

Dimerelloid brachiopod Dzieduszyckia from Famennian hydrocarbon seep deposits of Slaven Chert, Nevada, USA, with insights into systematics and paleoecology of the Dimerelloidea
RUSSELL S. SHAPIRO
Shapiro, R.S. 2024. Dimerelloid brachiopod Dzieduszyckia from Famennian hydrocarbon seep deposits of Slaven Chert, Nevada, USA, with insights into systematics and paleoecology of the Dimerelloidea. Acta Palaeontologica Polonica 69 (1): 87–107.
This study describes an assemblage of Dzieduszyckia brachiopods preserved in sedimentary barite deposits from the Famennian, Upper Devonian of Nevada, USA. The brachiopods lived in a methane-seep environment like those described from Sonora, Mexico, and the Western Meseta, Morocco. Although the original carbonate fabrics are not preserved, pockets of limestone within the brachiopod barite are depleted in with δ13C values ranging from -31.1 to -27.3‰ VPDB. The fossils are preserved as internal and external molds only but record enough detail to demonstrate key internal features of Dzieduszyckia such as a pronounced dorsal septum, septalium, and vertical dental plates. The lack of common bifurcation or trifurcation of the costae as well as larger size of the shells recommends against inclusion within D. sonora as previously published. Based on the number of costae per width of shell, it is possible that there are two distinct species in Nevada, like other localities in Morocco, the Urals, and South China. These new data expand the body of knowledge of these earliest dimerelloids and it is hypothesized that this seep-dwelling macrofauna arose from depauperate-successful lineages between the Famennian biotic crises. As nearly all global Dzieduszyckia deposits are associated with active tectonic margins, it is also suggested that tectonically-driven methane expulsion in low nutrient waters spurred the evolution of dimerelloids.
Key words: Brachiopoda, Dimerelloidea, hydrocarbon seeps, chemosynthesis-based communities, Devonian, Nevada.
Russell S. Shapiro [rsshapiro@csuchico.edu; ORCID: https://orcid.org/0000-0003-1769-0346], Department of Earth and Environmental Sciences, California State University, Chico, USA.
Received 3 February 2024, accepted 27 February 2024, published online 27 March 2024.
Copyright © 2024 R.S. Shapiro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Methane seep ecosystems are special geobiological targets as the food web is founded on a diverse microbial consortium that links methane oxidation to sulfate reduction primarily in subsurface conditions (Case et al. 2015). The modern dominant macrofauna endemic to these chemically challenging environments have adapted to the low nutrition state by evolving symbiotic relationships with sulfide and/or methane oxidizing bacteria (e.g., Levin et al. 2010; Pereira et al. 2021). Paradoxically, most of the time interval for seeps, the Late Devonian through Early Cretaceous, approximately 240 million years, was macroscopically dominated by one superfamily of brachiopods, the Dimerelloidea (Campbell and Bottjer 1995a, b; Sandy 2010; Kiel et al. 2014; Kiel and Peckmann 2019; Baliński et al. 2022). This challenges conventional ecological wisdom as no modern brachiopods host symbionts, brachiopods at modern seeps are suspension feeders, and the bauplan of most brachiopods is not advantageous for hosting symbionts (Kiel and Peckmann 2019; however, see Angiolini et al. 2019 for a discussion of large brachiopods). Furthermore, it is important to note that bivalves likely analogous to extant chemosymbiotic forms are found locally in abundance in older late Silurian and Middle Devonian seeps in Morocco (see review in Jakubowicz et al. 2022).
Currently there is no geochemical evidence to unambiguously argue for symbiosis with methane or sulfide oxidizers in the rock record. While modern symbiotic mollusks have adapted their soft parts to maximize symbiosis and tissues record highly 13C depleted sources, those clues are not transferred to the fossil record (Ferrier-Pagès and Leal 2018). The shells themselves record no influence of methane or other hydrocarbons, with isotopic ratios of carbon recording sea water (e.g., Peckmann et al. 2007). Biomarkers of symbiotic bacteria cannot be isolated from forms free-living at the seep site.
Instead, paleontologists rely on general and yet unique characteristics of the skeletal record of the macrofauna. It has been long recognized that the macrofauna of seeps and hydrothermal vents tends to be dominated by high abundance and low diversity, typically monospecific assemblages (Campbell and Bottjer 1995a, b). These taxa often (at least with hydrothermal vents) show gigantism relative to other members of their clade (Van Dover et al. 2002). There are no current skeletal features that define obligate symbioses. Instead, researchers use a systematic approaching, linking fossil taxa to modern, seep or vent-obligate groupings. Lineages, such as the bivalve families Lucinidae and Vesicomyidae and gastropod subfamily Alviniconchinae, tend to be symbiotic and are frequently found at seeps or vents (see summaries in Amano et al. 2022; Kaim 2022). There may be other characteristics such as buffering from effects of background and mass extinctions (e.g., Danise and Higgs 2015) but more work is necessary to substantiate these claims.
Therein lies the paradox. One extinct lineage of brachiopods, the Dimerelloidea, (however, see discussion on Cryptoporidae below) dominated seeps from the Late Devonian through Early Cretaceous, are nearly always found at seeps (and one hydrothermal vent system), and are among the largest Mesozoic brachiopods. However, no modern brachiopods have symbionts to demonstrate this evolved lifestyle (Kiel and Peckmann 2019; Baliński et al. 2022). One group of putative dimerelloids, the Cryptoporidae, are not found at seeps but are likely not related to other dimerelloids (see fuller discussion below).
The purpose of this study is to provide new information on the Dzieduszyckia, the oldest dimerelloid, from massive barite deposits of Nevada, USA. While the new details on paleoecology, taxonomy, and taphonomy may not solve the challenge of recognizing symbiosis in the rock record, it is hoped the information may fuel future research and spur new avenues of inquiry.
Institutional abbreviations.—USNM, Smithsonian Institute National Museum of Natural History, Washington, D.C., USA.
Historical background
The Dimerellidae, at the rank of family, were defined by Buckman (1918) for a group of “Rhynchonellacea” brachiopods with small to absent deltidial plates, triangular foramen, dorsal medial septa, or largely developed crura. He further noted that each genus likely possessed some of the key characteristics but not all. The group was named after Dimerella Zittel, 1870, and originally included one other genus, Rhynchonellina Gemmellaro, 1871, and two potential genera. The family Dimerellidae was raised in rank to the superfamily Dimerelloidea in Savage (1996: 257, 260) and is the currently accepted grouping (e.g., Williams et al. 2000; Manceñido 2000; Manceñido and Owen 2001; Manceñido et al. 2002). Currently, the Dimerelloidea are included within the Order Rhynchonellida Kuhn, 1949 (Savage 1996; Savage et al. 2002).
As of this study, the Dimerelloidea range from the Late Devonian to the Early Cretaceous and are composed of twelve short-lived genera (e.g., Ager 1968; Sandy and Campbell 1994; Campbell and Bottjer 1995a, b; Gischler et al. 2003; Kiel and Peckmann 2008; Sulser and Furrer 2008; Peckmann et al. 2011, 2013; Kiel et al. 2014, 2021; Sandy and Peckmann 2016; Palfy et al. 2017), though perhaps the grouping is paraphyletic (Kiel et al. 2021; Guo et al. 2022) (Fig. 1). There are conflicting taxonomic organizational schemes in the literature, but most researchers define at least two branches: a more basal branch containing the family Peregrinellidae (Ager, 1965), and the more derived families Dimerellidae (Buckman, 1918) and Halorellidae (Ager, 1965). Manceñido and Owen (1996, 2001) included the family Cryptoporidae into the superfamily, suggesting the largely Cenozoic group is derived from the dimerelloid stock. They note that Buckman (1918) also recommended this inclusion. This inclusion is significant to the present paper as there are no known cryptoporids from seep or vent settings and nearly all of the other dimerelloids are exclusive to seeps and, in the case of Anarhynchia, a hydrothermal vent (Little et al. 2004).
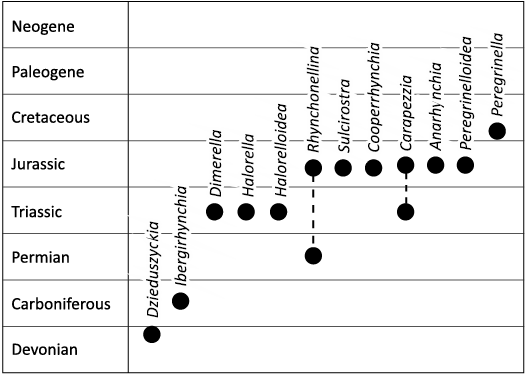
Fig. 1. Stratigraphic occurrences of dimerelloid brachiopod genera belonging to superfamily Dimerelloidea (Buckamn, 1918). Note that this arrangement is based solely on first appearance and does not imply evolutionary lineages. The Permian age of Rhynchonellina is based on Diener (1903) and Reed (1944).
However, in contradiction to the morphological assignment of taxonomic groups, Cohen and Bitner (2013) performed a phylogenetic analyses of small subunit (SSU/18S) and large subunit (LSU/28S) ribosomal DNA (rDNA) sequences from twelve rhynchonellide species, representing four extant superfamilies. Cryptopora gnomon, an extant cryptoporid and, thus, a dimerelloid sensu Manceñido and Owen (1996, 2001), was included in the study. The results defined three distinct clades in addition to the terebratulid and “inarticulate” outgroups. Cryptopora gnomon belongs to their Clade A1 which also includes species of Pugnacoidea, Hemithiridoidea, and Norelloidea. In fact, Cohen and Bitner’s (2013) study shows that genomic assessment of rhynchonellides is strongly incongruent with the morphological rankings, though a similar treatment of non-rhynchonellide brachiopods show quite good general agreement at higher taxonomic levels. Specifically, they draw attention to the Cryptoporidae as being controversial (Cohen and Bitner 2013: 215). Therefore, it is my opinion that the non-Cryptoporidae dimerelloids should be treated as a distinct, if paraphyletic grouping, of mostly hydrocarbon seep-obligate brachiopods.
The oldest genus of the dimerelloids is Dzieduszyckia. First noted by Georg G. Pusch in 1825 from the Kadzielnia quarry in Kielce, Góry Świętokrzyskie (Holy Cross Mountains) of Poland (Pusch 1833), the fossils were assigned to Terebratula amphitoma (Bronn, 1832) by Buch (1834). Unfortunately, in this publication, Buch included an unrelated Carboniferous specimen possessing a spiral brachidium. Roemer (1866: 671) recognized that the specimens figured by Buch (1834) were not T. amphitoma and reassigned the lot to a new species, Terebratula (?) kielcensis.
The species was formally redefined and named Dzieduszyckia by Siemiradzki (1909). Siemiradzki’s publication included the unrelated specimen, confounding taxonomists until the careful sleuthing of Biernat (1967) who detailed the error. In that critical manuscript, Biernat (1967) gave a thorough diagnosis as well as a comparison of all known occurrences up to 1967. The emended and current diagnosis for Dzieduszyckia focuses on the large biconvex shell with both valves sulcate; variably transverse form; anterior zigzag commissure; asymmetrical shell; radial costae, simple or dichotomous; prominent ventral beak, slightly incurved; small teeth with thin dental plates; dorsal septum; and thin, well-developed crura. The Late Devonian genus has a global yet spotty distribution including Poland, Morocco, Russia (North and South Urals), Kazakhstan, south China, Tajikistan, Mexico, and the USA (Fig. 2).
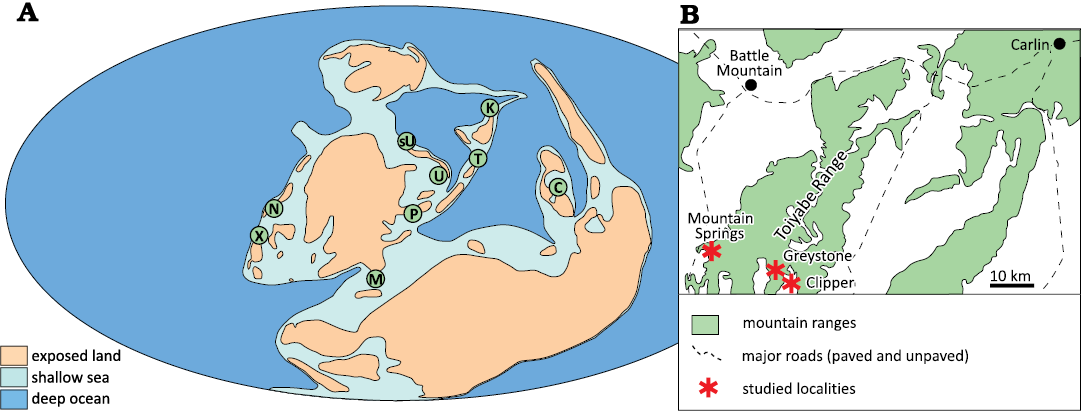
Fig. 2. A. Global distribution of dimerelloid brachiopod Dzieduszyckia in a reconstruction of Late Devonian paleogeography. B. Map showing studied Famennian localities in Nevada, USA. Abbreviations: C, China; K, Kazakhstan; M, Morocco; N, Nevada; P, Poland; sU, Subpolar Urals; U, Urals; T, Tajikistan; X, Mexico. Inset shows localities noted in the text. Image based on data and concept in Baliński and Biernat (2003: fig. 1) who used the reconstruction of Golonka et al. (1994). The position of Tajikistan is not well constrained.
Dzieduszyckia is found in high abundance, low diversity populations. Most dimerelloids, such as Halorella and Anarhynchia, comprise 99–100% of the macrofossil record in monospecific assemblages (e.g., Gischler et al. 2003; Peckmann et al. 2011; Palfy et al. 2017). At the same time, in seeps from Silurian to the Early Cretaceous, mollusks are common and diverse at some seep deposits, and they may or may not co-occur with some dimerelloid brachiopods (see taxonomic review list and references in Hryniewicz 2022).
In this regard, Dzieduszyckia is unique in that it has been found in samples with multiple species and more diverse fauna. At Sidi Amar, Western Meseta, Morocco, Dzieduszyckia is found as four species based on shell morphology (Termier and Termier 1948), emended to three species by Baliński and Biernat (2003). Analysis by subsequent authors have upheld the taxonomic splitting, though only two species, Dzieduszyckia crassicostata and Dzieduszyckia tenuicostata, are clearly distinct and common (e.g., Peckmann et al. 2007). The original taxonomic paper from South China also defined four species (Xian and Jiang 1978) though further analysis by Nie et al. (2016) argued for morphologic plasticity and narrowed the collections to one species. Biernat (1967) in her invaluable critical review of Dzieduszyckia noted similar variation in the Polish specimens from the Ruda Strawczyńska borehole but argued for environmental plasticity over taxonomic splitting. Additional details are provided below in the Systematic palaeontology section. Regardless, the data show that the variation between specimens of Dzieduszyckia at the outcrop scale exceeds those of other dimerelloids (e.g., Termier and Termier 1948; Nie et al. 2016). Also, it is common for Dzieduszyckia to be found amongst other Late Devonian fauna such as cephalopods, crinoids, and trilobites (e.g., Peckmann et al. 2007)
Summary of occurrences.—Poland: Dzieduszyckia kielcensis was first erected as the type species by Siemiradzki (1909) from material from a ca. 1825 collection by Pusch from the Kadzielnia quarry in Kielce, Holy Cross Mountains, in Poland (Pusch 1833; Roemer 1866; Biernat 1967). The original horizon has never since been relocated. Biernat (1967) was able to access material from a borehole at Ruda Strawczyńska near Kielce. The boring intersected a highly fossiliferous layer, similar to the original quarry, though other details are lacking. Unfortunately, there are no additional details on the facies, petrofabrics, or geochemistry beyond noting the brachiopods were from a layer of crystalline limestone with subordinate marl and clay (Pawłowska and Pawłowski 1978).
The general geology of the region of the Kadzielniańskie Range (the Holy Cross Mountains) is well understood. Devonian deposits comprise belts starting with intrashelf carbonate facies on the east and west separated by shallow water carbonates around Kielce.
Russia (Urals): The occurrence of Dzieduszyckia from the Ural Mountains has been noted for over a hundred years. The first mention was of the naming of Leiorhynchus bashkiricus Tschernyschew, 1887. Nalivkin (1947) noted L. bashkiricus from the western slopes of the Southern Urals and Rozman (1962) transferred the species to the genus Dzieduszyckia. Markovsky (1937) had listed the brachiopod as Nudirostra bashkirica, found with Leiorhynchus ursus in the Marzakayev (also transliterated at Murzakaevo and Murzakayev) beds of the Southern Urals (reported in Rozman 1960; see Sartenaer 1959). Two additional species, Dzieduszyckia prima Mizens, 2008, and Dzieduszyckia cordiformis Mizens, 2006, have also been recognized in the Southern Urals alongside Dzieduszyckia bashkirica.
Dzieduszyckia bashkirica (also erroneously listed as D. baschkirica or D. baschkiricus) has been used as a rare but valuable correlative fossil along with L. ursus for the “Famennian III” interval which equates to the Prolobites Ammonoid Zone (e.g., Baranov and Sartenaer 1996). The distribution of Dzieduszyckia in the Dzerzhinka section of the eastern slope of the Southern Urals was recently summarized in Gatovsky et al. (2017), noting that both D. bashkirica and D. cordiformis are confined to the upper half of the Murzakaevian (lower Trachytera Zone, upper Famennian). Mizens et al. (2003) noted Dzieduszyckia from the western slopes in the Zilair Synclinorium, west of Magnitogorsk in the Chelyabinsk Oblast. Additional locality details were recently summarized in Zhuravlev and Sokiran (2020) who also listed D. bashkirica in the Tchernyshev Swell along the eastern part of the Pechora Platform (Bolshaya Synya River, Sher-Nyadelta River) and Subpolar Urals (Kozhym River, Syvyu River) (Yudina 1997; Yudina et al. 2002; Tsyganko 2011). The authors also drew the connection to the Murzakaevian regional horizon of the western slope of the Southern Urals (e.g., Tagarieva 2012). Mizens and Mizens (2017) in their article on brachiopod distributions expanded the range of Dzieduszyckia downward into the Makarovian (lower to lower–middle Famennian) horizon in the west but restrained to the Murzakaevian on the eastern slope.
In general, the Famennian formations of the Western Urals (e.g., Zilair Synclinorium) are organic-rich and immature mudstones that were deposited in tectonically active back-arc rift and other basins (Gorozhanina et al. 2010). These deposits show intensive folding and overthrusting as a result of late Famennian tectonic collision (Ivanov and Puchkov 2020). The Eastern Urals at Dzherzhinka also record deep water deposition as a series of carbonate turbidites, perhaps on a deep-water ramp (Gatovsky et al. 2017). Similarly, Yudina et al. (2002) summarized the sequence in the Subpolar Urals as comprising gray to dark-gray, thin- to thick-bedded limestone, marls, and marly shales. There are also siliceous limestone with black chert nodules and lens-like nodular limestones. In general, the Subpolar Urals may be reconstructed as a series of isolated carbonate platforms rimmed by tectonically-driven subsidence basins (Gruzdev 2021).
Kazakhstan: The main record of Dzieduszyckia bashkirica comes from the Mugodzhar (= Mugudzhary) Hills in Kazakhstan (Rozman 1962) as well as the Zajsan-Irtysh region, but no additional details are provided for the latter. Rozman (1960) noted that Nudirostra (Dzieduszyckia) bashkirica is found in the Bakay Trough (north-northwest of Berchogur) within the cephalopod limestones of the Prolobites Zone.
The Famennian deposits are found in the Berchogur syncline along the western margin, overlying the Frasnian deposits (Rozman 1960). The entire sequence starts off with volcanic-rich Lower Devonian units and ends with nearly a kilometer of fine siliciclastic and carbonate beds (Nikolaeva et al. 2022). Rozman (1960) placed the brachiopods within “sandy limestone” equivalent to the Murzakaevian of the Urals. In many reconstructions, the units of the Mugodzhar Hills were formed on the edge of a microcontinent. Not long after deposition or perhaps concurrent, the region was deformed by major subduction or obduction as recorded in the eastern Mugodzhar zone (Golionko and Ryazantsev 2021).
Tajikistan: Dzieduszyckia bashkirica was reported by Menakova (1991) from the Shishkat section of the northern Zeravshan Range (Tien Shan) in central Tajikistan, near the border with Uzbekistan. This section has been well studied for nearly 100 years for its excellent and continuous middle Paleozoic sequence (Markovsky 1937; Sabirov 2019). The stratigraphic position was not noted by Menakova (1991), but in general, the Frasnian−Famennian of Shishkat is composed of deep water turbidites, chert, and lime mudstone. Based solely on an assumed Famennian age and using the detailed stratigraphic summaries of Bardashev et al. (2006), it is likely the brachiopods were recovered from the Kulyali Formation which is composed of thin to medium bedded limestone clastic sediments deposited as distal turbidites. Local brachiopod coquinas were noted by the authors though no faunal lists were provided. The overlying Surkhobi Formation, which ranges into the Carboniferous, is also primarily composed of deep-water debris flow deposits (Bardashev et al. 2006).
Reconstructing the paleotectonic setting of the region has been a significant challenge with often contradictory results. Based on the broader tectonic picture, it is likely that the Tien Shan and other regional blocks represent independent island arcs and microcontinental slivers that amalgamated between the Cambrian and Devonian before being sutured onto the leading edge of “Kazakhstania” and ultimately connecting to the Uralian suture (Domeier and Torvisk et al. 2014). It is also likely that tectonism was still active in this region during the Famennian though active arc volcanics are older (Middle Devonian) and younger (Early Carboniferous).
Morocco: Rhynchonellide brachiopods from isolated limestone lenses were first described by Termier (1936, 1938) and Termier and Termier (1948, 1949). The field context and age of the Dzieduszyckia deposits were further described by Hollard et al. (1970) and Hollard and Morin (1973). Additional critical papers on the brachiopods were presented by Ager (1968), Ager et al. (1976), Baliński and Biernat (2003), and Peckmann et al. (2007). A recent review by Jakubowicz et al. (2022) provides an excellent summary of the Moroccan localities.
All of the known Dzieduszyckia collections are from the eastern border of the Western Meseta. Most localities, including the Sidi Amar locality of Peckmann et al. (2007), are from unnamed Devonian limestone blocks found as olistoliths within deep water sandstone and shale of Carboniferous (late Visean) age (Ager et al. 1976). The Sidi Amar blocks are found along a low ridge that strikes parallel to the larger Tabaïnout mountain ridge to the west. The blocks were emplaced into the tectonic basin during the Variscan Orogeny (Walliser et al. 2000; Hoepffner et al. 2005). Therefore, the stratigraphic context is missing though the age is constrained (Peckmann et al. 2007). However, detailed petrologic and geochemical analysis of the limestone blocks clearly established a methane seep origin (Peckmann et al. 2007; Jakubowicz et al. 2022). A locality north of Khenifra along the southeastern slopes of the Gara de M’rirt contains rare, small, isolated blocks embedded within early Carboniferous strata and overlain by nodular griotte limestone (Ager et al. 1976; Becker et al. 2020). This area is referred to as the “Bou-Ounebdou section” in the older literature (e.g., Termier and Termier 1948; Ager et al. 1976). In all cases, brachiopods are generally found as dense coquinas of articulated shells. Coarse cements are rare except for spar within the interiors of articulated shells. It is noteworthy that at least two distinct species are found together at the Sidi Amar locality (Termier and Termier 1948; Peckmann et al. 2007; RSS personal observation; Fig. 3). In addition to these allochthonous deposits, Dzieduszyckia coquinas are found in situ within Famennian deposits west of the Carboniferous basin (Hollard and Morin 1973; El Hassani and Benfrika 2000).

Fig. 3. Field photograph showing two clearly distinguished dimerelloid brachiopod species, Dzieduszyckia tenuicostata (left) and Dzieduszyckia crassicostata (right), together in an allochthononous limestone boulder. Famennian (Upper Devonian), Sidi Amar, Western Meseta, Morocco.
South China: Xian and Jiang (1978) defined four species from Guizhou: Dzieduszyckia oblonga Xian in Xian and Jiang, 1978, Dzieduszyckia elongata Xian in Xian and Jiang, 1978, Dzieduszyckia xiasiensis Jiang in Xian and Jiang, 1978, and Dzieduszyckia acutiplicata Xian in Xian and Jiang, 1978, based on variations in shell shape, depth of the sulcus, and costae density. Further evaluation by Nie et al. (2016) on hundreds of specimens from a broader area concluded that the diversity was related to plasticity and driven by environmental conditions, thus only one species, D. oblonga, was valid.
Deposits of Dzieduszyckia in South China are unique as the fossils are found in outer platform oolite as well as intraplatform basinal limestone (Nie et al. 2016). Originally described from near Guiyang, Guizhou Province from deep water deposits (Xian and Jiang 1978; Xian et al. 1980), the distribution was expanded to include shallower, offshore platform turbidite deposits (Nie et al. 2016; Tong et al. 2021). The fossils comprise dense accumulations of largely articulated shells although one deep water locality at Changtang is noteworthy for transported disarticulated shells (Nie et al. 2016). Tong et al. (2021) performed a detailed sedimentological and stratigraphic analysis and concluded that the shells of the intraplatform basins were transported.
Mexico: Silicified molds of Dzieduszyckia were reported from Devonian massive barite deposits of Mazatán, east of Hermosillo, Sonora, Mexico by Noll et al. (1984). The fossils are found in two localities from both barite and reddish siltstone pockets within the barite of the Los Pozos Formation. Overall, the stratigraphy of the Guayacan Group is dominated by argillite and siliceous shales (Noll 1981). Boucot et al. (2008) reported conodonts of the late Marginifera Zone for the deposit at Rancho Los Chinos. The geology of the area is structurally quite complex (Noll et al. 1984). The authors note that the brachiopods are found in dense clusters within monospecific coquinas and that individuals are mostly articulated. The detailed paleoenvironment is not well known but a shelf-margin or upper slope deposit was proposed. Based on the co-occurrence of Nereites-facies trace fossils, thin-bedded argillite, siltite, chert, barite, and rare limestone, Boucot et al. (2008) narrowed the environment to bathyal depths in a continental rise environment. Further details restricted the setting to an accretionary tectonic margin beneath a highly productive water column (Canet et al. 2014). Canet et al. (2014) firmly established a methane seep ecosystem based primarily on significantly 13C depleted carbonates, 34S enriched barites showing sulfate reduction coupled to the anaerobic oxidation of methane, and gas-rich fluid inclusions within the barite.
USA: Occurrences of Dzieduszyckia in Laurussia were first confirmed by Cloud and Boucot (1971) from massive barite deposits of the Slaven Chert in Nevada. The Nevada fossils were originally misidentified as Halorella though the U.S. Geological Survey stratigraphers felt the barite beds should be older than Triassic. Following the clarification of Biernat (1967), Cloud and Boucot (1971) correctly recognized the genus and the Late Devonian age. Though no fossils have been formally presented, Boucot et al. (2008) noted that the Slaven Chert Dzieduszyckia were associated with conodonts of the lower and possibly upper Marginifera Zone in the northern Shoshone Range.
The Nevada brachiopods come from several open-pit barite mines in the northern Shoshone Range. Cloud and Boucot (1971) relied on U.S. Geological Survey collections from the Greystone, Hilltop and Mound Spring Mine (both of the latter are now part of the current Mountain Springs Mine). Additional fossils collected and described in the present paper come from the Clipper Mine. All the fossils are found in massive to coarsely laminated barite beds that are replacive after limestone and chert. The original depositional setting was a deep water outer continental slope or fringing island arc bathyal setting (e.g., Dubé 1988; Madrid et al. 1992). More details are provided below in the Geological setting section. These units may be considered syntectonic with the early stages of the Antler Orogeny in which deep marine units were obducted or overthrusted onto shelfal limestones (Koski and Hein 2003).
It is interesting to note that while there are no other Dzieduszyckia deposits described from the United States, there is a mention of “Leiorhynchoidea-like” brachiopods in spiculitic lime mudstone with interbedded black mudstone and siltstone from a restricted, low-energy marine Famennian settings of Utah (Silberling et al. 1995), several hundred kilometers east of the Dzieduszyckia beds. This environment would have been a foreland basin during the early stages of the Antler Orogeny.
Stratigraphy of occurrences.—All published dates of Dzieduszyckia place the genus as a Famennian fossil and the genus has been used to correlate deposits in the Ural Mountains (e.g., Mizens and Mizens 2017). A graphic compilation (Fig. 4), described below, converts the published zonations to the recent schematic of Spalletta et al. (2017). In the most robust study, Nie et al. (2016) provided strong evidence from conodonts that platform deposits of Dzieduszyckia in South China date to the Upper Triangularis Zone and are found through the Lower and Middle Crepida zones. Deeper water deposits in South China are slightly younger and range up through the Rhomboidea Zone. Thus, the South China fossils are constrained to the lower Famennian.

Fig. 4. Stratigraphic distribution of dimerelloid brachiopod Dzieduszyckia within the Famennian (Upper Devonian) based on conodonts and ammonoids. Biozones based on the divisions of Spalletta et al. (2017), retaining the divisions on lower, middle, upper, and uppermost Famennian. See text for age references.
Polish fossils were reported as potentially as old as Upper Trachytera to Lower Expansa zones (Biernat 1988). Peckmann et al. (2007) recorded the Sidi Amar deposit of Morocco as Upper Triangularis to Upper Marginifera zones, confirming the broader Famennian assignment of Hollard and Morin (1973) based on conodonts and goniatites. In contrast, Menakova (1991) recorded Dzieduszyckia in the Frasnian of Tajikistan but details on the specific fossils and stratigraphy are lacking. Boucot et al. (2008) noted that the Mexican Dzieduszyckia is from the Upper Marginifera Zone and the Nevada deposits had conodonts of the Lower and possibly Upper Marginifera zones but did not provide details.
Rozman (1960) noted that Lipina (1955) recorded Leiorhynchus (Dzieduszyckia) bashkiricus in the Prolobites Ammonoid Zone of the Western Urals and drew the equivalence to the Murzekayev Beds of Kazhakstan. This is also referred to as “Famennian III” in older literature. The Prolobites Zone approximately is coincident with the current Middle Marginifera to Middle Trachytera conodont zones. In the Central Urals, Mizens (2012) presented evidence that Dzieduszyckia prima was clearly assigned to the lower Makarov Horizon (Middle Triangularis–Trepida zones), here interpreted to be restricted to the Platys through Pectinata zones of Spalletta et al. (2017). In the Southeastern Urals, both species of Dzieduszyckia are noted from the upper Murzakaevian regional substage, equivalent to the Trachytera Conodont Zone (Gatovsky et al. 2017). Finally, in the northern or Subpolar Urals, Dzieduszyckia is recorded from the Utahensis through Granulosis zones (Zhuravlev and Sokiran 2020). While the data presented are not refined enough to draw a concrete picture of origination and expansion, it is clear that Dzieduszyckia is restrained to the Famennian, between the major extinction intervals (or biotic crises) of the Lower and Upper Kellwasser and the Hangenberg events.
Taxonomy: Thus far, there have been twelve species of Dzieduszyckia recognized from the Upper Devonian (Tables 1 and 2). However, there is still discussion on the validity of differentiating species versus intraspecific morphological plasticity as the genus is noted for its broad variability (e.g., Nalivkin 1947; Biernat 1967; Baliński and Biernat 2003; Nie et al. 2016).
Table 1. Species of dimerelloid brachiopod Dzieduszyckia with locations and key references.
|
Species |
Location |
Key references |
|
Dzieduszyckia kielcensis |
Poland |
|
|
Dzieduszyckia bashkirica |
Urals, Kazakhstan, Tajikistan |
|
|
Dzieduszyckia prima |
Middle Urals |
|
|
Dzieduszyckia cordiformis |
Eastern Urals |
|
|
Dzieduszyckia sonora |
Mexico, USA |
|
|
Dzieduszyckia crassicostata |
Morocco |
|
|
Dzieduszyckia intermedia |
Morocco |
|
|
Dzieduszyckia tenuicostata |
Morocco |
|
|
Dzieduszyckia oblanga |
China |
|
|
Dzieduszyckia elongata |
China |
|
|
Dzieduszyckia xiasensis |
China |
|
|
Dzieduszyckia acutiplicata |
China |
Table 2. Key distinguishing features of species of dimerelloid brachiopod Dzieduszyckia. NA, not available.
|
Species |
Shell margin |
Costae |
Additional key feature |
|
Dzieduszyckia kielcensis |
subrounded to pentagonal, subtransverse to transverse elongation |
variable |
NA |
|
Dzieduszyckia bashkiricus |
transversely oval to roundly |
variable but generally few and thick |
thickened and incurved ventral umbo |
|
Dzieduszyckia prima |
flatter profile, deep sulcus in both valves |
coarse and sparse |
smaller than D. bashkirica |
|
Dzieduszyckia cordiformis |
biconvex and heart-shaped |
fine and bifurcating |
NA |
|
Dzieduszyckia sonora |
subtriangular to subovate |
bifurcating and rarely trifurcating |
median size is smaller than European or Moroccan species |
|
Dzieduszyckia crassicostata |
wide, heart-shaped, and globulose |
thick, angular to rounded |
high dorsal septum that thickens posteriorly |
|
Dzieduszyckia intermedia |
asymmetric elongation |
weak radial ornamentation |
high dorsal septum that thickens posteriorly |
|
Dzieduszyckia tenuicostata |
triangular to fusiform shell |
many fine costae; faint and wide irregular plicae |
crura widen anteriorly and twist from horizontal to vertical. |
|
Dzieduszyckia oblanga |
transversely quadrate shell outline |
simple, angular in cross-section |
weak ventral sulcus |
|
Dzieduszyckia elongata |
rounded triangular shell with |
low and round costae, disappear posteriorly |
no sulcus |
|
Dzieduszyckia xianensis |
transversally oval outline |
simple, restricted to anterior of shell |
bisulcate |
|
Dzieduszyckia acutiplicata |
small, subrectangular |
abundant thin costae, many |
weakly bisulcate |
The various species are largely distinguished based on the overall shape of the external shell and the variation of size, shape, and diversity of the costae. Unfortunately, the critical analysis of the interior elements such as the septalium, crural bases, and crura, needs to be expanded upon for many species (Baliński and Biernat 2003).
Regardless, the following sections summarize the differences thus far reported. The type species, D. kielcensis, was distinguished by Biernat (1967) for its large, subrounded to pentagonal biconvex shell, subtransverse to transverse elongation, sulci in opposing valves, and variably sized costae. She noted that the interior of the shell demonstrated hinge plates, short dental plates and small teeth, a distinct septalium, and a dorsal septum that reaches halfway down the length of the shell. Both Nalivkin (1947) and Rozman (1962) described D. bashkirica as having variable shell morphologies but typified by a roundly triangular to transversely oval shell. Later, Menakova (1991) described D. bashkirica as distinctive from D. kielcensis due to its rounded triangular shape and smaller number of costae. Like D. kielcensis, the costae vary in their origination point on the shell relative to the umbo and thickness. The two species may be conspecific though D. bashkirica may be unique due to its thickened and incurved ventral umbo.
In the Middle and Eastern Urals, Mizens (2006, 2008) described two new species of Dzieduszyckia that were distinctive from D. bashkirica. Dzieduszyckia cordiformis from the section at Dzerzhinka is noteworthy for its fine costae and biconvex heart-shaped shell with opposing sulci. In addition, the costae are strongly dichotomous and triangular in cross-section. Mizens (2006) notes the similarity to D. tenuicostata from Morocco (below) but argued that the species was distinct due to the narrower shell and a shorter hinge. In a later paper, Mizens (2008) described D. prima from the Middle Urals. This species is like D. bashkirica but distinguished by its flatter profile, smaller size, deeper sulcus in both valves, and coarser and sparser costae.
The Moroccan species were originally described by Termier (1936) and Termier and Termier (1948) and later carefully re-evaluated by Baliński and Biernat (2003). Like the earlier review of Biernat (1967), they were challenged by defining the differences between D. crassicostata and D. intermedia. Both species possess a high dorsal septum that thickens posteriorly and terminates midway along the shell. The crura are ciliform, similar to D. kielcensis. The crural bases are closely set and near the ventral surface of the hinge plates and fused with the top of the median septum. In contrast, the authors agreed that D. tenuicostata is distinct largely due to its triangular to fusiform shell with many fine costae. Occasional faint and wide irregular plicae were noted on larger D. tenuicostata shells. The crura of D. tenuicostata widen anteriorly and twist from horizontal to vertical. Finally, the median septum is low though still extends half of the shell width.
Xian and Jiang (1978) distinguished four species of Dzieduszyckia from South China. Dzieduszyckia oblonga was noted for the transversely quadrate shell outline with simple costae and a weak ventral sulcus. The costae are more pronounced anteriorly than posteriorly. Dzieduszyckia elongata has a relatively larger and rounded triangular shell with distinct geniculation in the lateral and anterior commissures, and no fold or sulcus. Dzieduszyckia xiasiensis was distinguished by the transversally oval outline and biconvex shell with distinct sulcus in both valves. Finally, Dzieduszyckia acutiplicata was distinguished for having a smaller shell, though biconvex and weakly bisullcate and more and finer costae. However, a review by Nie et al. (2016) that included more specimens and additional localities concluded that the forms were all variations of a single species (D. oblonga; Yuanlin Sun, personal communication, 2022), though there are some patterns defined by facies (platform or intraplatform basin) and locality. The most recent study of Tong et al. (2021) included more specimens and additional localities but there was no further discussion of distinguishing separate species.
Dzieduszyckia sonora was described by Noll et al. (1984) from silicified molds preserved in barite in the Hermosillo region of Mexico. Although more poorly preserved and lacking some internal details, they felt a new species was justified due to the commonly bifurcating and rarely trifurcating costae. The biconvex shell varies greatly from subtriangular to subovate and possesses a gently incurved beak. The internal molds demonstrate short, widespread dental plates, a small septalium, and a prominent septum extending halfway along the shell. The authors also drew attention to the fact that the median size of D. sonora is smaller than European or Moroccan specimens. No details were provided on the crura or crural bases.
It should be noted that historically, many of these brachiopods were originally referred to the genus Leiorhynchus (or Nudirostra), a small costate rhynchonellide used as a biostratigraphic marker within the Devonian. In Xian and Jiang (1978), several fossils referred to as new species of Leiorhynchus may actually be Dzieduszyckia (Yuanlin Sun, personal communication 2022). Additional comments on the connection with Leiorhynchus are discussed below. Other incorrect generic assignments include Eoperegrinella from Morocco and China as well as the Triassic Halorella in Nevada (USA) and Morocco.
Geological setting
The brachiopod deposits described in this paper are found in rare, isolated lenses within the Upper Devonian Slaven Chert in the Shoshone and Toquima ranges, northcentral Nevada (USA). The exact stratigraphic position is difficult to establish as the Slaven Chert is highly faulted and folded with poor surface exposures. Nevertheless, some general comments on the stratigraphy and geologic setting may be deduced.
Originally described by Gilluly and Gates (1965) in Slaven Canyon near Mount Lewis in the Shoshone Range, the Slaven Chert is found throughout northcentral Nevada where it is a unit within the lower plates of one or several major low angle faults (Speed and Sleep 1982; Miller et al. 1984; Burchfiel and Royden 1991; Holm-Denoma et al. 2011, 2017). The older literature refers to this tectonic package as the “Roberts Mountains Allochthon” or “Western Facies Assemblage” (Ketner 2013) (Fig. 5). This package of units ranges from the Cambrian to uppermost Devonian and represents primarily low-energy, deep water deposition. Proximal and distal turbidites, debris-flow deposits, and hemipelagic deposits indicate submarine slope-fan-basin floor environments (Poole 1974). Although the lower Cambrian–Silurian formations contain terrigenous detrital grains, the younger Slaven Chert is relatively clean suggesting a distal or isolated depocenter.
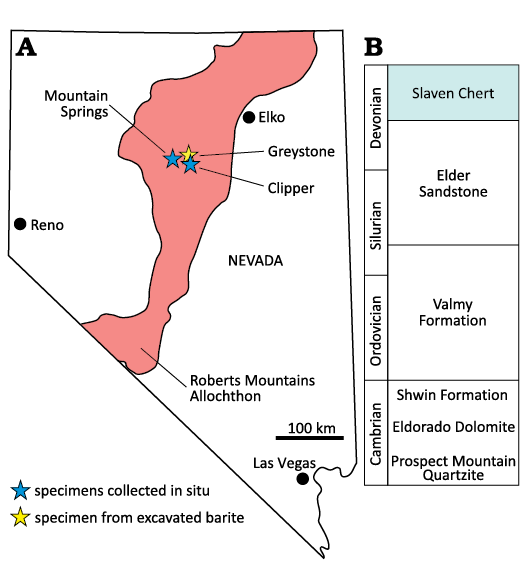
Fig. 5. A. The distribution of the Roberts Mountains Allochthon with key mines cited in this study. B. Stratigraphy of the Roberts Mountains Allochthon. Note exact position of Dzieduszyckia within the Slaven Chert is unknown due to intense faulting within the allochthon. Base map modified from Holm-Denoma et al. (2017). Stratigraphic chart based on Ketner (2013).
The majority of the Slaven Chert is composed of dark radiolarian and spicular chert with lesser amounts of barite, phosphate, siltstone and sandstone, rare limestone and very rare greenstone (Gilluly and Gates 1965; Koski and Hein 2003; Donkervoort et al. 2022). Beds are typically less than 10 centimeters thick though rare limestone layers in the Cortez Mountains reach over 5 meters in thickness (Gilluly and Masursky 1965) (Fig. 6). The total thickness is unknown but likely exceeds several hundreds of meters. Fossils are rare but include ostracods, tentaculitids, corals, bryozoans, and crinoid columnals (Gilluly and Masursky 1965).
There are multiple models on the depositional setting of the Slaven Chert though they all agree on a deep marine setting. Earlier models purport the Slaven Chert and the rest of the western facies assemblage to have been deposited along the slope of an island arc that collided with North America (Speed and Sleep 1982). Burchfiel and Royden (1991) presented evidence that the deformation and sedimentation more likely supported deposition in a back-arc extensional basin. An alternative model suggests that deposition happened in rift basins developed on Laurentian continental crust (Madrid et al. 1992; Finney et al. 1993).
The age of the Slaven Chert is taken as Famennian based on radiolarians and conodonts (Holdsworth 1980; Holdsworth and Jones 1980; Dubé 1988). However, Boundy-Sanders et al. (1999) reported a suite of conodonts, radiolarians, and sponge spicules from the type section at Slaven Canyon that pushed the maximum age to the Frasnian (late Rhenana stratigraphic Zone) (Ziegler and Sandberg 1990). It is of note that the authors also draw the connection to the Sonora district near Hermosillo, Mexico. As noted above, Boucot et al. (2008) specified that the Nevada Dzieduszyckia collections were associated with conodonts of the early and possibly late Marginifera zones.

Fig. 6. A typical Slaven Chert exposed in open pit mines from Famennian, Nevada. A. Interbedded dark chert and light grey barite. Beds above fault approximately 10 cm thick. B. Exposure of in situ brachiopod molds in floor of Mountain Springs Mine (Pit A).
Material and methods
Specimens for the present study were collected during surveys of active and closed open-pit barite mining operations in the northern Shoshone Range, Nevada between 2016 and 2020. Specifically, fossils were collected in situ from a bench floor of the Mountain Springs Mine (Pit A) and pit wall of the closed Clipper Mine. Additional specimens were collected from barite stockpiles at the Mountain Springs and Clipper Mines as well as the Greystone Mine (see localities in Fig. 5 and Table 3). In total, over fifty rock samples were collected and many of these provided multiple brachiopod specimens. All specimens are preserved as external or internal molds within barite. A few specimens from the Clipper Mine stockpile are preserved as calcite within coarsely recrystallized calcite. In total, over 100 individual fossils were recovered from the barite or barite-limestone samples.
Table 3. Nevada localities of dimerelloid brachiopod Dzieduszyckia noted in this paper. Note that Mountain Springs and Greystone are privately owned mines and Clipper is now on federally-managed (Bureau of Land Management) lands. Coordinates are in the WGS84 datum.
|
Mine |
Latitude, longitude |
Description |
Figured specimens |
|
Mountain Springs |
40.32045, -117.04322 |
in situ brachiopods collected from barite beds in Pit A |
none |
|
Mountain Springs |
40.31555, -117.04208 |
loose specimens collected from Pit C |
USMN PAL 794578 |
|
Clipper |
40.25089, -116.83484 |
in situ brachiopods collected from faulted massive barite in mine wall |
USMN PAL 794576, 794577, 794582–794586 |
|
Clipper |
40.24946, -116.83063 |
loose specimens recovered from high-grade barite stockpile |
USMN PAL 794579–794581 |
|
Greystone |
40.27160, -116.86859 |
singular loose specimen collected from excavated high-grade barite |
none |
Petrographic analysis was carried out on standard thin sections of 30 µm thickness on an Olympus BX-51 petrographic microscope at California State University, Chico. Polished samples of primarily limestone were also observed for cathodoluminescence with a Reliotron CL scope mounted on a Nikon Optiphot microscope with a 10× objective. The Reliotron was run with the vacuum at ~42 mTorr and an operating voltage of 7.8 kV and power at ~0.032 DCmA.
Samples were prepared for taxonomy with the use of air-hammers, focusing mainly on external molds. For photography and measurements, casts using modeling clay were utilized for some samples. Because of the incomplete nature of the preservation, standard taxonomic techniques could not be employed. Instead, brachiopod internal and external molds were measured for minimum width and numbers of costae. If the sulcus was present, the minimum “half-width” of the shell was also recorded (Fig. 7). Beaks were very rarely preserved so the length of the shells recorded are a minimum and are not statistically significant. In total, 93 individual shells were measured. Samples with less than six costae preserved were not included but were used for detailed description and/or thin-section petrography.
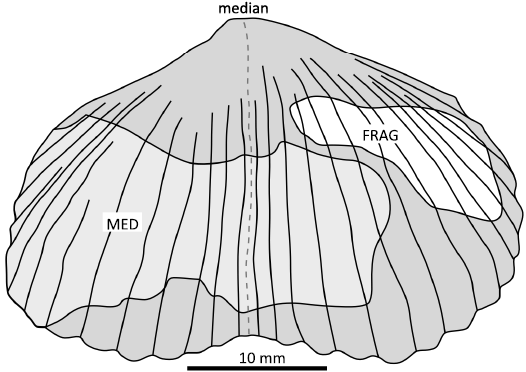
Fig. 7. Measurements of shell features. FRAG, measured fossil is a fragment of a valve. MED, fragment of valve that retains the margin and sulcus and can be used to estimate minimum width.
Stable carbon isotope analyses were performed on rare pockets of calcite within barite from the stockpile at Clipper Mine. Powders were recovered using a handheld rotary drill. Carbonate δ13C values were determined using a Picarro CRDS (G2131-i) coupled to Picarro Liaison interface and a modified AutoMate autosampler in the Will Berelson lab at the Department of Earth Sciences, University of Southern California. The powdered carbonate was weighed and placed in an AutoMate vial and sealed with a rubber septum screw-cap. The AutoMate acidified these samples on-line using 10% phosphoric acid, and the resulting CO2 was carried in a nitrogen stream, through a Nafion desolvating line, to the Picarro Liaison sampling bags. The flow of gas from the AutoMate into the Picarro Liaison’s discrete sample bag was mass flow controlled such that every bag had precisely the same mass of N2+ CO2. Each bag was then introduced to the Picarro CRDS for continuous analysis over an 8 minute interval. Drift over the course of a run (<10 h) was almost never above a few tenths of a permil and resulting Picarro standards (in house CaCO3 standard calibrated against VPDB) typically had a standard deviation of under 0.1‰.
Results
Petrology.—Nearly all the brachiopods are preserved within massive to banded barite beds preserved as crystal mosaics (Fig. 8A). Barite crystals vary in size and are randomly distributed. Most crystals are monocrystalline, others are fragments of radiating fans. Crystals are anhedral with “zigzag” irregular margins. Poikilotopic fabrics are common. In all cases, the crystal margins show no preferred orientation to the voids left by brachiopod valves, suggesting a later stage of diagenetic replacement.
At Clipper Mine, small (millimeter-scale) irregular pockets of calcite are rarely found with the brachiopods (Fig. 8B). The calcite crystals range in size from 28.3 to 256.6 µm with a skewed median of 93.1 µm (n = 218). Most crystals show strong cleavage lines and twinning is common. The crystals are anhedral with jagged margins and enfacial junctions. The smaller crystals could be construed as “microspar” but there is no clear connection to common limestone fabrics such as cements, micrite, or peloids. Very rare brachiopod valves are preserved in the limestone displaying remnants of the fibrous layer though the primary columnar layer is recrystallized into large, twinned calcite crystals (Fig. 8C).

Fig. 8. Slaven brachiopod sediments in thin-section from Famennian, Nevada. A. Void of brachiopod shells surrounded by poikilotopic barite. Note that the barite has no preferred orientation to the shells. B. Intergrowth of coarse twinned calcite and subhedral barite. Use of retardation plate causes barite (ba) to show as bright colors against the calcite (c). C. Poor preservation of layered brachiopod shells within coarse calcite.
One relatively large sample (~90×30 mm) of nearly pure limestone was recovered from the high-grade stockpile at Clipper. The hand specimen is dark grey-black like the barite but the low specific gravity and reaction to dilute hydrochloric acid provided field confirmation as limestone. In thin-section, the calcite crystals are similar to those found as pockets within the barite. While the polished hand specimen shows coarse wavy and parallel banding, there are no clearly defined petrofabrics in the calcite crystals. Under cathodoluminescence, the entire sample glows bright orange with no distinguishing zonation.
Taphonomy.—Except for rare potential preservation as replacive coarse calcite shells within irregularly-sized calcite, nearly all of the brachiopods are preserved as internal and external molds (Fig. 9). No complete shells were recognized; however, it is believed the shells were buried complete and many were articulated and the diagenetic replacement with barite led to the shell destruction. Multiple samples preserve evidence of both shells. Internal details of the hinge area including septalia and the dorsal septa are preserved on internal molds. Unfortunately, no crura or crural bases were observed in any specimen.
Details of the external shell preserve evidence of shell healing and fine details such as concentric growth lines. Based on comparison with samples collected in Morocco, it is believed there was minimal compaction or other deformation of the shells as the overall morphology is similar to Dzieduszyckia crassicostata and D. tenuicostata in preserving rounded biconvex and bisulcate shells.
Shells are commonly found in dense accumulations (Fig. 9B). Most shells are subparallel to each other suggesting preservation along bedding surfaces. Shell size, based on costae width as well as shell dimensions, varies throughout the surface suggesting mixed populations preserved in each sample. Unfortunately, the mode of preservation does not allow for recognition of fragmented shells.

Fig. 9. Preservation (in situ) of brachiopods within massive to bedded barite blocks at Clipper Mine (Famennian, Nevada). A. Details of preservation showing costae and internal features such as the septalium and septum in sample above the scale bar. B. Typical block with dense, subparallel accumulation as seen by molds.
In situ deposits at the Mountain Springs Mine record the brachiopods in beds of pure barite ranging from 50–100 mm thick. The beds are either massive or show subparallel bedding planes at the centimeter scale. As noted above from loose blocks, the fossils are generally subparallel to bedding. The presence of articulated shells suggests burial in place though the preservation is not sufficient for more detailed paleoecological interpretation. Comparison of samples between the three mines did not show any unique, locality-specific patterns.
Carbon isotopes.—Comparing the Slaven Chert limestones to other fossil hydrocarbon seeps is hampered by the pervasive diagenesis. Crucially, one cannot differentiate distinct microfabrics with diverse isotopic signatures common at seeps (e.g., micrite, botryoidal cement, isopachous rim cement). Analysis for biomarkers was not undertaken due to the levels of alteration.
The 43 measured samples from Nevada show a strongly 13C depleted carbonate signature (average δ13C = -30.24‰ VPDB), ranging from -31.1 to -27.3‰ VPDB (Batther 2020 and this study) (Fig. 10 and Table 4). Canet et al. (2014) presented five values from equivalent units in Sonora, Mexico, ranging from -24.3 to -20‰ VPDB (Fig. 10 and Table 4). The overall variation, ~4‰, is not as broad as at many seeps in the geological record. Whether this is a function of diagenetic homogenization or reflects an original signature is still under investigation. However, deep subsurface alteration would more likely increase rather than decrease the ratio of 13C to 12C. This shows that Nevada sites do show a geochemical signal for primary oxidation of methane as one of the inorganic carbon sources.

Fig. 10. δ13C isotopic ratios of limestone from Clipper Mine, Nevada compared to Mexico and Morocco (Famennian). Data from Batther 2020 and this study (Nevada); Canet et al. 2014 (Mexico); Peckmann et al. 2007 (Morocco).
Table 4. Carbon isotope data from limestones associated with dimerelloid brachiopod Dzieduszyckia in Nevada and Mexico. Values are relative to the Vienne Pee Dee Belemnite (VPDB) standard.
|
Location |
δ13C |
Reference |
|
Los Pozos. Sonora, Mexico |
-21.2 |
|
|
-20.0 |
||
|
-23.5 |
||
|
-22.3 |
||
|
-24.3 |
||
|
Clipper. Nevada, USA |
-28.6 |
|
|
-31.1 |
||
|
-30.9 |
||
|
-29.9 |
||
|
-31.7 |
||
|
-30.2 |
||
|
-30.4 |
||
|
-30.4 |
||
|
-29.7 |
||
|
-31.4 |
||
|
-31.2 |
||
|
-27.3 |
||
|
-30.8 |
||
|
-29.7 |
this study |
|
|
-30.2 |
||
|
-30.5 |
||
|
-30.5 |
||
|
-30.1 |
||
|
-30.2 |
||
|
-29.7 |
||
|
-29.0 |
||
|
-30.5 |
||
|
-30.0 |
||
|
-29.8 |
||
|
-30.3 |
||
|
-30.5 |
||
|
-30.3 |
||
|
-29.7 |
||
|
-29.7 |
||
|
-30.3 |
||
|
-30.5 |
||
|
-30.1 |
||
|
-30.8 |
||
|
-30.3 |
||
|
-30.6 |
||
|
-30.5 |
||
|
-30.3 |
||
|
-30.1 |
||
|
-30.6 |
||
|
-30.8 |
||
|
-30.5 |
||
|
-30.3 |
||
|
-30.3 |
Systematic palaeontology
Phylum Brachiopoda Duméril, 1805
Subphylum Rhynchonelliformea Williams et al., 1996
Class Rhynchonellata Williams et al., 1996
Order Rhynchonellida Kuhn, 1949
Superfamily Dimerelloidea Buckman, 1918
Family Halorellidae Ager, 1965
Subfamily Dzieduszyckiinae Savage, 1996
Genus Dzieduszyckia Siemiradzki, 1909
Type species: Terebratula (?) kielcensis Roemer, 1866; Upper Devonian, Kadzielnia, Poland.
Dzieduszyckia spp.
Figs. 11, 12.
1971 Dzieduszyckia sp.; Cloud and Boucot 1971: 175–180, p1. 1: 6–17.
Material.—Over one hundred specimens (including ten figured specimens USNM PAL 794576–794586 and materials of Cloud and Boucot 1971, Table 5: 160184–160193, 337301–337325) preserved as internal and external molds in diagenetic barite and rare crystalline limestone from the Clipper, Mountain Springs, and Greystone Mines, northern Toiyabe Range, Nevada, USA (see Table 3 for locality details). Note that the following discussion is based on new collections and the author did not review the collections at the USNM due to restrictions emplaced during the coronavirus epidemic (Table 5).
Table 5. Specimens of Dzieduszyckia sp. in the collections of the USNM.
|
USNM |
Scientific name (as filed) |
Reference |
Date collected |
Province/State |
Site/Station Number |
|
337301 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337302 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337303 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337304 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337305 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337306 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337307 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337308 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337309 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337310 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337311 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337312 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337313 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337314 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337315 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337316 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337317 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337318 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337319 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337320 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10453 |
|
|
337321 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10452 |
|
|
337322 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10452 |
|
|
337323 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10452 |
|
|
337324 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10452 |
|
|
337325 |
Dzieduszyckia sonora Noll et al. |
1980 |
Sonora, Mexico |
10452 |
|
|
160184 |
Dzieduszyckia sp. of Cloud & Boucot |
1957 |
Nevada, USA |
USGS GREEN LOC M134 |
|
|
160185 |
Dzieduszyckia sp. of Cloud & Boucot |
1957 |
Nevada, USA |
USGS GREEN LOC M134 |
|
|
160186 |
Dzieduszyckia sp. of Cloud & Boucot |
1957 |
Nevada, USA |
USGS GREEN LOC M134 |
|
|
160187 |
Dzieduszyckia sp. of Cloud & Boucot |
unknown |
Nevada, USA |
unrecorded |
|
|
160188 |
Dzieduszyckia sp. of Cloud & Boucot |
1957 |
Nevada, USA |
USGS GREEN LOC M134 |
|
|
160189 |
Dzieduszyckia sp. of Cloud & Boucot |
1957 |
Nevada, USA |
USGS GREEN LOC M134 |
|
|
160190 |
Dzieduszyckia sp. of Cloud & Boucot |
1950 |
Nevada, USA |
USGS GREEN LOC 25453 |
|
|
160191 |
Dzieduszyckia sp. of Cloud & Boucot |
1957 |
Nevada, USA |
USGS GREEN LOC M134 |
|
|
160192 |
Dzieduszyckia sp. of Cloud & Boucot |
1957 |
Nevada, USA |
USGS GREEN LOC M134 |
|
|
160193 |
Dzieduszyckia sp. of Cloud & Boucot |
1957 |
Nevada, USA |
USGS GREEN LOC M134 |
Description.—No complete shells were observed in this study but many external and internal molds preserved enough details for general comment on external form and internal details. Overall shell outlines include both rounded triangular and transverse biconvex shells though most fossils are preserved as incomplete fragments that do not define the margin (Fig. 11). Where a midline was preserved, shells have a median sulcus on both valves. As noted, no complete shells were recovered but the largest individual fragment is 73 mm wide. By using the presence of the sulcus, the largest half-width measured is 43 mm, yielding a minimum total width of 86 mm. Along the lateral and anterior margins, many shells are rounded to rectilinear. The ventral umbo is distinct and strongly recurved. In rare examples, fine growth rings and plicae were observed.

Fig. 11. External molds of dimerelloid brachiopod Dzieduszyckia spp., Famennian, Nevada, USA. A. USNM PAL 794576, block showing dense concentration of shells from in situ deposit, Mountain Springs Mine. B. USNM PAL 794577, cast showing typical triangular outline and deep sulcus. C. USNM PAL 794578, shell fragment showing relatively large costae and sulcus. D. USNM PAL 794579, specimen showing thinner costae and asymmetry about the midline. Note the rare bifurcation to left of midline. E. USMN PAL 794580, fragment of a more rounded shell with rounded costae. F. USNM PAL 794581, portion of shell showing radially divergent costae. G. USNM PAL 794582, external mold emphasizing strong biconvexity of shell.
Costae are well-preserved and show variability from sharp rectangular to rounded and triangular cross-sections. Many shells show complete costation; in others the costae on the external surface start approximately one third of the distance from the umbo. Bifurcation of costae is very rare (only two samples out of over 100 noted) and no trifurcation was observed.
While no crura were observed in this study, a number of critical internal features are preserved on internal molds (Fig. 12). A dorsal median septum is pronounced and preserved in multiple specimens. The septum is relatively thick in smaller shells but does not thicken with the increase in shell size. No complete shell was observed but based on the dimensions it is likely the septum extends to approximately one half of the shell length. Small dental sockets can be observed. The septalium is distinct and thicker than the septum. However, internal skeletal growth sequences are not preserved. Additionally, several fortuitous samples preserve the impressions of dental plates.

Fig. 12. Internal features of dimerelloid brachiopod Dzieduszyckia sp., Famennian, Nevada, USA. A. USNM PAL 794583, dorsal valve showing pronounced septum. B. USNM PAL 794584, internal mold of small shell showing deep septum and biconvex shell. C. USNM PAL 794585, view of hinge displaying septalium in upper, dorsal shell and vertical dental plates in lower, ventral shell. D. USNM PAL 794586, internal mold preserving relatively tall septalium. Scale bars 10 mm.
Discussion.—Noll et al. (1984) provided detailed descriptions of the external morphology of Dzieduszyckia from silicified fossils in barite from Sonora but did not undertake serial sectioning to show the crura shapes and bases. However, the authors argued for the erection of a new species, D. sonora, based primarily on bifurcation and trifurcation of costae in the mature shells and secondarily on the generally smaller size. They also stated that the Nevada fossils were the same species. In their table of twenty specimens, 18 show bifurcation and/or trifurcation (Noll et al. 1984: 1419, table 1). Only two of the over 100 brachiopods collected for the present study and only one of the specimens figured by Cloud and Boucot (1971: pl. 1: 15) shows any bifurcation. In each of those cases, only one or two costae were bifurcated. Noll et al. (1984: fig. 6) also relied on the size variation to connect the Mexican and Nevada specimens. They show a figure with the measured ratios of width to length of type specimens as well as means for D. sonora, D. kielcensis, D. crassicostata, and D. bashkirica. The Mexican species has a smaller width and length average (~30 mm for each dimension). Noll et al. (1984) did not provide the measurements of the Nevada specimens but lumped them with the Mexican samples. Using the minimum widths of Nevada shell fragments, the average minimum width is 43 mm, more in line with D. kielcensis and D. crassicostata. The data provided herein support distinguishing a separate species from D. sonora. Specifically, the lack of common bifurcation (and no trifurcation) of the costae, a relatively thin dorsal septum, and larger size are contradictory to the Mexican species diagnosis.
Ninety-three specimens were measured for number of costae relative to the width of the fossil fragment. However, the line of cross-section was random and not along the commissure due to the limited material. The results are plotted as a histogram with bins set to 0.10 costae/mm width (Fig. 13A).
The histogram may be interpreted in two ways. In the first, the observations represent variance within a single species, in which costae frequency width skews toward lower values. A scatter plot (Fig. 13B) shows the observed fragment width and observed costae have a significant (p = 7.751×10-5), if middling, positive correlation (Pearson’s rho = 0.41).
Alternatively, the data may represent two distinct species of brachiopods, with low (ca. 0.5 costae/mm) and high (ca. 1.0 costae/mm) median costae frequencies, respectively. In this interpretation, the lack of observed specimens with 0.8–0.9 costae/mm (Fig. 13A) coincides with each species distribution’s upper and lower tails, respectively. When the data are cut at this threshold, the tallied costae correspond more closely to fragment size. As shown in Fig. 13C, specimens with higher costa frequency (costa/width >0.8) have a strong, significant correlation between costae tally and fragment size (Pearson’s rho = 0.91; p = 4.69×10-10), while specimens with lower costa frequency (costae/width <0.8; red{?}) retain a strong significant association (Pearson’s rho = 0.69; p = 1.19×10-10). Based solely on these values, it may be possible to draw out two separate species. The low numbers of recovered specimens as well as the use of fragments of shells must be considered. Coupled with the lack of details of the crura, there is a hesitation to erect two distinct species. This relationship of two species of Dzieduszyckia inhabiting the same environment is similar to Morocco, where two dominant species—D. crassicostata and D. tenuicostata—are easily defined by density of costae relative to the shell (Termier and Termier 1948). Baliński and Biernat (2003) also noted that the crura of D. tenuicostata widen anteriorly and twist from horizontal to vertical, a comparison thus far not tested in Nevada. A similar situation is found in the Southern Urals where Mizens (2006) defined D. cordiformis as a distinct species from D. bashkirica and D. prima in part by its fine costae. Finally, a similar conclusion was drawn by Xian and Jiang (1978) in China, erecting D. acutiplicata based on its fine costae.
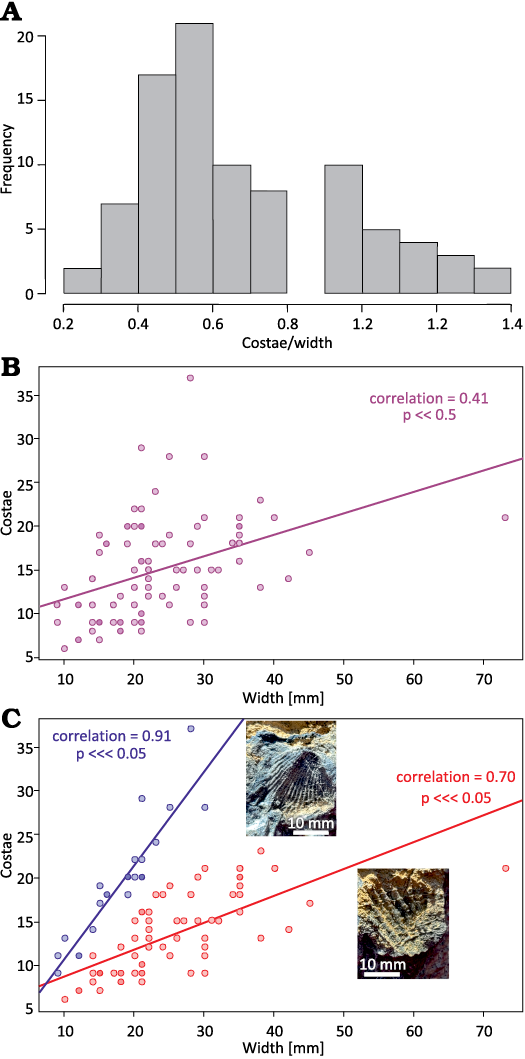
Fig. 13. Number of costae per width in centimeters of fragments in Dzieduszyckia sp. from Nevada. A. Histogram of data from all measured specimens. B. Best fit regression line assuming variance within a single species, in which costae frequency width skews toward lower values. C. Separation into two distinct species of brachiopods, with low (ca. 0.5 costa/mm) and high (ca. 1.0 costa/mm) median costae frequencies with best fit regression lines. Photos show one field sample with a representative of each group.
Discussion
Understanding the role of Dzieduszyckia, the oldest dimerelloid, in hydrocarbon seeps is the most critical question about this large Devonian genus. The carbon isotopic data from Nevada very strongly supports the connection to an original methane seep system even though the diagenetic alteration to barite destroys primary petrofabrics. Similar deposits in the Los Pozos Formation, Mexico, also show a geochemical record of hydrocarbon seepage. Indeed, Torres et al. (2003) expanded this connection to say that large scale bedded barite deposits are related to methane seepage; however, Batther (2020) provides convincing evidence that the methane seepage is coincidental to the barite deposits.
Expanding globally, the only other Dzieduszyckia deposit specifically analyzed for a connection to hydrocarbon seeps is the Sidi Amar locality in Morocco. Baliński and Biernat (2003) argued against a seep setting based on seawater stable carbon isotope ratios in shell material but Peckmann et al. (2007) performed a study using multiple fabrics and showed carbon isotope depletion relative to seawater. The values of Peckmann et al. (2007) were confirmed by the author using material also collected at Sidi Amar. Furthermore, Jakubowicz et al. (2022) argued that these values showed significant depletion compared to contemporaneously high Famennian sea water values though this is contradictory to the Nevada values not available to those authors.
In contrast, the diverse localities in South China have been argued not to be seep settings. This is largely based on the paleogeographic setting (outer carbonate platform as well as intrabasinal deep water deposits) and stratigraphy. While no geochemical data have thus far been published, preliminary data on 13 different petrofabrics provided to the author strongly argue for normal ocean carbonate δ13C values without hydrocarbon input (Yuanlin Sun, personal communication 2022). However, in the same communication it is noted that hydrocarbons are found as pockets in the limestone.
Broadening the discussion to a global scale, nearly all the other occurrences of Dzieduszyckia are from deeper water, organic-rich and likely low oxygen settings that are also connected with either active tectonism or slightly pre-dating major orogenic episodes. It should be re-emphasized that the distribution is global and across diverse ocean basins. Deep water slope and basin settings along active tectonic margins are among the most common regions for hydrocarbon seeps throughout the geological record (e.g., Sandy et al. 2012; Peckmann et al. 2013; Hryniewicz 2022).
Additionally, it is important to note that the dimerelloids got their start in the narrow window of two major extinction intervals: the Frasnian–Famennian Kellwasser Events and the end-Famennian Hangenberg Biotic Crisis. The analysis of this interval is beyond the scope of this paper but in general, the modern consensus is that these biotic crises were related to high productivity and concomitant oxygen depletion (Reershemius and Planavsky 2021) as well as eustatic transgressions that constrained vicariance-driven evolutionary diversity throughout the Famennian (Stigall 2012). It is possible that the Famennian biotic crises removed mollusks from seeps, allowing dimerelloids to access these environments (Baliński et al. 2022). Baliński et al. (2022) proposed that the dimerelloids evolved out of a lineage of brachiopods that were already adapted to dysoxic and low nutrient deep-water settings (e.g., Boyer and Droser 2007) and thus were generalists in depauperate conditions. I agree and would add that this environmental plasticity enabled Dzieduszyckia to be primed for the transgressions of the Famennian. There is still work to be done to develop this hypothesis but it is interesting how the leiorhynchids are found in similar deep water settings in the Frasnian and Famennian. In fact, the generic name Leioryhnchus has been used for species of Dzieduszyckia as well as the Carboniferous dimerelloid, Ibergirhynchia (Gischler et al. 2003). Potentially, both dimerelloids and leiorhynchids are related through the same ancestral rhynchonellide. Regardless of the origin, in this hypothesis, some localized populations found themselves coincidentally at sites of hydrocarbon seepage and adapted to this environment. This is why Dzieduszyckia, more so than other dimerelloids, is found at both seep and non-seep settings. Eventually, the populations that successfully inhabited seeps gave rise to the dimerelloid descendants such as Ibergirhynchia, Halorella, and Anarhynchia. Other subpopulations not found at seeps or vents became evolutionary dead-ends; however, see important discussion of non-seep dimerelloids in Kiel et al. (2021).
This conjectural model explains a few of the intriguing features about Dzieduszyckia. First, of all the dimerelloids, the genus shows the most diversity including the strong likelihood of multiple species cohabitating. This diversity is a representation of the morphological plasticity afforded during a protracted time of intense environmental perturbation. Post-Devonian seeps are monospecific with regards to brachiopods, perhaps alluding to a time when only derived seep-obligate forms were successful. Secondly, nearly all dimerelloids are only found in seep or vent settings, with the exception of some extraordinary Halorella and Sulcirostra deposits (Kiel et al. 2021). As the first dimerelloid, Dzieduszyckia evolved during a time of experimentation thus is found at both seep (Nevada, Mexico, Morocco) and non-seep settings (China and possibly Poland).
Conclusions
Dzieduszyckia brachiopods preserved in diagenetic barite and rare coarse limestone record a methane seep system from the Famennian (Late Devonian) time period. Preserved as internal and external molds, there is not enough internal structure preserved to allow for species diagnosis; however, the fossils are not equivalent to the diagnosis of D. sonora (Noll et al. 1984). Based on nearly 100 partial shell impressions, it may be possible to distinguish two separate species based on the coarseness and density of the costae, similar to situations in Morocco (Baliński and Biernat 2003; Peckman et al. 2007), the Urals (Mizens 2006), and South China (Xian and Jiang 1978). However, the recognition of one or two distinct species is not currently supported by the available material and will remain for future studies. The origination and global distribution of Dzieduszyckia in the dysoxic and nutrient-poor oceans of the Famennian, suggests that the Dimerelloidea may have evolved from the same ancestral stock as the leiorhynchids, another rhynchonellide from similar settings. Dzieduszyckia-bearing deposits are also connected in time to the interval between the Kellwasser and Hangenberg biotic crises. While Dzieduszyckia may have lived in both seep and non-seep settings, younger dimerelloids are obligate to seeps or hydrothermal vents with rare exceptions. Dzieduszyckia represents an initial experimental phase in the dimerelloid lineage with subsequent genera volumetrically dominating hydrocarbon seeps for 230 million years.
Acknowledgements
The author is grateful to the many years of discussions of seeps with Andrzej Kaim (ZPAL, Institute of Paleobiology Polish Academy of Science, Warsaw, Poland), Jörn Peckmann (Universität Hamburg, Germany), Kathy Campbell (University of Auckland, New Zealand), and Michał Jakubowicz (Adam Mickiewicz University, Poznań, Poland). M-I SWACO, and in particular, Ryan Dockstader, provided access to the private barite mines and encouraged the research. Two graduate students, Harpreet Batther and Brady Derick (California State University, Chico, USA), conducted detailed studies of the barite deposits. Geochemical analysis was graciously provided by Will Berelson (University of Southern California, Los Angeles, USA) and Alex Sessions (California Institute of Technology, Pasadena, USA). Kathleen Ritterbush (University of Utah, Salt Lake City, USA) conducted the statistical analysis presented in Fig. 13. Fieldwork in Morocco was facilitated by Michał Jakubowicz and funded by the California State University. Most critical to this manuscript were the papers provided by Andrzej Kaim, Andrzej Baliński (both ZPAL), Anita Mizens (Institute of Geology and Geochemistry, Ural Branch, Russian Federation), and Yuanlin Sun (Peking University, Bejing, China), the latter translated from the original Chinese. All other translations were by the author who accepts responsibility for inaccuracies. The present manuscript benefited immensely from very thorough and detailed reviews by Miguel Manceñido (La Plata Natural Sciences Museum, Argentina), Andrzej Baliński (ZPAL), and two anonymous reviewers, who significantly improved the quality of this paper.
References
Ager, D.V. 1965. Mesozoic and Cenozoic Rhynchonellacea. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology, H, Brachiopoda, 597–632. Geological Society of America, University of Kansas Press, Lawrence.
Ager, D.V. 1968. The supposedly ubiquitous Tethyan brachiopod Halorella and its relations. Journal of the Palaeontological Society of India 5–9: 54–70.
Ager, D.V, Cossey, S.P.J., Mullin, P.R., and Walley, C.D. 1976. Brachiopod ecology in Mid-Palaeozoic sediments near Khenifra, Morocco. Palaeogeography, Palaeoclimatology, Palaeoecology 20: 171–185. Crossref
Amano, K., Kiel, S., Hryniewicz, K., and Jenkins, R.G. 2022. Bivalvia in hydrocarbon seeps. In: A. Kaim, J.K. Cochran, and N.H. Landman (eds.), Ancient Hydrocarbon Seeps. Topics in Geobiology 53: 267–322. Crossref
Angiolini, L., Crippa, G., Azmy, K., Capitani, G., Confalonieri, G., Della Porta, G., Griesshaber, E., Harper, D.A.T., Leng, M.J., Nolan, L., Orlandi, M., Posenato, R., Schmahl, W.W., Banks, V.J., and Stephenson, M.H. 2019. The giants of the phylum Brachiopoda: a matter of diet? Palaeontology 62: 889–917. Crossref
Baliński, A. and Biernat, G. 2003. New observations on rhynchonelloid brachiopod Dzieduszyckia from the Famennian of Morocco. Acta Palaeontologica Polonica 48: 463–474.
Baliński, A., Bitner, M.A., and Jakubowicz, M. 2022. Brachiopods at hydrocarbon seeps. In: A. Kaim, J.K. Cochran, and N.H. Landman (eds.), Ancient Hydrocarbon Seeps. Topics in Geobiology 53: 223–252. Crossref
Baranov, V.V. and Sartenaer, P. 1996, Momarhynchus, new Lower Famennian rhynchonellid brachiopod genus from Yakutia. Bulletin de L’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre 66: 37–42.
Bardashev, I.A., Bardasheva, N.P., Weddige, K., and Ziegler, W. 2006. Stratigraphy of the Shiskat Paleozoic reference section (Southern Tien-Shan, Central Tajikistan). Senckenbergiana lethaea 86: 289–319. Crossref
Batther, H. 2020, The biogeochemistry of Devonian barite deposits in the Roberts Mountains Allochthon, Nevada. M.Sc. Thesis, California State University, Chico. 138 p.
Becker, R.T., Aboussalam, Z.S., El Hassani, A., Hartenfels, S., and Hüneke, H. 2020. Devonian and basal Carboniferous of the allochthonous nappes at Mrirt (eastern part of Western Meseta)—review and new data. Hassan II Academy of Science and Technology, Frontiers in Science and Engineering 10: 87–194.
Biernat, G. 1967. New data on the genus Dzieduszyckia Siemiradzki, 1909 (Brachiopoda). Acta Paleontologica Polonica 12: 133–156.
Biernat, G. 1988. Famennian brachiopods of the Holy Cross Mountains, Poland. In: N.J. McMillan, A.F. Embry, and D.J. Glass (eds.), Devonian of the World. Proceedings of the Second International Symposium on the Devonian System. Canadian Society of Petroleum Geologists, Memoir 14: 327–355.
Boucot, A.J., Poole, F.G., Amaya-Martinez, R., Harris, A.G., Sandberg, C.A., and Page, W.R. 2008. Devonian brachiopods of southwesternmost Laurentia; biogeographic affinities and tectonic significance. In: R.B. Blodgett and G.D. Stanley, Jr. (eds.), The Terrane Puzzle, New Perspectives on Paleontology and Stratigraphy from the North American Cordillera. Geological Society of America Special Paper 442: 77–97. Crossref
Boundy-Sanders, S.Q., Sandberg, C.A., Murchey, B.L., and Harris, A.G. 1999. A late Frasnian (Late Devonian) radiolarian, sponge spicule, and conodont fauna from the Slaven Chert, northern Shoshone Range, Roberts Mountains Allochthon, Nevada. Micropaleontology 45: 62–68. Crossref
Boyer, D.L. and Droser, M.L. 2007. Devonian monospecific assemblages: new insights into the ecology of reduced-oxygen depositional settings. Lethaia 40: 321–333. Crossref
Bronn, H.G. 1832. Ergebnisse meiner naturhistorisch-ökonomischen Reisen. Vol. 2. Skizzen und Ausarbeitungen über Italien, nach einem zweyten Besuche im Jahre 1827. xviii + 686 pp. Karl Groos, Heidelburg.
Buch, L., von 1834. Über Terebrateln, mit einen Versuch, sie zu classificiren und zu beschreiben. 124 pp. Königliche Akademie der Wissenschaften, Berlin.
Buckman, S.S. 1918. The Brachiopoda of the Namyau Beds, Northern Shan States, Burma. Memoirs of the Geological Survey of India, Palaeontologia Indica (new series) 3 (2): 1–299.
Burchfiel, B. and Royden, L. 1991. Antler orogeny: a Mediterranean-type orogeny. Geology 19: 66–69. Crossref
Campbell, K.A. and Bottjer, D.J. 1995a. Brachiopods and chemosymbiotic bivalves in Phanerozoic hydrothermal vent and cold-seep paleoenvironments. Geology 23: 321–324. Crossref
Campbell, K.A. and Bottjer, D.J. 1995b. Peregrinella; an Early Cretaceous cold-seep-restricted brachiopod. Paleobiology 21: 461–478. Crossref
Canet, C., Anadón, P., González-Partida, E., Alfonso, P., Rajabi, A., Pérez-Segura, E., and Alba-Aldave, L.A. 2014. Paleozoic bedded barite deposits from Sonora (NW Mexico): Evidence for a hydrocarbon seep environment of formation. Ore Geology Reviews 56: 292–300. Crossref
Case D.H., Pasulka, A.L., Marlow, J.J., Grupe, B.M., Levin, L.A., and Orphan, V.J. 2015. Methane seep carbonates host distinct, diverse, and dynamic microbial assemblages. mBio 6 (6): e01348-15. Crossref
Cloud, P.E., Jr. and Boucot, A.J. 1971. Dzieduszyckia in Nevada. In: J.T. Dutro, Jr. (ed.), Paleozoic Perspectives, a Tribute to G. Arthur Cooper. Smithsonian Contributions to Paleobiology 3: 175–180.
Cohen, B.L. and Bitner, M.A. 2013. Molecular phylogeny of Rhynchonellide articulate brachiopods (Brachiopoda, Rhynchonellida). Journal of Paleontology 87: 211–216. Crossref
Danise, S. and Higgs, N.D. 2015. Bone-eating Osedax worms lived on Mesozoic marine reptile deadfalls. Biology Letters 11 (4): A20150072. Crossref
Diener, C. 1903. Permian fossils of the Central Himalayas. Palaeontologia Indica (Series 15) 1 (5): 1–204.
Domeier, M. and Torvisk, T.H. 2014. Plate tectonics in the late Paleozoic. Geoscience Frontiers 5: 303–350. Crossref
Donkervoort, L., Presita, A., Griffiths, R., Bowell, R.J., Fallowfield, K., and Barr, M. 2022. Environmental geology of barite deposits in Nevada. Economic Geology and the Bulletin of the Society of Economic Geologists 18: 517–584. Crossref
Dubé, T.E. 1988. Tectonic significance of Upper Devonian igneous rocks and bedded barite, Roberts Mountains Allochthon, Nevada, U.S.A. In: N.J. McMillan, A.F. Embry, and D.J. Glass (eds.), Devonian of the World. Canadian Society of Petroleum Geologists Memoir 14: 235–249.
Duméril, A.M.C. 1805. Zoologie analytique, ou méthode naturelle de classification des animaux, rendue plus facile à l’aide de tableaux synoptiques. 344 pp. Allais, Paris. [imprint date 1806]. Crossref
El Hassani, A. and Benfrika, E.M. 2000. The Devonian of the Moroccan Meseta: biostratigraphy and correlations. Courier Forschungsinstitut Senckenberg 225: 195–209.
Ferrier-Pagès, C. and Leal, M.C. 2018. Stable isotopes as tracers of trophic interactions in marine mutualistic symbioses. Ecology and Evolution 9: 723–740. Crossref
Finney, S.C., Perry, B.D., Emsbo, P., and Madrid, R.J. 1993. Stratigraphy of the Roberts Mountains Allochthon, Roberts Mountains and Shoshone Range, Nevada. In: M.M. Lahren, J.H. Trexler, Jr., and C. Spinosa (eds.), Crustal Evolution of the Great Basin and the Sierra Nevada, 197–230. University of Nevada, Department of Geological Sciences, Reno.
Gatovsky, Y.A., Nikolaeva, S.V., Gorozhanina, E.N., Kulagina, E.I., Gorozhanin, V.M., Alekseev, A.S., Mizens A.G, and Mizens, L.I. 2017. The Dzerzhinka section (eastern slope of the South Urals): an overview of the fauna and facies around the Devonian–Carboniferous boundary. Stratigraphy 14: 143–178. Crossref
Gemmellaro, G.G. 1871. Studi paleontologici sulla fauna del calcaire a Terebratula janitor del nord de Sicilia. Giornale de Scienze Naturali de Economiche di Palermo 7: 73–108.
Gilluly, J. and Gates, O. 1965. Tectonic and igneous geology of the northern Shoshone Range, Nevada. U.S. Geological Survey Professional Paper 465: 1–153. Crossref
Gilluly, J. and Masursky, H. 1965. Geology and ore deposits of the Cortez quadrangle, Nevada: U.S. Geological Survey Bulletin 1175: 1–105.
Gischler, E., Sandy, M.R., and Peckmann, J. 2003. Ibergirhynchia contraria (F.A. Roemer, 1850), an Early Carboniferous seep-related rhynchonellide brachiopod from the Harz Mountains, Germany—a possible successor to Dzieduszyckia? Journal of Paleontology 77: 293–303. Crossref
Golionko, B.G. and Ryazantsev, A.V. 2021. Deformation and structural evolution of metamorphic complexes of the Taldyk antiform in the East Mugodzhar Zone of Urals (West Kazakhstan) [in Russian]. Geodinamika i Tektonofizika 12: 48–59. Crossref
Golonka, J., Ross, M.L., and Scotese, C.R. 1994. Phanerozoic paleogeographic and paleoclimatic modeling maps. In: A.F. Embry, B. Beauchamp, and D.J. Glass (eds.), Pangea: Global Environments and Resources. Canadian Society of Petroleum Geologists, Memoir 17: 1–47.
Gorozhanina, E.N., Pazukhin, V.N., and Puchkov, V.N. 2010. The paleogeodynamic conditions of redeposition of conodont elements in the Late Devonian–Early Carboniferous sediments of the Southern Urals [in Russian]. Doklady Akademii Nauk 432: 560–564. Crossref
Gruzdev, D.A. 2021. Late Devonian–early Carboniferous isolated carbonate platforms of the north of the Urals and Pay-Khoy [in Russian]. Vestnik Geonauk 2021 (10): 3–15. Crossref
Guo, Z., Chen, Z.-Q., Harper, D.A.T., and Huang, Y. 2022. Permian–Triassic phylogenetic and morphologic evolution of rhynchonellide brachiopods. Paleobiology 48: 99–119. Crossref
Hoepffner, C., Soulaimani, A., and Piqué, A. 2005. The Moroccan Hercynides. Journal of African Earth Sciences 43: 144–165. Crossref
Holdsworth, B.K. 1980. Preliminary radiolarian zonation for Late Devonian through Permian time. Geology 8: 281–285. Crossref
Holdsworth, B.K. and Jones, D.L. 1980. A provisional radiolaria biostratigraphy, Late Devonian through Late Permian. U.S. Geological Survey Open-file Report 80-876: 1–32. Crossref
Hollard, H. and Morin, P. 1973. Les gisements de Dzieduszyckia (Rhynchonellida) du Famennien inférieur du massif Hercynien central du Maroc. Service Géologique du Maroc, Notes et Mémoires 249: 7–14.
Hollard, H., Lys, M., Mauvier, A., and Morin, P. 1970. Précisions sur l ́age Famennien inférieur des Dzieduszyckia (Rhynchonellida) du Massif Hercynien central du Maroc. Comptes Rendus de l ́Academie des Sciences Paris, Série D 270: 3177–3180.
Holm-Denoma, C.S., Hofstra, A.H., Noble, P.J., and Leslie, S.A. 2011. Paleozoic stratigraphy and kinematics of the Roberts Mountains Allochthon in the Independence Mountains, northern Nevada. In: R. Steininger and B. Pennell (eds.), Great Basin Evolution and Metallogeny, 1039–1054. DEStech Publications, Lancaster.
Holm-Denoma, C.S., Hofstra, A.H., Rockwell, B.W., and Noble, P.J. 2017. The Valmy thrust sheet: a regional structure formed during the protracted assembly of the Roberts Mountains allochthon, Nevada, USA. Geological Society of America Bulletin 129: 1521–1536. Crossref
Hryniewicz, K. 2022. Ancient hydrocarbon seeps of the world. In: A. Kaim, J.K. Cochran, and N.H. Landman (eds.), Ancient Hydrocarbon Seeps. Topics in Geobiology 53: 571–635. Crossref
Ivanov, K.S. and Puchkov, V.N. 2020. Stratigraphy and tectonics of the Zilair Synclinorium, Urals [in Russian]. Izvestiya Ural’skogo Gosudarstvennogo Gornogo Universiteta 60 (4): 67–72. Crossref
Jakubowicz, M., Berkowski, B., Hryniewicz, K., and Belka, Z. 2022. Middle Palaeozoic of Morocco: the earliest-known methane seep metazoan ecosystems. In: A. Kaim, J.K. Cochran, and N.H. Landman (eds.), Ancient Hydrocarbon Seeps. Topics in Geobiology 53: 479–516. Crossref
Kaim, A. 2022. A review of gastropods at ancient hydrocarbon seeps. In: A. Kaim, J.K. Cochran, and N.H. Landman (eds.), Ancient Hydrocarbon Seeps. Topics in Geobiology 53: 323–374. Crossref
Ketner, K.B. 2013. Stratigraphy of lower to middle Paleozoic rocks of northern Nevada and the Antler orogeny. U.S. Geological Survey Professional Paper 1799: i–v + 1–23. Crossref
Kiel, S. and Peckmann, J. 2008. Paleoecology and evolutionary significance of an Early Cretaceous Peregrinella-dominated hydrocarbon-seep deposit on the Crimean Peninsula. Palaios 23: 751–759. Crossref
Kiel, S. and Peckmann, J. 2019. Resource partitioning among brachiopods and bivalves at ancient hydrocarbon seeps: a hypothesis. PLoS One 14 (9): e0221887. Crossref
Kiel S., Glodny J., Birgel D., Bulot L.G., Campbell K.A., Gaillard C., Graziano R., Kaim A., Lazar I., Sandy M.R., and Peckmann J. 2014. The paleoecology, habitats, and stratigraphic range of the enigmatic Cretaceous brachiopod Peregrinella. PLoS One 9 (10): e0109260. Crossref
Kiel, S., Huemer, J., Gussone, N., Berndt, J., Krystyn, L., Zuschin, M., and Peckmann, J. 2021. Brachiopods in early Mesozoic cryptic habitats: continuous colonization, rapid adaptation, and wide geographic distribution. Palaeogeography, Palaeoclimatology, Palaeoecology 583: 110668. Crossref
Koski, R.A. and Hein, J.R. 2003. Stratiform barite deposits in the Roberts Mountains Allochthon, Nevada: a review of potential analogs in modern sea-floor environments. U.S. Geological Survey Bulletin 2209-H: 1–10.
Kuhn, O. 1949. Lehrbuch der Paläozoologie. 326 pp. E. Schweizerbart, Stuttgart.
Levin, L.A., Mendoza, G.F., Gonzalez, J.P., Thurber, A.R., and Cordes, E.E. 2010. Diversity of bathyal macrofauna on the northeastern Pacific margin: the influence of methane seeps and oxygen minimum zones. Marine Ecology 31: 94–110. Crossref
Lipina, O.A. 1955. Foraminifera of the Tournaisian stage and the upper part of the Devonian in the Volga-Urals region and the western slopes of the Middle Urals [in Russian]. Trudy Instituta Geologii Akademii Nauk SSSR 163: 1–96.
Little, C.T.S., Danelian, T., Herrington, R.J., and Haymon, R.M. 2004. Early Jurassic hydrothermal vent community from the Franciscan Complex, California. Journal of Paleontology 78: 542–559. Crossref
Madrid, R.J., Poole, F.G., and Wrucke, C.T. 1992. Rocks of the Antler Orogen—the Roberts Mountains Allochthon. In: B.C. Burchfiel, P.W. Lipman, and M.L. Zoback (eds.), The Cordilleran Orogen, Conterminous U.S., The Geology of North America G-3, 28–34. Geological Society of America, Boulder.
Manceñido, M.O. 2000. A systematic summary on the stratigraphic distribution of Jurassic rhynchonellide genera (Brachiopoda). In: R.L. Hall and P.L. Smith (eds.), Advances in Jurassic Research 2000. Proceedings of the Fifth International Symposium on the Jurassic System (Vancouver, Canada, August 1998), 387–396. GeoResearch Forum 6, Trans Tech Publications, Zurich.
Manceñido, M.O. and Owen, E.F. 1996. Post-Paleozoic rhynchonellides: an overview. In: P. Copper and J. Jin (eds.), Brachiopods. Proceedings of the Third International Brachiopod Congress 1995, 189–200. A.A. Balkema, Rotterdam.
Manceñido, M.O. and Owen, E.F. 2001. Post-Palaeozoic Rhynchonellida (Brachiopoda): classification and evolutionary background. In: C.H.C. Brunton, L.R.M. Cocks, and S.L. Long (eds.), Brachiopods Past and Present. The Systematics Association Special Volume Series 63: 189–200. Crossref
Manceñido, M.O., Owen, E.F., Savage, N.M. and Dagys, A.S. 2002. Dimerelloidea. In: R.L. Kaesler (ed.), Treatise on Invertebrate Paleontology. Part H, Brachiopoda, Revised, Volume 4: Rhynchonelliformea, 1236–1245. Geological Society of America and The University of Kansas, Boulder and Lawrence.
Markovsky, A.P. 1937. The Kshtut River Basin (the middle part of the Zeravshan range) [in Russian]. In: V.G. Mukhin et al. (eds.), Geologiya Uzbekskoy SSR 2, 175–195. ONTI: NKTP, Leningrad.
Menakova, G.N. 1991. Brachiopods. In: M.R. Dzhalilov (ed.), Atlas of the Fossil Fauna and Flora of Tadzhikistan, Ordovician, Silurian, Devonian [in Russian], 177–200. Donish, Dushanbe.
Miller, E.L., Holdsworth, B.K., Whiteford, W.B., and Rodgers, D. 1984. Stratigraphy and structure of the Schoonover Sequence, northeastern Nevada: Implications for Paleozoic plate-margin tectonics. Geological Society of America Bulletin 95: 1063–1076. Crossref
Mizens, A.G. 2006. First findings of representatives of the genera Zilimia and Dzieduszyckia (Brachiopoda) in the Famennian limestones near Verkhne-Uralsk (eastern slope of the southern Urals) [in Russian]. In: V.A. Koroteyev (ed.), Institut Geologii i Geokhimii im. Akad. A.N. Zavaritskogo; informatsionnyy sbornik nauchnykh trudov, Rossiyskaya Akademiya Nauk, Ural’skoye Otdeleniye, Yekaterinburg, Rossiy, Yezhegodnik 2005: 28–33.
Mizens, A.G. 2008. Lower Famennian brachiopods of the genus Dzieduszyckia from the Gubakha Formation, western slope, Middle Urals [in Russian]. In: V.A. Koroteyev (ed.), Institut Geologii i Geokhimii im. Akad. A.N. Zavaritskogo; informatsionnyy sbornik nauchnykh trudov, Rossiyskaya Akademiya Nauk, Ural’skoye Otdeleniye, Yekaterinburg, Rossiy, Yezhegodnik 2007: 53–60.
Mizens, A.G. 2012. Biostratigraphic division of Givetian–Famennian deposits of the Middle and South Urals by brachiopods [in Russian]. Litosfera 2: 43–54.
Mizens, A.G., Abramova, A.N., and Artyushkova, O.V. 2003. First findings of Famennian brachiopods of Dzieduszyckia Siemiradzki genus on western side of the Zalair Megasynclinorium) [in Russian]. In: V.A. Koroteyev (ed.), Institut Geologii i Geokhimii im. Akad. A.N. Zavaritskogo; informatsionnyy sbornik nauchnykh trudov, Rossiyskaya Akademiya Nauk, Ural’skoye Otdeleniye, Yekaterinburg, Rossiy, Yezhegodnik 2002: 34–38.
Mizens, A.G. and Mizens, L.I. 2017. Stratigraphic significance of Devonian brachiopods of the Urals [in Russian]. Litosfera 17 (4): 62–72. Crossref
Nalivkin, D.V. 1947. Class Brachiopoda [in Russian]. In: D.V. Nalivkin and V.N. Verber (eds.), Atlas rukovodâŝih form iskopaemykh faun SSSR. Tom 3. Devonskaya sistema, 63–134. VSEGEI-Gosgeoltehizdat, Leningrad.
Nie, T., Guo, W., Sun, Y.L., Shen, B., Yin, B.-A. Tang, Z.-H., Li, Y.-K., Huang, X.-L., and Mai, C. 2016. Age and distribution of the Late Devonian brachiopod genus Dzieduszyckia Siemiradzki, 1909 in southern China. Palaeoworld 25: 600–615. Crossref
Nikolaeva, S.V., Kulagina, E.I., Mustapayeva, S.N., Alekseev, A.S.. Gatovsky, Y.A., Denayer, J., Ohar, V., Kurilenko, A.V., Zhaimina, V.J., and Mychko, E.V. 2022. Progress in the study of the Devonian–Carboniferous boundary sections in the Berchogur Depression (Mugodzhary Mountains, western Kazakhstan). Paläontologische Zeitschrift 96: 415–447. Crossref
Noll, J.H. 1981. Geology of the Picacho Colorado Area, Northern Sierra de Cobachi, Central Sonora, Mexico. 169 pp. Unpublished M.Sc. Thesis, Northern Arizona University, Flagstaff.
Noll, J.H., Dutro, J.T., Jr., and Beus, S.S. 1984. A new species of the Late Devonian (Famennian) brachiopod Dzieduszyckia from Sonora, Mexico. Journal of Paleontology 58: 1412–1421.
Palfy, J., Kovacs, Z., Price G.D., Voros, A., and Johannson, G.G. 2017. A new occurrence of the Early Jurassic brachiopod Anarhynchia from the Canadian Cordillera confirms its membership in chemosynthesis-based ecosystems. Canadian Journal of Earth Sciences (Revue Canadienne des Sciences de la Terre) 54: 1179–1193. Crossref
Pawłowska, K. and Pawłowski, S. 1978. Charakterystyka utworów paleozoicznych (karbon, dewon) na podstawie otworu wiertniczego w Rudzie Strawczyńskiej. Kwartalnik Geologiczny 22: 679–696.
Peckmann, J., Campbell, K.A., Walliser, O.H., and Reitner, J. 2007. A Late Devonian hydrocarbon-seep deposit dominated by dimerelloid brachiopods, Morocco. Palaios 22: 114–122. Crossref
Peckmann, J., Kiel, S., Sandy, M.R., Taylor, D.G., and Goedert, J.L. 2011. Mass occurrences of the brachiopod Halorella in Late Triassic methane-seep deposits, eastern Oregon. Journal of Geology 119: 207–220. Crossref
Peckmann, J., Sandy, M.R., Taylor, D.G., Gier, S., and Bach, W. 2013. An Early Jurassic brachiopod-dominated seep deposit enclosed by serpentinite, eastern Oregon, USA. Palaeogeography, Palaeoclimatology, Palaeoecology 390: 4–16. Crossref
Pereira, O.S., Gonzalez, J., Mendoza, G.F., Le, J., Coscino, C.L., Lee, R.W., Cortés, J., Cordes, E.E., and Levin, L.A. 2021. The dynamic influence of methane seepage on macrofauna inhabiting authigenic carbonates. Ecosphere 12 (10): e03744. Crossref
Poole, F.G. 1974. Flysch deposits of Antler foreland basin, western United States. In: W.R. Dickinson (ed.), Tectonics and Sedimentation. Society of Economic Paleontologists and Mineralogists Special Publication 22: 58–82. Crossref
Pusch, G.G. 1833. Geognostische Beschreibung von Polen sowie der übrigen Nordkarpathen-Länder. Erster Theil. 388 pp. J.G. Cotta’schen Buchhandlung, Stuttgart.
Reed, F.R.C. 1944. Brachiopoda and Mollusca from the Productus limestones of the Salt Range. Palaeontogica Indica, Memoirs of the Geological Survey of India (new series) 23 (2): 1–596.
Reershemius, T. and Planavsky, N.J. 2021. What controls the duration and intensity of ocean anoxic events in the Paleozoic and the Mesozoic? Earth-Science Reviews 221: 103787. Crossref
Roemer, F.A. 1866. Geognostische Beobachtungen im Polnischen Mittergebirge. Zeitschrift der Deutsche geologische Gesselschaft 18: 667–690.
Rozman, K.S. 1960. The stratigraphy of the Famennian and lower Tournaisian deposits in the Mugodzhary and adjacent areas of the Urals [in Russian]. Izvestiya Akademii Nauk SSSR, Seriya geologicheskaya 12: 35–45.
Rozman, K.S. 1962. Stratigraphy and brachiopods of the Famennian stage of the Mugodzhary and adjacent areas [in Russian]. Trudy Geologičeskogo Instituta Akademii Nauk SSSR 50: 1–195.
Sabirov, A. 2019. Definition of foraminiferal zones in the Famennian stage (Upper Devonian) of Tajikistan. Journal of Foraminiferal Research 49: 163–177.
Sandy, MR. 2010. Brachiopods from ancient hydrocarbon seeps and hydrothermal vents. In: S. Kiel (ed.), The Vent and Seep Biota. Topics in Geobiology 33: 279–314. Crossref
Sandy, M.R. and Campbell, K.A. 1994. New rhynchonellid brachiopod genus from Tithonian (Upper Jurassic) cold seep deposits of California and its paleoenvironmental setting. Journal of Paleontology 68: 1243–1252. Crossref
Sandy, M.R. and Peckmann, J. 2016. The Early Cretaceous brachiopod Peregrinella from Tibet; a confirmed hydrocarbon-seep occurrence for a seep-restricted genus. Palaeontologische Zeitschrift 90: 691–699. Crossref
Sandy, M.R., Lazăr, I., Peckmann, J., Birgel, D., Stoica, M., and Roban, R.D. 2012. Methane-seep brachiopod fauna within turbidites of the Sinaia Formation, Eastern Carpathian Mountains, Romania. Palaeogeography, Palaeoclimatology, Palaeoecology 323–325: 42–59. Crossref
Sartenaer, P. 1959. Leiorhynchus or Nudirostra. Journal of Paleontology 33: 964–965.
Savage, N.M. 1996. Classification of Paleozoic rhynchonellid brachiopods. In: P. Copper and J. Jin (eds.), Brachiopods. Proceedings of the Third International Brachiopod Congress, 249–260. A.A. Balkema, Rotterdam. Crossref
Savage, N.M., Owen, E.F., Manceñido, M.O., and Dagys, A.S. 2002. Part H, Brachiopoda (revised), vol. 4, ch. 3. In: R.L. Kaesler (ed.), Treatise on Invertebrate Paleontology. Part H, Brachiopoda, Revised, 1132–1218. Geological Society of America and The University of Kansas, Boulder and Lawrence. Crossref
Siemiradzki, J. 1909. Zbiory L. Zejsznera z kieleckiego dewonu. Sprawozdanie Komisji Fizyograficznej. Akademia Umiejętności 43: 62–94.
Silberling, N.J., Nichols, K.M., Macke, D.L., and Trappe, J. 1995. Upper Devonian–Mississippian stratigraphic sequences in the distal Antler foreland of western Utah and adjoining Nevada. U.S. Geological Survey Bulletin 1988-H: 1–33.
Spalletta, C., Perri, M.C., Over, D.J., and Corradini, C. 2017. Famennian (Upper Devonian) conodont zonation: revised global standard. Czech Geological Survey, Prague Bulletin of Geosciences 92: 31–57. Crossref
Speed, R.C. and Sleep, N.H. 1982. Antler orogeny and foreland basin—a model. Geological Society of America Bulletin 93: 815–828. Crossref
Stigall, A.L. 2012. Speciation collapse and invasive species dynamics during the Late Devonian “Mass Extinction”. GSA Today 22 (1): G128A.1. Crossref
Sulser, H. and Furrer, H. 2008. Dimerelloid rhynchonellide brachiopods in the Lower Jurassic of the Engadine (Canton Graubünden National Park, Switzerland). Swiss Journal of Geosciences 101: 203–222. Crossref
Tagarieva, R. 2012. Conodont biodiversity of the Frasnian–Famennian boundary interval (Upper Devonian) in the Southern Urals. Bulletin of Geosciences 88: 297–314. Crossref
Termier, H. 1936. Etudes géologiques sur le Maroc central et le Moyen−Atlas septentrional. Tome III. Paléontologie, Pétrographie. Service Géologique du Maroc, Notes et Mémoires 33: 1087–1421.
Termier, H. 1938. Sur l’existence des Halorella au Dévonien supérieur. Compte Rendu sommaire des Séances de la Société Géologique de France 7: 1–108.
Termier, H. and Termier, G. 1948. Les phénomènes de spéciation dans le genre Halorella. Service Géologique du Maroc, Notes et Mémoires 71: 47–63.
Termier, H. and Termier, G. 1949. Sur les genres Halorella et Dzieduszyckia. Service Géologique du Maroc, Notes et Mémoires 74: 113–115.
Tong, L., Dong, L., Yuan, Z.-W., Ling, K., Ning, M., Qin, S.-J., Yang, R.-Y., Liu, W., Nie, T., Li, C.-Q., and Sun, Y.-L. 2021, Taphonomic features and population dynamics of the Upper Devonian Famennian Dzieduszyckia fauna in South China [in Chinese]. Gudili Xuebao (Journal of Paleogeography) 23 (6): 1210–1224.
Torres, M.E., Bohrmann, G, Dubé, T.E., and Poole, F.G. 2003. Formation of modern and Paleozoic stratiform barite at cold methane seeps on continental margins. Geology 31: 897–900. Crossref
Tschernyschew, T.N. 1887. Fauna of the middle and upper Devonian of western slope of the Urals [in Russian].Trudy Geologičeskogo Komiteta 3: 1–208.
Tsyganko, V.S. 2011. Devon zapadnogo sklona severa Urala i Pai-Khoia (stratigrafiâ, printsipy raschleneniâ, korrelyatsiâ). 356 pp. UrO RAN, Ekaterinburg.
Van Dover, C.L., German, C.R., Speer, K.G., Parson, L.M., and Vrijenhoeka, R.C. 2002. Evolution and biogeography of deep-sea vent and seep invertebrates. Science 295: 1253–91257. Crossref
Walliser, O.H., El Hassani, A., and Tahiri, A. 2000. Mrirt: A key area for the Variscan Meseta of Morocco. Service Géologique du Maroc, Notes et Mémoires 399: 93–108.
Williams, A., Carlson, S.J., and Brunton, C.H.C. 2000. Rhynchonelliformea. In: R.L. Kaesler (ed.), Treatise on Invertebrate Paleontology, Vols. 2, 3, Pt. H, Brachiopoda, revised, 193–902. Geological Society of America and University of Kansas, Boulder and Lawrence.
Williams, A., Carlson, S., Brunton, C.H.C., Holmer, L.E., and Popov, L.E. 1996. A supra-ordinal classification of the Brachiopoda. Philosophical Transactions of the Royal Society, Biological Sciences 351: 1117–1193. Crossref
Xian, S.Y. and Jiang, Z.L. 1978. Brachiopoda [in Chinese]. In: Stratigraphic and Paleontological Working Team of Guizhou Province (ed.), Paleontological Atlas of Western China (Guizhou Province), Volume 1, 251–337. Geological Publishing House, Beijing.
Xian, S.Y., Wang, S.D., Zhou, X.Y., Xiong, J.F., and Zou, T.R. 1980. Stratigraphy and Paleontology of Nandan-type Devonian in South China [in Chinese]. 161 pp. People’s Press of Guizhou Province, Guiyang.
Yudina, Y.A. 1997. Rhynchonellids (Brachiopoda) from the Barma deposits of the western slope of the South Urals [in Russian]. Paleontological Journal 5: 63–67.
Yudina, A.B., Racki, G., Savage, N.M., Racka, M., and Małkowski, K. 2002. The Frasnian–Famennian events in a deep-shelf succession, Subpolar Urals: biotic, depositional and geochemical records. Acta Palaeontologica Polonica 47: 355–372.
Zhuravlev, A.V. and Sokiran, E.V. 2020. Frasnian–Famennian (Upper Devonian) transition in the northern hemisphere (NE Laurussia and NE Siberia)—an overview. Bulletin of Geosciences 95: 419–439. Crossref
Ziegler, W. and Sandberg, C.A. 1990. The Late Devonian standard conodont zonation. Courier Forschungsinstitut Senckenberg 121: 1–115.
Zittel, K.A. 1870. Ueber den Brachial-Apparat bei einigen jurassischen Terebratuliden und über eine neue Brachiopoden-Gattung Dimerella. Palaeontographica 17 (5): 211–222.
Acta Palaeontol. Pol. 69 (1): 87–107, 2024
https://doi.org/10.4202/app.01059.2023