Brachiosaurus ribs, and probably not homologous with whatever structure Marsh (1896) illustrated, which itself is difficult to interpret.
Janensch (1914) named a second Brachiosaurus species, B. brancai, based on material recovered from the Tendaguru Formation of Tanzania (then Deutsch-Ostafrika). This species is much better represented than B. altithorax, and a mounted skeleton based primarily on the referred specimen MB.R.2181 forms the spectacular centerpiece of the atrium of the Museum für Naturkunde Berlin (Janensch 1950a). As a result, the popular conception of Brachiosaurus has rested on this referred species. However, Paul (1988) showed that some significant differences exist between the two species and proposed that B. brancai be placed in a subgenus Brachiosaurus (Giraffatitan). This suggestion was not followed, but Taylor (2009) demonstrated that the two species are distinguished by at least 26 characters of the dorsal and caudal vertebrae, coracoids, humeri, ilia, and femora, and placed the African species in its own full genus as Giraffatitan brancai. This name is now in general use for Janensch’s (1914) Tanzanian species, and the name Brachiosaurus refers only to the American type species.
As discussed by Taylor (2009: 788–789), several additional North American specimens have been referred to Brachiosaurus altithorax, but none of these referrals can be made confidently due to a lack of overlapping material with the type specimen. D’Emic and Carrano (2019) tentatively referred the skull USNM 5730 and the Potter Creek postcranial material BYU 9754(4744)/USNM 21903 to B. altithorax, but did so only “based on lack of evidence for more than one brachiosaurid from the Upper Jurassic of North America” (D’Emic and Carrano 2019: 736). There is definitely evidence for multiple individuals of brachiosaurids in the Late Jurassic of North America (e.g., Maltese et al. 2018), but they are not necessarily referable to the species B. altithorax or even the genus Brachiosaurus. As a result, FMNH PR 25107 remains the only definitive specimen of Brachiosaurus at the time of writing.
Pneumaticity in sauropod ribs.—Among extant animals, crocodylians, birds, and mammals have pneumatic spaces in their skulls, but these are found in the postcranial skeletons of only one group: birds (e.g., Duncker 1971). Among extinct animals, postcranial skeletal pneumaticity (PSP) is more widely distributed, occurring in pterosaurs, theropod dinosaurs (including birds) and sauropodomorphs — but not ornithischian dinosaurs (e.g., Benson et al. 2012).
In sauropods, PSP is found most often in the vertebrae, where it is all but ubiquitous, but is also found less frequently in other bones including the scapulae, coracoids, ilia and chevrons (e.g., Cerda et al. 2012, Zurriaguz et al. 2017). Also among the sites of pneumatic features are the dorsal ribs. A variety of different features are found on different parts of ribs: principally the capitulum and tuberculum and the area between them. Such features are most often found on the posterior face of the rib but not infrequently on the anterior. Features include fossae (wide, shallow excavations), foramina (narrow, deep excavations leading to an internal air-space) and other possible traces of pneumatic diverticula pressed up against the ribs (Fig. 5).
As with pneumatic features in other bones, it is likely that the excavation of ribs by diverticula of the pulmonary system followed paths laid down by blood vessels, so that vascular foramina expanded to accommodate invasive diverticula and led to the development of pneumatic fossae and foramina (Taylor and Wedel 2021).
Description of Brachiosaurus ribs.—The preserved material of the Brachiosaurus altithorax holotype FMNH PR 25107 does not include any cervical ribs, or indeed any cervical material. Of the two caudal vertebrae, one is complete and includes the short and featureless caudal ribs that are fused to the centrum and neural arch, appearing as transverse processes of the vertebra. The dorsal ribs are the material of interest. Riggs (1901, 1903, 1904) did not give a count of these ribs, but as we shall see below, there are five of them.
Two of the ribs are preserved in much more detail than the others—in particular, they are the only ribs that preserve the capitulum and tuberculum. Their serial positions cannot be determined, beyond that they were not positioned either very anteriorly or very posteriorly within the trunk, so we arbitrarily designate these as Rib A and Rib B.
Rib A (Fig. 2).—This was illustrated by Riggs (1903: fig. 6) and slightly more informatively by Riggs (1904: pl. 75: 5). It consists of two pieces: a complete proximal end, and some but not all of the shaft. We interpret it as a right rib with the posterior aspect facing upwards in the jacket, based on the concavity of the available face and the curvature of the shaft. The rib is well preserved except for signs of reconstruction in the too-neat lamination of the tuberculum. It measures 53 cm across the capitulum and tuberculum.

Fig. 2. Sauropod dinosaur Brachiosaurus altithorax Riggs, 1903, holotype FMNH PR 25107 from Dinosaur Quarry No. 13 near Grand Junction, Colorado, dating to the Kimmeridgian–Tithonian ages of the Late Jurassic, right dorsal rib “Rib A” in posterior view with proximal to the left. A1, the whole proximal half of the rib; a distal portion also exists, of similar length but without features relevant to this study; A2, close-up of the tuberculum, highlighting the complex network of support structures that show signs of speculative reconstruction. Circles highlight two possible sites of the “second tubercle” referred to by Riggs (1901: 549, 1903: 303, 1904: 239) based on Marsh’s illustration (1896: figs. 7, 8), reproduced here in Fig. 4; A3, close-up of the pneumatic foramen in the shaft of the rib, showing natural bone texture around the margin and no indication of breakage. Scale bars provide only a rough indication of the size of the elements: see the text for measurements.
The rib’s most interesting feature is a small, oval pneumatic opening located about 60 cm down the shaft from the proximal end. It has been carefully prepared, and has finished bone inside: it is not a result of damage or an artifact of preparation. The opening measures 49 mm proximodistally and 25 mm mediolaterally. Its depth is 22 mm at both the medial edge and proximal end, and 18 mm at both the proximal edge and distal end. It is difficult to see the inner margins of the cavity. However, feeling around inside the opening, it seems likely that it extended further distally into the rib, although the possible extension has understandably not been prepared out. This is in agreement with Riggs’s (1903: 304–305) description: “The anterior surface of the shaft below the head is perforated by a large foramen which leads to an internal cavity in the shaft.” (Riggs 1903: 304, 1904: 239 both say that this opening is on the anterior face, and Taylor 2009: 792 followed his assessment, but herein we interpret this face as posterior.)
Rib B (Fig. 3).—This was not described or illustrated by Riggs, but it may be the rib measuring “fully nine feet (2.745 m) in length” that he refers to (Riggs 1903: 304, 1904: 239). Like Rib A, it consists of a well-preserved complete proximal end, and some but not all of the more distal portion. It is possible that some more distal part of Rib A or B has been lost, making up the full length of 9 feet that Riggs repeatedly cites. We interpret Rib B as a left rib with the posterior aspect upwards. It measures 56 cm across the capitulum and tuberculum.
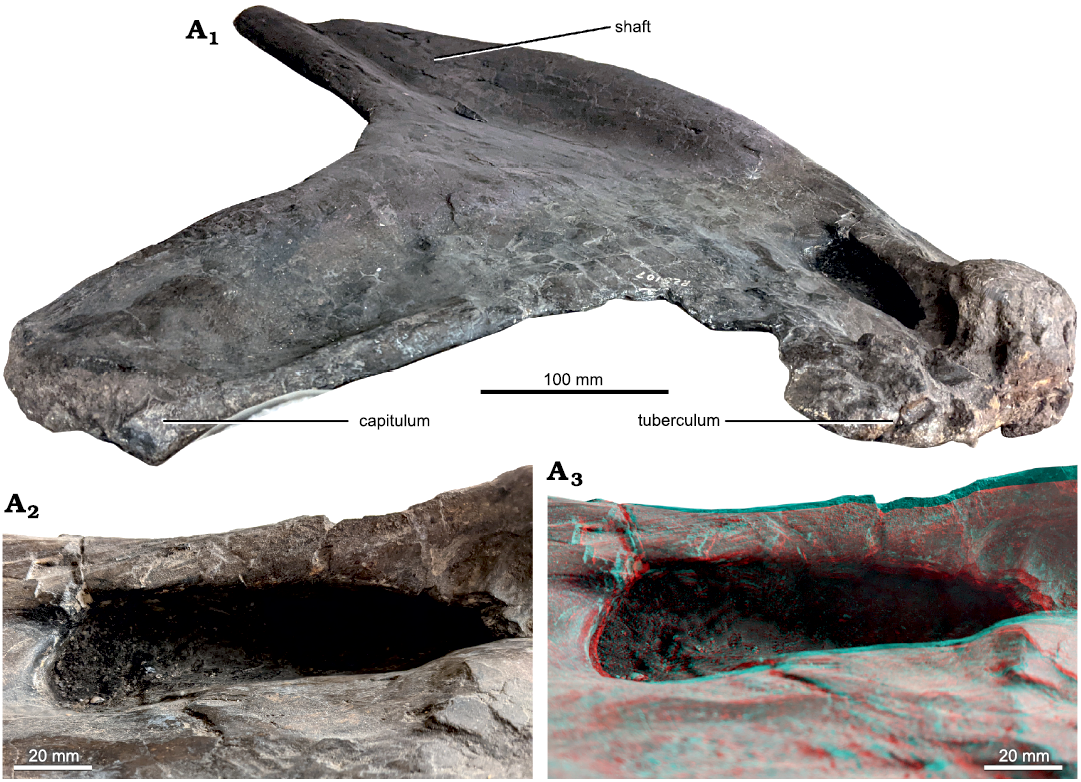
Fig. 3. Sauropod dinosaur Brachiosaurus altithorax Riggs, 1903, holotype FMNH PR 25107 from Dinosaur Quarry No. 13 near Grand Junction, Colorado, dating to the Kimmeridgian–Tithonian ages of the Late Jurassic, left dorsal rib “Rib B”. A1, the whole rib, posterior face in proximal view. Foreshortening makes the shaft look shorter and narrower than it actually is: the position of the rib between two shelves makes it impossible to photograph in true posterior view; A2, close-up of the pneumatic opening in the tuberculum in medial view, with anterior to the bottom; A3, red-cyan anaglyph of the same, indicating the form and depth of the fossa. Scale bars provide only a rough indication of the size of the elements: see the text for measurements.
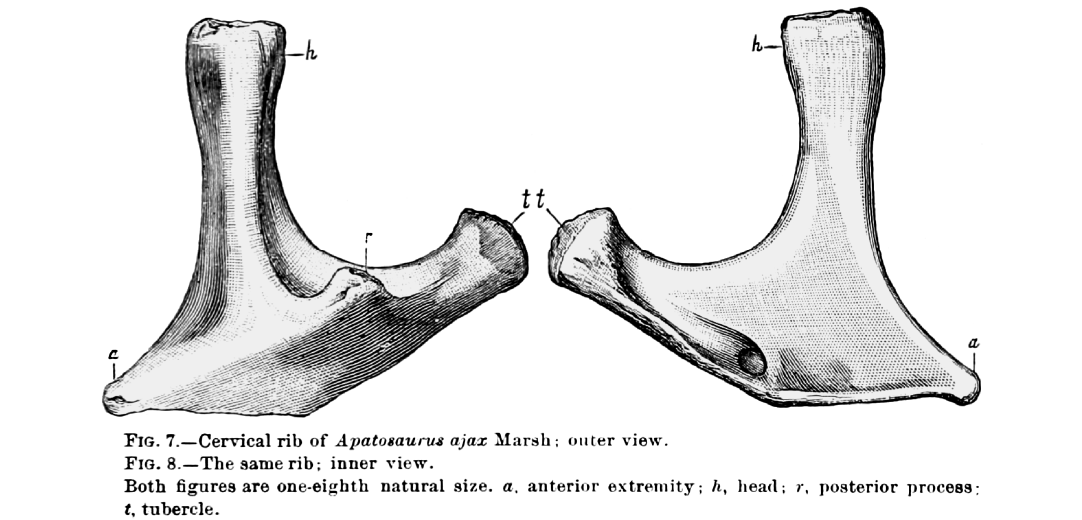
Fig. 4. A cervical rib of sauropod dinosaur Apatosaurus ajax Marsh, 1877 (specimen number unknown), as illustrated by Marsh (1896: figs. 7, 8), including the original caption. Note the “posterior process” marked as “r” in the illustration. This is probably the “second tubercle” referred to by Riggs (1901: 549, 1903: 303, 1904: 239), which he considered some part of one or more of the Brachiosaurus ribs to be homologous with.
The pneumatic opening in this rib is not described by Riggs (1903, 1904), though it was mentioned briefly by Taylor (2009: 792). It more closely resembles that documented by Marsh (1896: figs. 7, 8) in Brontosaurus excelsus (Fig. 5A) and those found in other sauropods, in that it invades the tuberculum rather than the shaft. Specifically, the lateral portion of the tuberculum is anteroposteriorly deeper than the medial part, projecting posteriorly from the surface of the rib, and the opening is in the medial face of this projection, extending laterally into the bone. The opening has been fully prepared out and is lined with finished bone. It is shaped like a teardrop flattened on one side, extending parallel to the rib shaft. It measures 120 mm proximodistally and has a maximum width of about 33 mm near its distal end: an exact measurement is impossible to determine because the medial margin of the opening merges smoothly onto the posterior face of the rib rather than ending in a lip, as the lateral margin does. The depth of the opening is about 50 mm towards its proximal end and 33 mm at its distal end. It is possible that the opening extended further proximally into the tuberculum.
Other ribs of the Brachiosaurus altithorax holotype.—Riggs’s (1901, 1903, 1904) descriptions mention Rib A and possibly Rib B, as discussed above. The collection contains three further ribs, for a total of five. Of these, one has only part of the proximal end, one only a section of the shaft, and one a partial proximal end and a broken-off more distal portion.
None of these ribs have visible pneumatic features. All of the ribs are large, heavy, and presumably fragile, and we have not attempted to move them from their supporting jackets. It is possible the hidden faces of these ribs preserve pneumatic features, but there is no particular reason to expect that they do.
Pneumaticity in ribs of referred Brachiosaurus specimens.—Jensen (1985, 1987) referred several specimens to Brachiosauridae, and tentatively to the species B. altithorax. These include at least one rib, probably three, although ambiguities in these papers make it uncertain what he intended: (i) Jensen (1987: fig. 1F) illustrates the proximal portion of a rib, but the caption does not specify what specimen or taxon it was considered to belong to. It is implied but not stated to be part of the Potter Creek brachiosaur, and appears to have a featureless surface. (ii) Jensen (1987: fig. 6B, also appearing as Jensen 1985: fig. 4B) shows a fiberglass resin cast of a “Jensen/Jensen quarry brachiosaur rib”, but the image contains almost no detail beyond the fact that is not the same rib as the one in fig. 1F of the same publication. (iii) Jensen (1987: fig. 8B) shows the proximal portion of another rib, visibly different from both the others, but the caption says “Supersaurus vivianae, right lateral view of mid-cervical vertebra”, which is obviously incorrect. The identity and assignment of this rib is therefore unknown.
There are no discernible pneumatic features on any of these ribs in Jensen’s (1985, 1987) illustrations, and Colin Boisvert (personal communication 2023) inspected these elements in collections and found no pneumatic features. These elements, together with all the brachiosaur material from the Dry Mesa and Jensen/Jensen quarries, are currently under restudy.
Pneumaticity in ribs of other brachiosaurids.—We are now in a position to survey occurrences of pneumaticity in the dorsal ribs of all known brachiosaurids:
– Brachiosaurus: Two different pneumatic features in two ribs in the holotype specimen, no other features recognised in type or referred specimens.
– Giraffatitan: Foramina in both anterior and posterior faces of the tuberculum in a single rib on MB.R.2181, but not apparently in any other rib.
– Cedarosaurus: No pneumatic features observed: two ribs “retain portions of the rib heads and clearly show that no pneumatic foramina was[sic!] present” (Tidwell et al. 1999: 25).
– Venenosaurus: A single 40 mm wide pneumatic foramen on the posterior surface of a right-sided rib leading proximally into a cavity in the capitulum (Tidwell et al. 2001: 153).
– Lusotitan: Although some dorsal rib fragments were excavated (Lapparent and Zbyszewski 1957), they were only briefly described and could not be located for the redescription of Mannion et al. (2013).
– Vouivria: “No rib heads are complete enough to determine whether the posterior surface was excavated” (Mannion et al. 2017: 37).
– Abydosaurus: No dorsal rib material was included in the specimens reported by Chure et al. (2010).
In summary, pneumatic features are known from the ribs only of Brachiosaurus itself (two ribs), Giraffatitan (one rib) and Venenosaurus (one rib), although see below on asymmetry of inference.
Discussion
Pneumatic sites in sauropod ribs.—The most common location of pneumatic features in the ribs of sauropods is the tuberculum, as seen in, among others, Brontosaurus excelsus (Fig. 5A) and Giraffatitan brancai (Fig. 5B). The pneumatic opening of Rib B of Brachiosaurus altithorax conforms to this pattern, although the shape and orientation of the opening is different from previously observed features. In particular, the invasion of bone in Rib B extends in a lateral direction and excavates a laterally positioned ridge on the posterior face of the rib, whereas other observed openings penetrate the bone anteriorly (from the posterior face) or posteriorly (from the anterior face).
The next most common location of pneumaticity is the space between the tuberculum and capitulum, as seen in Apatosaurus louisae (Fig. 5C) and Malawisaurus dixeyi (Fig. 5D), and in a different form in Brontomerus mcintoshi (Fig. 5E; Taylor et al. 2011: 84–85).
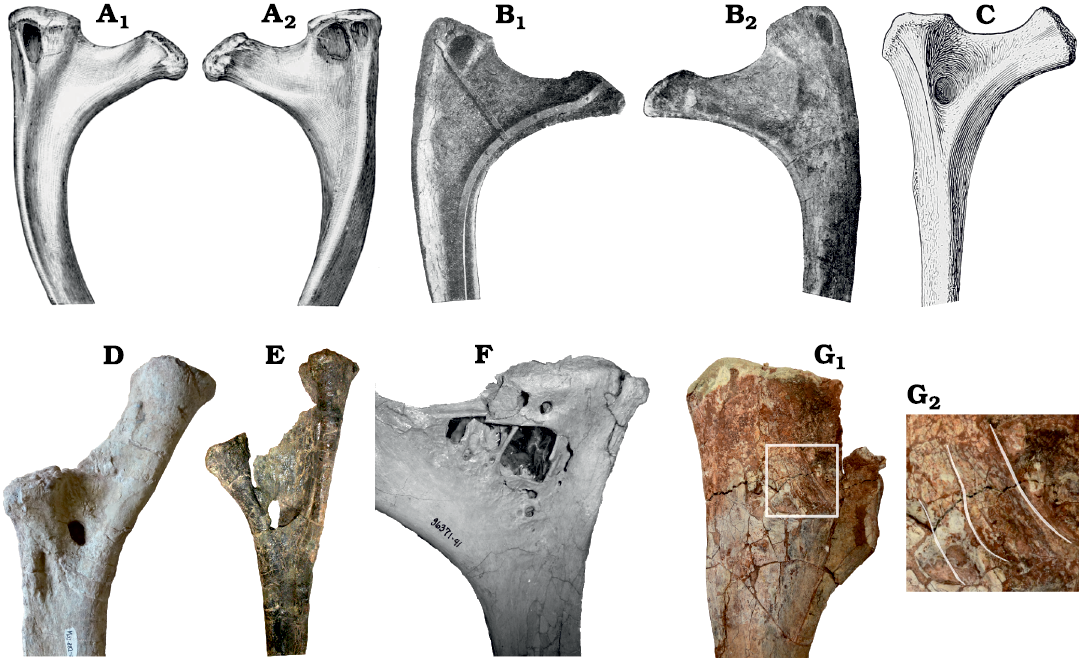
Fig. 5. Gallery of pneumatic features in a selection of dorsal ribs of sauropods, showing a range of pneumatic morphologies from most (A) to least (G) typical. A. Brontosaurus excelsus Marsh, 1879, specimen not indicated but likely the holotype YPM 1980 from Como Bluff, Wyoming; Kimmeridgian–Tithonian (Late Jurassic). A1, unspecified right dorsal rib in anterior view showing pneumatic fossa in tuberculum (modified from Marsh 1896: fig. 9); A2, same rib in posterior view showing a corresponding fossa in the tuberculum, traversed by an accessory lamina (modified from Marsh 1896: fig. 10). B. Giraffatitan brancai (Janensch, 1914), specimen not indicated but likely MB.R.2181 from Tendaguru, Tanzania; Kimmeridgian–Tithonian (Late Jurassic), that forms the core of the mounted skeleton in the atrium of the Museum für Naturkunde Berlin. B1, 2nd left dorsal rib in posterior view, showing pneumatic fossa in tuberculum (modified from Janensch 1950b: fig. 108); B2, same rib in anterior view, showing corresponding fossa in the tuberculum (modified from Janensch 1950b: fig. 107). C. Apatosaurus louisae Holland 1915, holotype CM 3018 from Dinosaur National Monument, Utah; Kimmeridgian–Tithonian (Late Jurassic), 2nd right dorsal rib in anterior view, showing pneumatic fossa between capitulum and tuberculum (modified from Gilmore 1936: pl. 29). D. Malawisaurus dixeyi (Haughton, 1928), Mal-282-2 from Karonga District, northern Malawi; Aptian (Early Cretaceous), left dorsal rib in posterior view, showing pneumatic foramen between capitulum and tuberculum, and fossa below capitulum. Photograph by Eric Gorscak. E. Brontomerus mcintoshi Taylor, Wedel, and Cifelli, 2011, OMNH 27766 from Grand County, eastern Utah; Aptian–Albian (Early Cretaceous), right dorsal rib 1 in posterior view, showing a narrow sheet of bone connecting capitulum and tuberculum and a pneumatic space entering the shaft in front of it. Photograph by MPT, used in Taylor et al. (2011: fig. 7). F. Rapetosaurus krausei Curry Rogers and Forster, 2001, SMM P2007.4.1 from Mahajanga basin, northwestern Madagascar; Maastrichtian (latest Cretaceous), dorsal rib, position and orientation unknown, showing a complex set of pneumatic features in the tuberculum and between it and the capitulum. Photograph by Kristina Curry Rogers. G. Rukwatitan bisepultus Gorscak, O’Connor, Stevens, and Roberts, 2014, holotype RRBP 07409 from Rukwa Rift Basin, southwestern Tanzania; Aptian–Cenomanian (Middle Cretaceous). G1, anterior ?left dorsal rib in posterior view; G2, close-up with highlights indication the locations of thin ridges described as a “capitulotubercular web” and interpreted as pneumatic by Gorscak et al. (2014: 1142–1143). Not to scale. Photograph by Eric Gorscak and Pat O’Connor.
Pneumatization of the capitulum is for some reason much rarer, and to date has only been recorded in Venenosaurus dicrocei (Tidwell et al. 2001: fig. 11.9).
Other pneumatic configurations also exist: for example, the complex of fossae and foramina in Rapetosaurus krausei (Fig. 5F) and the “pneumatic webbing” between the capitulum and tuberculum of Rukwatitan bisepultus (Fig. 5G; Gorscak et al. 2014: 1142). Similar “webbing” is also found in a less developed form in Mnyamawamtuka moyowamkia (Gorscak and O’Connor 2019: fig. 18).
The location of the foramen on Rib A of the B. altithorax holotype FMNH PR 25107 represents a unique location, appearing as it does some way down the shaft of a rib whose proximal portion appears devoid of pneumatic features. The closest approximation to this condition in another sauropod is perhaps in a right dorsal rib of the mamenchisaurid Xinjiantitan shanshanensis SSV 12001 (Zhang et al. 2022: fig. 14A), but the foramen in that case is located much more proximally than in Rib A.
All of these pneumatic fossae and foramina correspond to the seven-location schema of Wedel and Taylor (2023), which predicts that pneumatic features in costal elements would follow vascular foramina from the segmental and intercostal arteries.
The segmental arteries pass behind the ribs on their circuit of the centrum, providing channels for pneumatization of the posterior aspect of the proximal portion of the ribs, the tubercula and capitula and region between them. (Note that “posterior” here is really posteromedial, because the parapophysis is usually positioned anteroventral relative to the diapophysis rather than directly ventral, so that the rib is “folded back” against the torso.) The segmental arteries also less frequently vascularize and subsequently lead to pneumatization of the anterior aspect of the ribs of the next vertebra. Meanwhile, intercostal arteries extend along and beyond the length of the rib shaft, providing opportunities for vascularization and subsequent pneumatization.
However, while the pneumatization of the proximal portions of ribs, likely by diverticula following the segmental arteries, is relatively common in sauropods, pneumatization of the shafts, likely by diverticula following the intercostal arteries, is rare. Brachiosaurus Rib A provides the only documented occurrence.
Variability of pneumatic features.—There is no reason to suppose that vascularization of the vertebra that carried Rib A was any different from that of Rib B. (Or, if they are the left and right ribs of the same vertebrae, that this vertebra was vascularized differently on one side from the other.) Yet in following the segmental and intercostal arteries, the pneumatic diverticula in the region of these ribs did very different things. In Rib A, the proximal part of the rib, which is the only part pneumatized in most sauropod specimens, is entirely devoid of pneumatic features, yet a small, lipped foramen penetrates the shaft about 60 cm down. In Rib B, a broader, less well-defined pneumatic fossa is present in the lateral ridge on the posterior face of the tuberculum, and there are no discernable pneumatic features on the shaft.
Variability of pneumatic features in sauropod bones has been documented in the literature—differences between different species or specimens (e.g., McIntosh 1990), among successive vertebrae of a single individual (e.g., Diplodocus carnegii CM 82, Hatcher 1901), including bilateral asymmetry that may be consistent along the column (e.g., Saltasaurus loricatus PVL 4017, Zurriaguz and Alvarez 2014) or seemingly random (e.g., Giraffatitan brancai MB.R.2181, Wedel and Taylor 2013), and even asymmetry within a single vertebra (e.g., Xenoposeidon proneneukos NHMUK PV R2095, Taylor and Naish 2007). All of this is in accord with Witmer’s (1997: 64) conception of pneumatic diverticula as “opportunistic pneumatizing machines”, and similar variability in pneumaticity of ribs further corroborates this interpretation.
It is particularly odd that even in well-preserved specimens with complete or nearly-complete sets of ribs, pneumatic cavities are typically only present in one or two ribs in a given individual, examples include CM 3018, the holotype of Apatosaurus louisae (Gilmore 1936: pl. 29), and Giraffatitan brancai MB.R.2181 (Janensch 1950b: figs. 89–108). The quest to understand the evolution of postcranial pneumaticity in dinosaurs is already complicated by an asymmetry of inference: pneumatization of a single bone, such as a middle caudal vertebra of Giraffatitan brancai, is sufficient to demonstrate that diverticula of the respiratory system at least occasionally pneumatized that element in that taxon, but no number of apneumatic examples can prove the inverse (see discussion in Wedel and Taylor 2013). This problem becomes extreme in the case of dorsal ribs: if CM 3018 had a nearly complete set of dorsal ribs, missing only the right second rib, we would have no reason to suspect that dorsal rib pneumaticity was present in that specimen or in the species Apatosaurus louisae more generally; in fact, given a nearly full set of 19 apneumatic ribs, we might confidently but erroneously infer that dorsal rib pneumaticity was absent. We know of no antidote to this problem other than to keep documenting every available instance of dorsal rib pneumaticity.
Conclusions
FMNH PR 25107 is a well-known and even iconic specimen: not only was it described in some detail in the three papers by Riggs (1901, 1903, 1904) and revisited by Taylor (2009), it has provided scorings in multiple phylogenetic analyses, including those of Mannion et al. (2013) regarding the Portuguese brachiosaurid Lusotitan atalaiensis, and many subsequent analyses that use this matrix as a starting point. Furthermore, casts of the preserved material (the last seven dorsal vertebrae, sacrum, first two caudals, dorsal ribs, left coracoid, right humerus, ilium and femur) provided the core of a skeletal mount that was erected at the Field Museum in 1993 by Prehistoric Animal Structures, Inc. (Pridmore 1933; Carlozo 1993; Taylor 2014). It was the centerpiece of the museum until its removal in 1999 to make way for the Tyrannosaurus rex “Sue”. At this point it was transferred to Terminal 1 at O’Hare Airport on indefinite loan to United Airlines (Keown et al. 2000), where it remains to this day and is seen by sixty thousand people every day. It is striking that even such a well-known specimen, a hundred and twenty years after its description, can continue to provide unique anatomical features leading to novel insights.
Acknowledgements
We thank Bill Simpson (FMNH) for arranging a collections visit for MPT at very short notice. Eric Gorscak (Midwestern University Illinois campus, Downers Grove, USA) kindly supplied photographs of the ribs of Malawisaurus dixeyi; he and Patrick O’Connor (Ohio University, Athens, Ohio, USA) supplied the photograph of the rib of Rukwatitan bisepultus. Kristina Curry Rogers (Macalester College, Saint Paul, Minnesota, USA) supplied the photograph of the rib of Rapetosaurus krausei. We thank Colin Boisvert (UC Davis, Davis, California) for permission to cite a personal communication. Daniel E. Barta (Oklahoma State University, Stillwater, Oklahoma, USA) handled this manuscript efficiently, and it benefitted from three helpful and generous peer-reviews from Jerry Harris (Utah Tech University, St. George, Utah, USA), Virginia Zurriaguz (Universidad Nacional de Río Negro, Rio Negro Province, Argentina) and Pat O’Connor. We are grateful to them all for their help in improving this contribution.
References
Benson, R.B.J., Butler, R.J., Carrano, M.T., and O’Connor, P.M. 2012. Air-filled postcranial bones in theropod dinosaurs: physiological implications and the “reptile”-bird transition. Biological Reviews of the Cambridge Philosophical Society 87: 168–193. Crossref
Carlozo, L. 1993. Out of the Past. Chicago Tribune, 30 January 1993.
Cerda, I.A., Salgado, L., and Powell, J.E. 2012. Extreme postcranial pneumaticity in sauropod dinosaurs from South America. Paläontologische Zeitschrift 86: 441–449. Crossref
Chure, D., Britt, B.B., Whitlock, J.A., and Wilson, J.A. 2010. First complete sauropod dinosaur skull from the Cretaceous of the Americas and the evolution of sauropod dentition. Naturwissenschaften 97: 379–91. Crossref
Cope, E.D. 1877. On a gigantic saurian from the Dakota epoch of Colorado. Paleontology Bulletin 25: 5–10.
Curry Rogers, K. and Forster, C.A. 2001. The last of the dinosaur titans: a new sauropod from Madagascar. Nature 412: 530–534. Crossref
D’Emic, M.D. and Carrano, M.T. 2019. Redescription of brachiosaurid sauropod dinosaur material from the Upper Jurassic Morrison Colorado, Formation, USA. The Anatomical Record 303: 732–758. Crossref
Duncker, H.-R. 1971. The lung air-sac system of birds. Advances in Anatomy, Embryological and Cell Biology 45: 1–171. Crossref
Gilmore, C.W. 1936. Osteology of Apatosaurus with special reference to specimens in the Carnegie Museum. Memoirs of the Carnegie Museum 11: 175–300. Crossref
Gorscak, E. and O’Connor, P.M. 2019. A new African titanosaurian sauropod dinosaur from the middle Cretaceous Galula Formation (Mtuka Member), Rukwa Rift Basin, Southwestern Tanzania. PLOS ONE 14 (2): e0211412. Crossref
Gorscak, E., O’Connor, P.M., Stevens, N.J., and Roberts, E.M. 2014. The basal titanosaurian Rukwatitan bisepultus (Dinosauria, Sauropoda) from the middle Cretaceous Galula Formation, Rukwa Rift Basin, southwestern Tanzania. Journal of Vertebrate Paleontology 34: 1133–1154. Crossref
Hatcher, J.B. 1901. Diplodocus (Marsh): its osteology, taxonomy and probable habits, with a restoration of the skeleton. Memoirs of the Carnegie Museum 1: 1–63. Crossref
Haughton, S.H. 1928. On some reptilian remains from the Dinosaur Beds of Nyasaland. Transactions of the Royal Society of South Africa 16: 67–75. Crossref
Holland, W.J. 1915. A new species of Apatosaurus. Annals of the Carnegie Museum 10:143–145. Crossref
Janensch, W. 1914. Übersicht über der Wirbeltierfauna der Tendaguru-Schichten, nebst einer kurzen Charakterisierung der neu aufgeführten Arten von Sauropoden. Archiv für Biontologie, Berlin III 1 (1): 81–110.
Janensch, W. 1950a. Die Skelettrekonstruktion von Brachiosaurus brancai. Palaeontographica (Supplement 7) 3: 97–103.
Janensch, W. 1950b. Die Wirbelsaule von Brachiosaurus brancai. Palaeontographica (Supplement 7) 3: 27–93.
Jensen, J.A. 1985. Three new sauropod dinosaurs from the Upper Jurassic of Colorado. Great Basin Naturalist 45: 697–709. Crossref
Jensen, J.A. 1987. New brachiosaur material from the Late Jurassic of Utah and Colorado. Great Basin Naturalist 47: 592–608.
Keown, B. and Tribune staff writer. 2000. Replica of dinosaur fossil gives O’Hare passengers monstrous welcome. Chicago Tribune, 20.01.2000.
Lapparent, A.F. de and Zbyszewski, G. 1957. Les dinosauriens du Portugal. Mémoires des Services Géologiques du Portugal Nouvelle Série 2: 1–63.
Maltese, A., Tschopp, E., Holwerda, F., and Burnham, D. 2018. The real Bigfoot: a pes from Wyoming, USA is the largest sauropod pes ever reported and the northern-most occurrence of brachiosaurids in the Upper Jurassic Morrison Formation. PeerJ 6:e5250. Crossref
Mannion, P.D., Upchurch, P., Barnes, P.N., and Mateus, O. 2013. Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriforms. Zoological Journal of the Linnean Society 168: 98–206. Crossref
Mannion, P.D., Allain, R., and Moine, O. 2017. The earliest known titanosauriform sauropod dinosaur and the evolution of Brachiosauridae. PeerJ 5: e3217. Crossref
Marsh, O.C. 1877. Notice of new dinosaurian reptiles from the Jurassic formation. American Journal of Science 3 (14): 514–516. Crossref
Marsh, O.C. 1879. Notice of new Jurassic reptiles. American Journal of Science 18 (108): 501–505. Crossref
Marsh, O.C. 1896. The dinosaurs of North America. In: Extract from the 16th Annual Report of the U.S. Geological Survey, 1894–1895, Part I, 133–244. Government Print Office, Washington DC.
McIntosh, J.S. 1990 Species determination in sauropod dinosaurs with tentative suggestions for their classification. In: K. Carpenter and P.J. Currie (eds.), Dinosaur Systematics: Approaches and Perspectives, 53–69. Cambridge University Press, Cambridge. Crossref
Osborn, H.F. and Mook, C.C. 1921. Camarasaurus, Amphicoelias and other sauropods of Cope. Memoirs of the American Museum of Natural History, New Series 3: 247–387.
Paul, G.S. 1988. The brachiosaur giants of the Morrison and Tendaguru with a description of a new subgenus, Giraffatitan, and a comparison of the world’s largest dinosaurs. Hunteria 2 (3): 1–14.
Pridmore, J. 1993. Brach to the Future. Chicago Tribune, 2 July 1993.
Riggs, E.S. 1901. The largest known dinosaur. Science 13: 549–550. Crossref
Riggs, E.S. 1903. Brachiosaurus altithorax, the largest known dinosaur. American Journal of Science 15: 299–306. Crossref
Riggs, E.S. 1904. Structure and relationships of opisthocoelian dinosaurs. Part II, the Brachiosauridae. Field Columbian Museum, Geological Series 2 (6): 229–247.
Taylor, M.P. 2009. A re-evaluation of Brachiosaurus altithorax Riggs 1903 (Dinosauria, Sauropoda) and its generic separation from Giraffatitan brancai (Janensch 1914). Journal of Vertebrate Paleontology 29: 787–806. Crossref
Taylor, M.P. 2014. Gilles Danis of P.A.S.T on the Chicago Brachiosaurus mount. Sauropod Vertebra Picture of the Week, 29 May 2014. Crossref
Taylor, M.P. and Naish, D. 2007. An unusual new neosauropod dinosaur from the Lower Cretaceous Hastings Beds Group of East Sussex, England. Palaeontology 50: 1547–1564. Crossref
Taylor, M.P. and Wedel M.J. 2021. Why is vertebral pneumaticity in sauropod dinosaurs so variable? (version 5) Qeios 1G6J3Q.5. Crossref
Taylor, M.P., Wedel, M.J., and Cifelli, R.L. 2011. A new sauropod dinosaur from the Lower Cretaceous Cedar Mountain Formation, Utah, USA. Acta Palaeontologica Polonica 56: 75–98. Crossref
Tidwell, V., Carpenter, K., and Brooks, W. 1999. New sauropod from the Lower Cretaceous of Utah, USA. Oryctos 2: 21–37.
Tidwell, V., Carpenter, K., and Meyer, S. 2001. New titanosauriform (Sauropoda) from the Poison Strip Member of the Cedar Mountain Formation (Lower Cretaceous), Utah. In: D.H. Tanke and K. Carpenter (eds.), Mesozoic Vertebrate Life: New Research Inspired by the Paleontology of Philip J. Currie, 139–165. Indiana University Press, Bloomington.
Wedel, M.J. and Taylor, M.P. 2023. Blood vessels provided the template for vertebral and costal pneumatization in sauropod dinosaurs. Zenodo [available online, https://doi.org/10.5281/zenodo.10420716].
Wedel, M.J. and Taylor, M.P. 2013. Caudal pneumaticity and pneumatic hiatuses in the sauropod dinosaurs Giraffatitan and Apatosaurus. PLOS ONE 8 (10): e78213. Crossref
Witmer, L.M. 1997. The evolution of the antorbital cavity of archosaurs: a study in soft-tissue reconstruction in the fossil record with an analysis of the function of pneumaticity. Journal of Vertebrate Paleontology 17 (S1): 1–76. Crossref
Zhang, X.-Q., Li, N., Xie, Y., Li, D.-Q., and You, H.-L. 2022. Redescription of the dorsal vertebrae of the mamenchisaurid sauropod Xinjiangtitan shanshanesis Wu et al. 2013. Historical Biology [published online, https://doi.org/10.1080/08912963.2022.2147428]. Crossref
Zurriaguz, V.L. and Álvarez, A. 2014. Shape variation in presacral vertebrae of saltasaurine titanosaurs (Dinosauria, Sauropoda). Historical Biology 26: 801–809. Crossref
Zurriaguz, V., Martinelli, A., Rougier, G.W., and Ezcurra, M.D. 2017. A saltasaurine titanosaur (Sauropoda: Titanosauriformes) from the Angostura Colorada Formation (upper Campanian, Cretaceous) of northwestern Patagonia, Argentina. Cretaceous Research 75: 101–114. Crossref
Acta Palaeontol. Pol. 68 (4): 709–718, 2023
https://doi.org/10.4202/app.01105.2023