
Immature skulls of the theropod dinosaur Coelophysis bauri from Ghost Ranch, New Mexico
JEB E. BUGOS and SKYE N. MCDAVID
Bugos, J.E. and McDavid, S.N. 2024. Immature skulls of the theropod dinosaur Coelophysis bauri from Ghost Ranch, New Mexico. Acta Palaeontologica Polonica 69 (4): 549–564.
The theropod dinosaur Coelophysis bauri is well known from the famous Rhaetian, Late Triassic mass death assemblage from the Coelophysis Quarry at Ghost Ranch, New Mexico. This research describes the skulls of two small individuals of C. bauri. Both skulls are mostly complete, though they are missing premaxillae, and most posterior skull bones are either missing or badly damaged. Both specimens preserve hyoids, making them excellent candidates for hyoid histology. These two skulls have different proportions from other Coelophysis skulls from Ghost Ranch, especially in the large size of the orbit, short and flat triangular teeth, and thinner bones. These differences are interpreted as representing ontogenetic variation, with these skulls representing skeletally immature, partially developed individuals.
Key words: Theropoda, Triassic, Coelophysis, ontogeny, Chinle Formation.
Jeb E. Bugos [jbugos@cmnh.org; ORCID: https://orcid.org/0000-0003-2336-1442 ],Case Western Reserve University, 10900 Euclid Avenue, Cleveland, Ohio 44106 USA; current address: Cleveland Museum of Natural History, 1 Wade Oval Drive, Cleveland, Ohio 44106 USA.
Skye N. McDavid [mail@skyemcdavid.com; ORCID: https://orcid.org/0000-0003-2529-1812 ] corresponding author, Delaware Valley Paleontological Society, PO Box 686, Plymouth Meeting, Pennsylvania 19462 USA.
Received 9 June 2024, accepted 20 September 2024, published online 30 October 2024.
Copyright © 2024 J.E. Bugos and S.N. McDavid. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Discovered in 1947 by Edwin Colbert’s AMNH field team near Abiquiú in Rio Arriba County, Northern New Mexico (Fig. 1), the Ghost Ranch Coelophysis Quarry, also known as the Whitaker Quarry (e.g., Lucas et al. 2005; Rinehart et al. 2009), is a remarkable site representing a mass death assemblage of Late Triassic animals, primarily the approximately 2-meter-long theropod dinosaur Coelophysis bauri (Schwartz and Gillette 1994). Following its original discovery, two phases of excavations removed nearly 30 blocks of material which were distributed among repositories across the USA and Canada (Colbert 1989). The first period of excavation led by the American Museum of Natural History in 1947 and 1948 provided most of the specimens included in Colbert’s (1989) monographic description of C. bauri, and the second period of excavation in 1981 and 1982 was a joint effort between the Carnegie Museum of Natural History, the Yale Peabody Museum, the Museum of Northern Arizona, and the New Mexico Museum of Natural History and Science (Colbert 1989; Schwartz and Gillette 1994; Rinehart et al. 2009).
Fig. 1. Location of the Ghost Ranch Coelophysis Quarry (asterisk) in New Mexico, USA.
The quarry is located in the Rhaetian (Upper Triassic) “siltstone member” of the Chinle Formation (Colbert 1989; Hunt and Lucas 1989, 1991; Schwartz and Gillette 1994; Zeigler et al. 2008; Marsh and Parker 2020; but see Rinehart et al. 2009 for alternate stratigraphic nomenclature). Schwartz and Gillette (1994) provide a thorough review of the geology and paleoenvironment of the Coelophysis Quarry. The paleoenvironment has been interpreted as semi-arid with seasonal precipitation and monsoons (Schwartz and Gillette 1994; Therrien and Fastovsky 2000; Whiteside et al. 2015), while the quarry itself is a river channel overbank deposit, representing a small fluvial channel that was clogged with animal carcasses and filled with silty sediment during flooding.
The Ghost Ranch Coelophysis Quarry is a unique site due to the large number of individuals of a single species, the quality of preservation, and the catastrophic way in which the death assemblage was formed. The quarry is also exceptional among Chinle Formation and stratigraphically equivalent theropod localities because it likely represents a single, catastrophic event rather than a time-averaged assemblage, and is therefore uniquely useful for gaining insights into behavior and intraspecific variation in Coelophysis (Therrien and Fastovsky 2000). Known from an abundance of material, C. bauri (Cope 1887, 1889) is among the most studied non-avian theropods, and certainly the most studied Triassic theropod, though its anatomy remains under-described. The Ghost Ranch Coelophysis Quarry has produced possibly over a thousand individuals of C. bauri displaying a range of variation that has been attributed to sexual dimorphism (Colbert 1990; Paul 1993; Schwartz and Gillette 1994; Smith and Merrill 2006), ontogenetic variation (Colbert 1989, 1990; Rinehart et al. 2009; Griffin and Nesbitt 2016), and intraspecific variation (Colbert 1989; Griffin and Nesbitt 2016; Barta et al. 2018, 2022). Though specimens from other sites in the region have been referred to C. bauri (Padian 1986; Colbert 1989), these referrals are uncertain, so only the Ghost Ranch Coelophysis Quarry specimens can confidently be assigned to C. bauri (Nesbitt et al. 2007; Marsh and Parker 2020). One other dinosaur has been identified in the Ghost Ranch Coelophysis Quarry sample: Daemonosaurus chauliodus, an early-diverging saurischian known from a single skull and associated partial cervical series (Sues et al. 2011; Nesbitt and Sues 2021). The Ghost Ranch Coelophysis Quarry assemblage also includes phytosaurs (Hunt and Lucas 1993; Goldsmith et al. 2023), drepanosauromorphs (Harris and Downs 2002; Pritchard and Nesbitt 2017), shuvosaurids (Nesbitt 2007), crocodylomorphs (Nesbitt et al. 2006; Rinehart et al. 2009; Nesbitt 2011), sphenodontians (Heckert et al. 2008) and fish (Schaeffer 1967).
Coelophysis bauri has a complicated taxonomic and nomenclatural history, which was summarized by Lucas et al. (2005). The closely related genus Megapnosaurus from Southern Africa, which Raath (1969, 1977) and Colbert (1989) originally considered a separate genus, has sometimes been considered a synonym of Coelophysis (Paul 1988, 1993; Downs 2000; Bristowe and Raath 2004; see also McDavid and Bugos 2022 for nomenclature).
Although the Ghost Ranch Coelophysis Quarry provides an exceptionally large sample of Coelophysis from a variety of ontogenetic stages (Colbert 1989, 1990; Rinehart et al. 2009), no neonates or especially small immature individuals have yet been reported. Colbert’s (1989) claim of juvenile Coelophysis remains preserved in the stomach of an older individual was later shown to represent the remains of indeterminate reptiles and a crocodylomorph rather than Coelophysis (Nesbitt et al. 2006). The skulls described in this paper are significantly smaller than most individuals collected from the Coelophysis Quarry (see e.g., Colbert 1989, 1990; Rinehart et al. 2009). Although they would be categorized as “juveniles” following the terminology of Hone et al. (2016), we refer to skeletal maturity in this study, where maturity is defined as “the animal [having] reached [mature] status under a particular criterion” and immaturity is defined as “not [having] reached this threshold” (modified from Hone et al. 2016: 2). Immature remains of early theropods are rare in the fossil record (Carpenter and Alf 1994; Bever and Norell 2009), so as one of the only examples of ontogenetic series of Triassic theropods, the Ghost Ranch Coelophysis Quarry sample is especially important for understanding plesiomorphic dinosaur development and morphology.
Institutional abbreviations.—AMNH, American Museum of Natural History, New York, USA; CM, Carnegie Museum of Natural History, Pittsburgh, USA; CMNH, Cleveland Museum of Natural History, Cleveland, USA; MCZ, Museum of Comparative Zoology, Harvard University, Cambridge, USA; NHMZ, Natural History Museum of Zimbabwe, Bulawayo, Zimbabwe (QG designates specimens originally accessioned at the Zimbabwe Museum of Human Sciences, Harare, formerly the Queen Victoria Museum, transferred to NHMZ); NMMNH, New Mexico Museum of Natural History and Science, Albuquerque, USA; YPM, Yale Peabody Museum, New Haven, USA.
Material and methods
CMNH 50957 and MCZ VPRA 4326 were collected from the Ghost Ranch Coelophysis Quarry during the original AMNH excavation in 1947–1948 (Colbert 1989). The quarry is located in the informally-defined “siltstone member” of the Upper Triassic Chinle Formation (Schwartz and Gillette 1994; Ziegler et al. 2008; Marsh and Parker 2020; but see Rinehart et al. 2009 for alternative stratigraphic nomenclature) and the matrix surrounding the fossils is a red and light gray siltstone.
One block, originally designated AMNH Block XII (see Colbert 1989: fig 6.2) was sent to the Cleveland Museum of Natural History, where it is cataloged as CMNH 10971. This block contained skeletons of at least 14 individuals (Lee Hall, personal communication 2020) referred to C. bauri, as well as phytosaur material (Goldsmith et al. 2023; JEB personal observations).
Five C. bauri skulls of varying completeness have been recovered from Block XII, and one additional partial skull including skull roof, maxilla, and dentary elements with teeth has been identified in situ in the block but remains unprepared. Of the skulls removed from the block, the largest is CMNH 11882, a nearly complete, dorsoventrally compressed skull with a crushed premaxilla, which is 196.9 mm long. Based on its larger and more robust bones, CMNH 50955, an incomplete and highly distorted skull missing many posterior skull elements, is possibly from a larger individual than CMNH 11882 despite the specimen itself being smaller at 153.8 mm in length. However, given the high intraspecific variability of growth trajectories of dinosaurs generally (Griffin and Nesbitt 2016; Hone et al. 2016; Griffin et al. 2021) and C. bauri specifically (Griffin and Nesbitt 2016; Barta et al. 2022), size alone is not a reliable proxy for ontogenetic age. CMNH 50956, a broken right maxilla with nine teeth and exposed unerupted replacement teeth, is 97.3 mm long. CMNH 50958 is 87.4 mm long and contains the anterior end of a skull, including the maxilla, premaxilla, dentary, and 39 variously complete teeth from the premaxilla, maxilla, and dentary.
The skulls, along with a significant amount of Coelophysis postcranial material and some phytosaur elements, were removed from the block by volunteer fossil preparator Dale Zelinski at the CMNH Paleo Lab. It is unknown from where in the block CMNH 50957 was removed or if there were postcranial skeletal elements associated with the skull other than cervical ribs and vertebrae found articulated with the skull. These vertebrae were broken during storage and left unattached to avoid further damage. The authors’ attempts to identify any additional postcranial material associated with CMNH 50957 based on notes and photographs taken during preparation of the block were unsuccessful.
MCZ VPRA 4326 was extracted from AMNH Block X (Colbert 1989). The authors’ attempts to locate preparation records at MCZ and AMNH were unsuccessful. It is likely that it was prepared at the AMNH in the 1950s (Downs 2000) before it became standard practice to take notes during fossil preparation. If preparation notes were taken, they are presumed lost. It is unknown whether MCZ VPRA 4326 was associated with any postcranial material. One small skull from Block X, MCZ VPRA 4333, is associated with a right manus and partial cervical series. However, this skull is badly crushed and largely uninformative. MCZ VPRA 4328 is an isolated posterior portion of a maxilla of a small individual and an isolated quadrate of a similarly-sized individual, both of which are partially covered by matrix. MCZ VPRA 4327 includes both a large, complete, and well-preserved mature or nearly-mature skull and disarticulated cranial elements of a smaller individual, including a right jugal, possible quadratojugal and quadrate, possible hyoid, and possible frontals. It should be noted that the identification of disarticulated elements other than the jugal in MCZ VPRA 4327 is highly tentative due to the preservation and preparation of this specimen.
Additional skulls and skull casts from the AMNH, CMNH, MCZ, and YPM collections were examined directly by the authors for comparative purposes. Additional comparisons to material not in these collections were made based on casts, published literature and photographs provided to the authors.
Description
General description and comments.—CMNH 50957 (Fig. 2) is a small, mostly complete, mostly articulated skull of a skeletally immature Coelophysis bauri, including the lower jaw and hyoids but lacking sclerotic rings, associated with cervical ribs and vertebrae. The length of the skull is 96.7 mm long as preserved. As is common in skulls from this quarry, the jaws are closed very tightly (Colbert 1989), obscuring some of the lower jaw and dentition. The skull is mediolaterally compressed, also common for Coelophysis skulls from this site, although distortion of this skull is minor in comparison to some others from the quarry (e.g., MCZ VPRA 4333; Colbert 1989: 55). As noted by Rinehart et al. (2009: 103), skulls are more susceptible to distortion than other skeletal elements in Ghost Ranch C. bauri specimens. The skull is visible in lateral view from both the right and left sides and in dorsal view, although the left side is badly distorted and partially obscured by matrix containing disarticulated vertebrae. Most of this description will refer to the right side of the skull due to its better exposure and preservation.
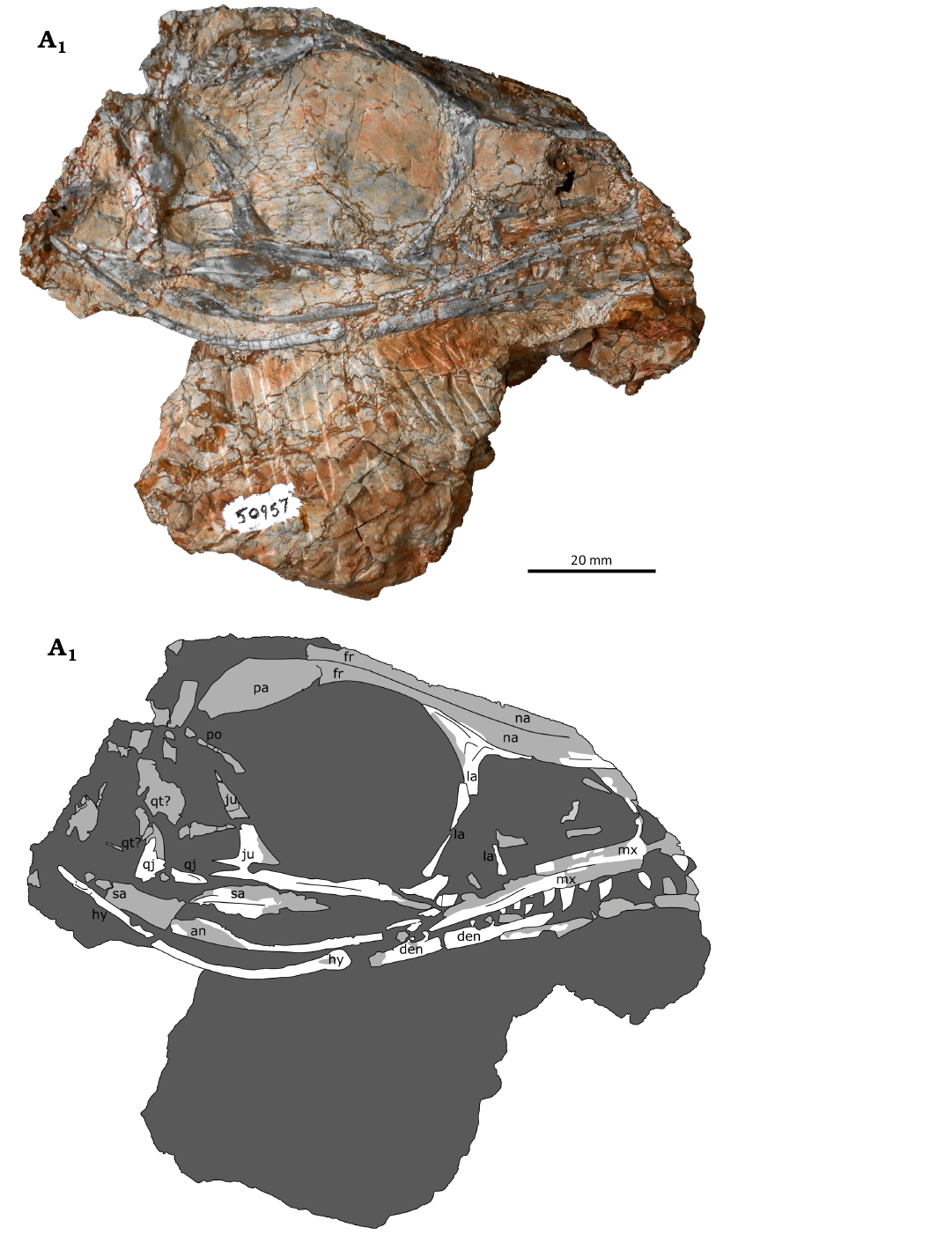
Fig. 2. Immature skull of theropod dinosaur Coelophysis bauri (Cope, 1887), CMNH 50957, from the Rhaetian, Upper Triassic of Ghost Ranch, New Mexico, USA, in right lateral view; photograph (A1) and interpretive line drawing (A2). Damaged bone is indicated in medium gray, matrix is in dark gray. Abbreviations: an, angular; den, dentary; fr, frontal; hy, hyoid; ju, jugal; la, lacrimal; mx, maxilla; na, nasal; pa, parietal; po, postorbital; qj, quadratojugal; qt, quadrate; sa, surangular.
The skull has sustained some taphonomic damage, and some elements are missing. Both the posterior and anterior ends of the skull are broken, with at least part of the parietal and the supraoccipital missing posteriorly. Many of the bones on the posterior end of the skull are badly damaged and disarticulated. Anteriorly, both premaxillae are entirely missing, as are the anteriormost portions of the maxillae, dentaries, and some teeth, although the anterior margin of the small antorbital fossa is preserved on the right side. The antorbital fenestra is present within the antorbital fossa, but its precise borders are obscured by matrix. The right side of the skull is otherwise well preserved.
The skull is very lightly built from thin bones, and in lateral view the openings of the skull have a larger area than the bone itself. The orbit is proportionately large at 28.4 mm, measured at the widest point dorsoventrally. The posterior border of the orbit is displaced, preventing an accurate measurement along the anteroposterior axis. It is bordered dorsally by the frontals, anteriorly by the lacrimal, ventrally by the jugal, and posteriorly by the jugal and postorbital. The antorbital fossa is relatively short, 19.9 mm anteroposteriorly and 14.4 mm tall dorsoventrally. The maxilla makes up its anterior and ventral borders, and the posterior and dorsal borders are defined by the lacrimal. The border of the lateral temporal fenestra is disarticulated. It is made up of the postorbital, jugal, quadratojugal, and quadrate, all of which are at least somewhat broken. The external naris is not preserved.
MCZ VPRA 4326 (Fig. 3) is mostly complete and partially articulated, including a lower jaw, hyoid fragments, and part of the sclerotic ring. The maxilla, lacrimal, jugal, and lower jaw are especially well preserved. Posterior skull bones are present but badly warped, and many are unidentifiable or obscured by other taphonomically displaced bones. The supraoccipital and foramen magnum are identifiable but damaged. The premaxillae and premaxillary teeth are missing. The articulation between the premaxillae and maxillae of coelophysoids has been described as “loose” (Tykoski 1998), possibly explaining the lack of premaxillae in both specimens: loosely attached premaxillae may separate from the rest of the skull comparatively easily. The preserved skull length, measured from the anteriormost point of the maxilla to the posteriormost point of the skull is 98.6 mm. Unlike CMNH 50957 and most C. bauri specimens, MCZ VPRA 4326 is preserved with its jaw open, allowing for more detailed study of the lower jaw and dentary teeth. The skull is mediolaterally crushed. After removing the skull from the block, it was attached to a plaster cradle with wax. We made no attempt to remove the fragile specimen from the cradle, and as such it is only fully visible in left lateral view, with parts of the skull visible in posterior and dorsal view. This description therefore refers almost entirely to the left side of the skull.
Maxilla.—The maxilla of CMNH 50957 (Fig. 2) is a thin, L-shaped bone made up of two long processes that form the anterior and ventral borders of the antorbital fenestra. The maxilla of MCZ VPRA 4326 is better preserved than that of CMNH 50957, and the ascending process of the maxilla curves to form part of the dorsal border of the antorbital fenestra. The anterior border of the antorbital fenestra is approximately rectangular in MCZ VPRA 4326, unlike in more mature skulls (CMNH 50955, CMNH 50958, AMNH FARB 7224, AMNH FARB 30638, YPM VP 41196, MCZ VPRA 4327) where it has the shape of a steep parabola. The larger of the two maxillary processes is a horizontal bar that contains the tooth row. The lateral surface of this process contains a longitudinal depression that spans approximately the entire exposed length of the tooth row. This feature is present in both specimens, but especially apparent in MCZ VPRA 4326, where there appears to be a bulge or rim close to the ventral border of the maxilla and on the ventral and anterior borders of the antorbital fenestra. A proportionally deeper longitudinal depression is also present in larger, more mature C. bauri skulls (CMNH 50955, CMNH 50958, AMNH FARB 7224, YPM VP 41196). It is especially prominent in AMNH FARB 7224, though in this specimen it may have been exaggerated by taphonomic distortion.
The right maxilla of CMNH 50957 is 44.0 mm long as preserved, but the maxilla is broken on its anterior end, close to where it would articulate with the premaxilla. Posteriorly, the long horizontal process of the maxilla extends to approximately the border between the orbit and the antorbital fenestra. The contact between this process of the maxilla and the ventral end of the lacrimal is obscured by matrix. Ventral to this area of matrix, the posterior end of the maxilla is disarticulated from the jugal. The preserved section of the maxillary tooth row is 32.2 mm long and contains 11 teeth, of which 7 are complete.
The left maxilla of MCZ VPRA 4326 (Fig. 3) is nearly complete, with a length of 56.4 mm. Unlike in CMNH 50957, the horizontal process of the maxilla extends posteriorly past the lacrimal and its posteriormost point is located ventral to the orbit. The maxilla articulates with both the lacrimal and the jugal. The ventral end of the vertical process of the lacrimal flares out into a subtriangular footplate, as in CMNH 50955 and possibly AMNH FARB 7224. The ventral border of this footplate articulates with the maxilla in its anterior half, and in its posterior half with the jugal (Fig. 4). Fourteen teeth are preserved in the maxilla of MCZ VPRA 4326, and most of these are complete and in situ (Table 1). There is a suboval area of missing bone on the anterior border of the maxilla close to where the external naris would be if it were preserved in this specimen; this area of missing bone is most likely a case of taphonomic damage or damage during preparation. In mature C. bauri (e.g., YPM VP 41196, AMNH FARB 7224, AMNH FARB 7240, AMNH FARB 30638, MCZ VPRA 4327), the anterior end of the tooth row in the maxilla and adjacent posterior portion of the premaxilla curve upward to form a subnarial gap (Tykoski 1998). In MCZ VPRA 4326, the anterior end of the maxilla is horizontal, so the portion of the subnarial gap that would be formed by the maxilla is not present. An incipient subnarial gap is present on the somewhat more mature CM 31374, suggesting that a large subnarial gap visible in mature individuals develops later in ontogeny. The promaxillary foramen, an opening in the anterior end of the antorbital fossa in some early averostran-line theropods (Marsh and Rowe 2020) which evolves into a promaxillary fenestra in some later theropods (e.g., Godefroit et al. 2008), is not present in any C. bauri specimen studied here, or any coelophysoid for which maxillae are known.

Fig. 3. Immature skull of theropod dinosaur Coelophysis bauri (Cope, 1887), MCZ VPRA 4326 from the Rhaetian, Upper Triassic of Ghost Ranch, New Mexico, USA, in left lateral view; photograph (A1) and interpretive line drawing (A2). Damaged bone is indicated in medium gray, matrix is indicated in dark gray. Photo by SNM © President and Fellows of Harvard College. Abbreviations: an, angular; den, dentary; fr, frontal; hy, hyoid; ju, jugal; la, lacrimal; mx, maxilla; na, nasal; pa, parietal; qj, quadratojugal; qt, quadrate; sa; surangular; sclr, sclerotic ring; socc, supraoccipital.
Lacrimal and prefrontal.—The lacrimal (Figs. 2–4) is a thin bone composed of two processes: a vertical process that separates the orbit from the antorbital fenestra and a horizontal process that articulates with the frontal and the nasal. No suture is visible between the horizontal process of the lacrimal and the prefrontal. In CMNH 50957, the ventral end of the lacrimal is a triangular footplate partially obscured by matrix, but this is fully visible in MCZ VPRA 4326. In MCZ VPRA 4326, the middle of the vertical process is very thin (3.6 mm) but flares out into a subtriangular footplate (Fig. 4) with a width of 13.1 mm that articulates with the maxilla and jugal. In both specimens, the footplate is bordered by thin (<1 mm in CMNH 50957) ridges on the anterior and posterior sides. Each of these ridges flares out into a wider base towards the ventral end of the footplate. The lacrimal of the more mature skull CMNH 50955 is similar in overall morphology to MCZ VPRA 4326, though crucially its vertical process is much thicker: 7.9 mm at its narrowest point, with a less flared base of the footplate at 22.4 mm. In the much larger YPM VP 41196 (Fig. 5), the vertical process of the lacrimal is 9.9 mm at its narrowest point.
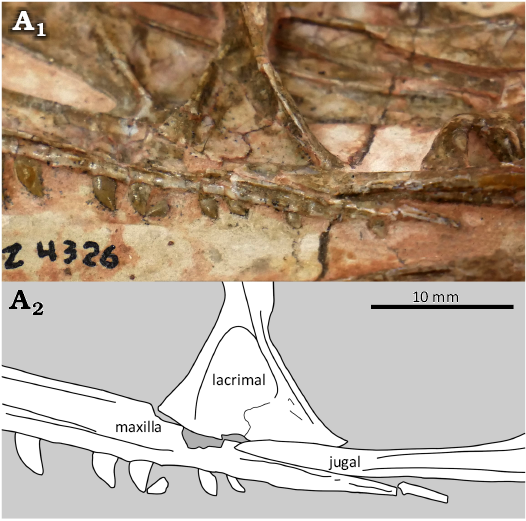
Fig. 4. The theropod dinosaur Coelophysis bauri (Cope, 1887), MCZ VPRA 4326 from the Rhaetian, Upper Triassic of Ghost Ranch, New Mexico, USA, closeup of articulation between maxilla, lacrimal, and jugal; photograph (A1) and interpretive line drawing (A2).

Fig. 5. The theropod dinosaur Coelophysis bauri (Cope, 1887), mature skull YPM VP 41196 from the Rhaetian, Upper Triassic of Ghost Ranch, New Mexico, USA.
Nasals and frontals.—No sutures are visible between the nasals and frontals in CMNH 50957. Based on comparisons with MCZ VPRA 4326 and other Coelophysis skulls, at least the posteriormost part of the nasals is present in CMNH 50957. Lateral crushing of the skull has separated the left and right nasals of MCZ VPRA 4326 from each other. The nasal is a thin bone (<1 mm dorsoventrally) that forms the anterior portion of the roof of the skull. It sits on top of the maxilla, and contacts the lacrimal. The nasal-frontal suture in MCZ VPRA 4326 is generally straight and approximately perpendicular to the anteroposterior axis of the skull. (Fig. 3) In comparison, this suture is curved in more skeletally mature C. bauri (e.g., MCZ VPRA 4327, also see Colbert 1989) and M. rhodesiensis (Raath 1977). Additionally, the skeletally mature holotype of the coelophysoid “Syntarsus” kayentakatae was reported to have posteriorly forked nasals (Tykoski 1998), so the morphology of this area is likely subject to both interspecific and ontogenetic variation.
Postorbital.—The right postorbital is preserved but badly damaged in CMNH 50957 (Fig. 2), making a detailed description impossible. This T-shaped bone composes the dorsal half of the border between the orbit and the lateral temporal fenestra. It comprises three processes: a ventral process which articulates with the dorsal tip of the jugal to form the posterior border of the orbit, a posterior process which is disarticulated and damaged in CMNH 50957, and an anterior process which articulates with the posteriormost edge of the frontal. The left postorbital of CMNH 50957 is also preserved but damaged, and it does not show the same lateral compression as the rest of the skull. No postorbital was confidently identifiable in MCZ VPRA 4326 as this area of the skull is badly damaged.
Sclerotic ring.—A partial sclerotic ring, consisting of four ossicles, is preserved in MCZ VPRA 4326 (Fig. 3). The ossicles are rectangular in shape and their length is approximately double their width. This is significantly different from the rounded trapezoid shape of the ossicles in NMMNH P-42200 described by Rinehart et al. (2009). NMMNH P-42200 was described as a “juvenile” by Rinehart et al. and is of comparable size to MCZ VPRA 4326. One possibility is that NMMNH P-42200 is more skeletally mature despite being of similar size. The antorbital fenestra of NMMNH P-42200 also has a parabolic profile similar to mature specimens (see Discussion) and it is at least plausible for these specimens to represent different ontogenetic stages given the high variability of growth trajectories in C. bauri (Barta et al. 2022). If this is the case, then it suggests an ontogenetic shift from a flat to globose eye during ontogeny. (See Rinehart et al. 2009 and citations therein for comparisons of sclerotic ring and eye types.)
Jugal.—The jugal (Fig. 6) is a thin bone with a long (30.3 mm in CMNH 50957 and 29.4 mm in MCZ VPRA 4326) horizontal bar connecting the quadratojugal and the maxilla and a shorter (12.5 mm in CMNH 50957 and 9.9 mm in MCZ VPRA 4326) vertical process that separates the orbit from the lateral temporal fenestra. In both specimens, the posterior end of the jugal is forked into two posterior processes, with the quadratojugal articulating with the jugal by having its anterior process fit into the slot formed by the forked jugal, as in a small, “juvenile” specimen of Megapnosaurus rhodesiensis (Bristowe and Raath 2004) and “subadult” holotype of Panguraptor lufengensis (You et al. 2014), but unlike the more mature C. bauri neotype (SNM personal observations) and M. rhodesiensis. Although Raath (1977) reconstructed NHMZ QG 278 as being posteriorly forked, the posterior process of this specimen is actually broken anterior to the reconstructed fork, and bears an articular facet on its ventral side only, as in the C. bauri neotype. Raath’s (1977: 37) description of the jugal as “forked at the posterior end” refers to the separate vertical and horizontal processes, not a fork in the posterior process itself. CM 31374, a C. bauri described as “subadult” (Bristowe and Raath 2004) shows a morphology intermediate between the immature condition where the quadratojugal articulates in a slot in the jugal and the mature condition where the quadratojugal articulates on the ventral side of the jugal, suggesting an intermediate ontogenetic stage. In later theropods such as tyrannosaurids (Carr 1999) and Majungasaurus (Ratsimbaholison et al. 2016), the posteriorly forked jugal is observed in individuals of various ontogenetic stages, so this feature cannot be used to reliably assess ontogeny in later theropods. Notably, the posteriorly forked jugal is also variable in the sauropodomorph Plateosaurus (Lallensack et al. 2021), though further work is needed to determine whether this variation is ontogenetic.
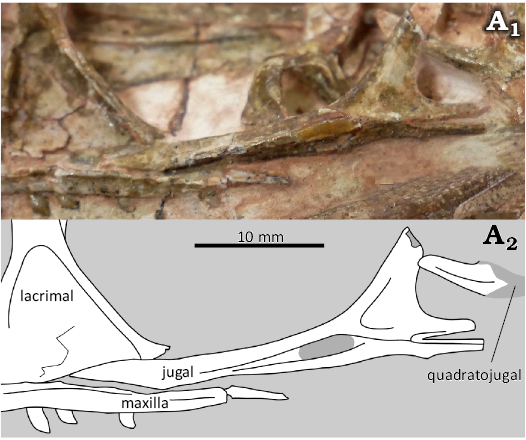
Fig. 6. The theropod dinosaur Coelophysis bauri (Cope, 1887), MCZ VPRA 4326 from the Rhaetian, Upper Triassic of Ghost Ranch, New Mexico, USA, closeup of jugal; photograph (A1) and interpretive line drawing (A2).
The jugal forms the ventral border and part of the posterior border of the orbit, and is excluded from the border of the antorbital fenestra by the lacrimal. A longitudinal ridge is present in both specimens, as identified in a small specimen of M. rhodesiensis (Bristowe and Raath 2004). Although the jugal of CMNH 50957 is disarticulated, in MCZ VPRA 4326 it is partially articulated with the maxilla and lacrimal, but not the quadratojugal. On the “juvenile” M. rhodesiensis figured by Bristowe and Raath (2004), there appears to be an indentation on the dorsal border of the horizontal process of the jugal near the anterior tip, which is not present on MCZ VPRA 4326. This area is damaged in CMNH 50957. However, it is not clear from Bristowe and Raath’s (2004) figures whether this indent represents a genuine anatomical feature or simply an area of damaged bone, and this feature is not mentioned in their text. The dorsal process of the jugal is broken on both Coelophysis specimens described here; in CMNH 50957 a fragment is broken off but still in close association, and in MCZ VPRA 4326 most of this process is missing.
Quadratojugal and quadrate.—The quadratojugal of MCZ VPRA 4326 (Fig. 3) is preserved but disarticulated. It is a small, L-shaped bone, closely resembling that of the large, presumably mature M. rhodesiensis specimen NHMZ QG 194 illustrated by Raath (1977). The quadrate is also preserved in MCZ VPRA 4326, but poorly so and partially obscured by the quadratojugal and matrix. The quadratojugal and quadrate of CMNH 50957 are both present but broken into several pieces and poorly preserved.
Dentary.—The dentary is a long, thin bone that forms the anterior portion of the lower jaw and contains the lower jaw teeth. It articulates posteriorly with the angular and surangular, and it forms the anterior and dorsal borders of the mandibular fenestra. As is typical of C. bauri specimens from the Ghost Ranch Coelophysis Quarry, the tight closure of the jaws in CMNH 50957 obscures most of the dentary (Fig. 2). Two dentary teeth are exposed in CMNH 50957.
MCZ VPRA 4326 is preserved with jaws open and part of its lower jaw is preserved (Fig. 3). Of the bones visible in lateral view, the angular is complete and well-preserved, while the posteriormost portion of the surangular and anterior half of the dentary are broken off. The portion of the dentary which is present is better preserved than most other skull bones, though like the others it is subject to taphonomic cracking and crushing. Six dentary teeth are present, five of which are in their alveoli.
Surangular and angular.—The surangular is a generally flat bone on the posterior side of the lower jaw. In CMNH 50957, a prominent longitudinal ridge extends for 14.2 mm along the surangular starting from its contact with the angular and flattening anteriorly (Fig. 2). It would articulate anteriorly with the dentary, but the dentary is disarticulated in this specimen. In MCZ VPRA 4326, the lower jaw is articulated. The mandibular fenestra is 16.3 mm long and 2.9 mm wide, and has the shape of a parallelogram (Fig. 3). The angular is an L-shaped bone that sits below the angular and forms the ventral and posterior borders of the mandibular fenestra, and part of the posteroventral border of the lower jaw. The morphology of the angular-dentary contact more closely resembles that of M. rhodesiensis as illustrated by Raath (1977) than it does the large C. bauri neotype AMNH FARB 7224. Likewise, the mandibular fenestra morphology of MCZ VPRA 4326 is somewhat similar to that illustrated by Raath (1977) in M. rhodesiensis and differs significantly from the elliptical mandibular fenestra of the C. bauri neotype.
Dentition.—The teeth of CMNH 50957 (Figs. 2, 7) are variously complete, broken, obscured by matrix, or missing. A total of 13 teeth are preserved on the right side of the skull. All teeth on the left side are obscured by matrix. All premaxillary teeth are missing. Of the 13 exposed teeth, all are in situ with crowns exposed and no roots visible. Four maxillary teeth and two dentary teeth are complete and exposed, with the rest either broken and incomplete or significantly obscured by matrix as listed in Table 1. Of the two maxillary teeth and two dentary teeth that have both mesial and distal edges exposed, all have visible serrations along the distal and mesial edges (Appendix 1). These serrations do not extend to the apex of the teeth in CMNH 50957, but in complete teeth of the larger individuals CMNH 50955 and CMNH 11882, the serrations extend to the apex of the tooth. There is no evidence of tooth replacement in this specimen, although new replacement teeth in Coelophysis would only be visible on the lingual side of the tooth row, which is not exposed in CMNH 50957.
In MCZ VPRA 4326, 20 teeth are preserved and visible, all on the left side of the skull (Fig. 3). No teeth on the right side are visible due to the plaster cradle of the specimen. As in CMNH 50957, all premaxillary teeth are missing. Serrations are not apparent on any of the teeth, though this is most likely due to damage during preparation, as some other specimens (e.g., AMNH FARB 7241, YPM VP 41196, CMNH 50955) have serrations present in some areas but visibly damaged by preparation in other areas (SNM and JEB personal observations). The Ghost Ranch material that was prepared at the AMNH in the 1950s, presumably including this specimen, was prepared without the aid of magnification, resulting in some damage (Downs 2000; see also Munyikwa and Raath 1999). MCZ VPRA 4326 preserves 14 maxillary teeth. As in CMNH 50957, there is no evidence of tooth replacement, but the lingual side of the tooth row is obscured. The condition of each maxillary tooth is listed in Table 1.
Table 1. Preservation status of maxillary teeth in juvenile Coelophysis bauri skulls MCZ VPRA 4326 and CMNH 50957. As the anterior portion of the maxilla is missing in CMNH 50957, tooth positions are estimated based on the assumption that both skulls have the same number of maxillary tooth alveoli and that there are no missing teeth posterior to the last tooth preserved in the maxillary tooth row of CMNH 50957. Mx, maxillary.
|
Tooth position |
Tooth condition |
|
|
MCZ VPRA 4326 |
CMNH 50957 |
|
|
Mx 1 |
complete, in situ |
missing |
|
Mx 2 |
missing |
missing |
|
Mx 3 |
complete, small portion |
missing |
|
Mx 4 |
complete, in situ |
missing |
|
Mx 5 |
missing |
missing |
|
Mx 6 |
complete, in situ |
missing |
|
Mx 7 |
root in situ, crown missing |
missing |
|
Mx 8 |
complete, small portion |
complete but partially obscured by matrix |
|
Mx 9 |
complete, in situ |
missing |
|
Mx 10 |
missing |
complete but partially obscured by matrix |
|
Mx 11 |
partially obscured by matrix |
missing |
|
Mx 12 |
complete, in situ |
complete but cracked mid-crown |
|
Mx 13 |
crown broken off but present |
broken |
|
Mx 14 |
complete, in situ |
complete, in situ |
|
Mx 15 |
complete, in situ |
complete, in situ |
|
Mx 16 |
missing |
broken near base; tip |
|
Mx 17 |
complete, in situ |
broken |
|
Mx 18 |
missing |
broken |
|
Mx 19 |
missing |
broken |
|
Mx 20 |
complete, in situ |
complete, in situ; |
Eleven maxillary teeth are present in CMNH 50957, with varying degrees of damage. The maxillary tooth row extends posteriorly to the posterior margin of the antorbital fenestra and anteriorly until the broken edge of the skull below the anterior margin of the antorbital fenestra, with a total length of 34.2 mm. The anteriormost maxillary tooth has serrations visible on the distal edge, but the mesial edge of the tooth is obscured by matrix so the presence or absence of serrations cannot be determined. According to Colbert’s (1989) description of the dentition, this would indicate that at least the first maxillary tooth is missing from the skull, as he observed that the first maxillary tooth of C. bauri has no serrations. However, some CMNH specimens show serrations on the first maxillary tooth, so this is probably a misinterpretation based on damage during preparation. The preservation status of serrations and number of serrations per millimeter of selected CMNH specimens is listed in Appendix 1. It is possible that the first preserved tooth of CMNH 50957 is the second maxillary tooth which according to Colbert (1989) only has posterior serrations, in which case this individual has a much lower number of teeth than the 18 expected based on Colbert’s description or the 20 present in MCZ VPRA 4326. The longest complete maxillary tooth of CMNH 50957 has a crown height of 4.3 mm while the shortest has a crown height of 1.1 mm.
Two dentary teeth are visible in right lateral view towards the anterior end of CMNH 50957. The tight closure of the jaws obscures most of the dentary teeth. The anterior tooth has a crown height of 3.4 mm and a crown base length of 1.7 mm, and the posterior tooth has a crown height of 4.3 mm and a crown base length of 2.0 mm, making it the largest tooth visible in these two skulls.
MCZ VPRA 4326 is preserved with open jaws (Fig. 2), but only the posterior part of the dentary is preserved, meaning the anterior portion of the tooth row is missing. Six teeth are present, most of which are in their alveoli.
The teeth of both specimens are all flat labiolingually and approximately triangular in lateral profile (Fig. 7): this is classified as ziphodont dentition (Hendrickx et al. 2015). There is a slight distal curvature near the apex of the teeth. The teeth of CMNH 50957 display slight ridges, which can be classified as flutes. (Hendrickx et al. 2015) These are not visible on MCZ VPRA 4326, most likely due to damage during mechanical preparation. The teeth appear lenticular to lanceolate in cross section, but the preservation and preparation of these skulls made cross sectional shape difficult to assess as they are only fully visible on the labial side. The triangular ziphodont dentition of these two immature specimens stands in contrast to the long, recurved teeth of large, mature C. bauri skulls (Fig. 7).

Fig. 7. Closeup of maxillary teeth of theropod dinosaur Coelophysis bauri (Cope, 1887) from the Rhaetian, Upper Triassic of Ghost Ranch, New Mexico, USA, showing detailed morphology. A. Immature specimen CMNH 50957. B. Robust mature specimen CMNH 50955.
Hyoids.—Both left and right hyoids are present in CMNH 50957 (Fig. 2). The preserved hyoid bones are the first ceratobranchials as identified by Colbert (1989). The right hyoid is one of the best-preserved bones in the skull, with minimal crushing and a total preserved length of 48.6 mm. The long, thin, curved shaft of the hyoid flares out into a rounded articular surface on the anterior end. The posterior end is broken and the exposed cross section of the shaft has a diameter of 1.2 mm along the dorsoventral axis. The left hyoid is visible in left lateral view of CMNH 50957, but is less complete than the right hyoid. Its posterior end is broken.
In MCZ VPRA 4326, three fragments of the left hyoid are preserved (Fig. 3), one close to the mandibular fenestra and two further fragments near the posterior end of the skull. Mangled among the posterior skull bones are four additional fragments which most likely represent parts of the left and right hyoids, identifiable by their rod-like morphology and flared ends.
Though theropod hyoids have been known for well over a century (Marsh 1884) and have been reported in several well-preserved specimens (e.g., Marsh 1884; Gilmore 1946; Rowe 1989; Russell and Dong 1993; Bristowe et al. 2004; Hu et al. 2018), they remain poorly described. They are universally described as thin and rod-like, and have little morphological diversity among non-paravian theropods, which largely retain plesiomorphic archosaurian hyoid morphology (Li et al. 2018). Unsurprisingly, the hyoids of immature Coelophysis are morphologically very similar to those reported in “Syntarsus” kayentakatae (Rowe 1989; Tykoski 1998) and Megapnosaurus rhodesiensis (Bristowe et al. 2004; Bristowe and Raath 2004), but notably the elements originally identified as hyoids by Raath (1977) are actually furculae (Tykoski et al. 2002). Intriguingly, hyoids are unusually common in the Coelophysis Quarry sample: at least six other skulls preserve partial hyoids (CMNH 50955, YPM VP 41196, MCZ VPRA 4327, MCZ VPRA 4333, AMNH FARB 7239, and AMNH FARB 7240) and two more (AMNH FARB 7242 and a disarticulated small skull also cataloged under the number MCZ VPRA 4327) preserve elements tentatively identified as hyoid fragments.
Discussion
Ontogeny.—CMNH 50957 and MCZ VPRA 4326 are of particular interest for ontogenetic study because they represent individuals partway through skeletal development. Coelophysis demonstrates high levels of intraspecific variation in growth and skeletal development, so size alone cannot be used as a reliable indicator of an individual’s skeletal maturity or ontogenetic age (Griffin and Nesbitt 2016; Griffin et al. 2021; Barta et al. 2022). Previous studies of growth and development in this species (Griffin and Nesbitt 2016; Barta et al. 2022) focused on postcranial elements such as limbs, so published data on cranial ontogeny is minimal. This description of immature Coelophysis bauri skulls aims to identify features that vary between mature and immature skulls of early theropods.
Most literature on theropod cranial ontogeny has focused on tyrannosaurids (e.g., Rozhdestvensky 1965; Russell 1970; Bakker et al. 1988; Carr 1999, 2010, 2020; Currie and Dong 2001; Currie 2003; Carr and Williamson 2004; Tsuihiji et al. 2011; Woodward et al. 2020; Voris et al. 2021) and other Late Cretaceous theropods (e.g., Bever and Norell 2009; Bhullar et al. 2012; Ratsimbaholison et al. 2016), but based on what is known for these more derived theropods, CMNH 50957 and MCZ VPRA 4326 are not unusual for skeletally immature individuals. These skulls are small and delicate compared to the more robust skulls of larger, more mature and presumably adult specimens such as CMNH 11882 (Fig. 8).

Fig. 8. The theropod dinosaur Coelophysis bauri (Cope, 1887), mature skull CMNH 11882 from the Rhaetian, Upper Triassic of Ghost Ranch, New Mexico, USA.
The high orbit length to skull length ratio of CMNH 50957 and MCZ VPRA 4326 is also expected in young dinosaurs (Coombs 1982; Chure 1998; Carr 1999; Salgado et al. 2005; Choiniere et al. 2013) and vertebrates more generally (Emerson and Bramble 1993). Although an accurate measurement of total skull length cannot be taken in either specimen due to damage to the anterior and posterior ends of the skulls, the orbit makes up at most 37% of the length of CMNH 50957. Meanwhile, CMNH 11882, a large and skeletally mature skull, is 196.9 mm long with a 33.2 mm orbit, so it has a ratio of 17%, closer to that of skeletally mature tyrannosaurids (Carr 1999).
The antorbital fenestra of MCZ VPRA 4326 has an almost rectangular profile on its anterior end, as in an immature Allosaurus sp. from Portugal (Rauhut and Fechner 2005) but unlike skeletally mature Coelophysis bauri (YPM VP 41196, AMNH FARB 7224, CMNH 50955) and Megapnosaurus rhodesiensis (NHMZ QG 194; Munyikwa and Raath 1999) where the anterior border of the antorbital fenestra resembles a parabola. Notably, the holotype skull of Panguraptor lufengensis, described as “subadult”, shows an intermediate condition (You et al. 2014). The anterior profile of the antorbital fenestra may therefore possibly be used as an indicator of skeletal maturity in some theropods, though this does not seem to be ontogenetically variable in tyrannosaurids (e.g., Carr 1999).
There are also notable differences in the dentition of CMNH 50957 and MCZ VPRA 4326 as compared to other Coelophysis skulls. Colbert’s (1989) description indicates variation in the number of teeth, but did not recognize significant morphological variation which is visible in the skulls examined for this study. The teeth of CMNH 50957 and MCZ VPRA 4326 are flatter in cross section and exhibit less curvature than larger specimens, giving them a roughly triangular shape in lateral view. This morphology can be classified as ziphodont, while the larger specimens display a more recurved conidont shape (sensu Hendrickx et al. 2015). The larger specimens also have a different ratio of crown base length to crown height, with larger specimens having proportionately taller teeth. On average, the crown base width of complete and fully erupted maxillary teeth in CMNH 50957 (n = 6) is 77% of the crown height. In the moderately sized CMNH 50958 (n = 14) and YPM VP 41196 (n = 8) , these ratios are 56% and 55% respectively, while in the very robust CMNH 50955 (n = 9), it is 67%. Further study of Coelophysis teeth with a larger dataset may confirm this trend of proportionately longer teeth in larger individuals, potentially indicating ontogenetic or individual variation in tooth shape and size. Two larger individuals, CMNH 50955 and CMNH 11882, also have higher density of serrations than CMNH 50957. Among the exposed teeth with mesial and/or distal carinae visible, teeth in the very robust CMNH 50955 have an average of 6.4 serrations per mm (n = 23), CMNH 11882 has an average of 9.2 serrations per mm (n = 16), and CMNH 50957 has an average of 10.2 serrations per mm (n = 9). This increase in serration size from smaller to larger individuals reflects an increase in the size of the teeth themselves.
The ontogenetic transition from ziphodonty to conidonty may suggest an ontogenetic shift in diet and/or feeding mechanism. Younger or less mature individuals may have used their ziphodont dentition to “deflesh” carcasses, cutting away small pieces of soft tissue to be swallowed, as is observed in the extant squamate Varanus komodoensis (D’Amore and Blumenschine 2009) while larger individuals would have been able to swallow proportionally larger amounts of tissue and bone, like Varanus salvator which uses its conidont teeth to grasp then swallow whole prey (Rieppel 1979). Our hypothesis is corroborated by the presence of a well-preserved crocodylomorph femoral head preserved as gut contents in a large, mature C. bauri (Nesbitt et al. 2006). The feeding mode described by Bakker (1986: 260) is probably limited to smaller, less mature individuals with ziphodont dentition. However, the analogy between Varanus spp. and Coelophysis is limited as Varanus exhibits extensive cranial kinesis (Rieppel 1979) that was probably absent in non-avialan dinosaurs (Holliday and Witmer 2008).
The first ceratobranchials of the hyoid are preserved in CMNH 50957, as well as CMNH 50955. Colbert (1989) also notes hyoids preserved in eight Coelophysis skulls, presumably including MCZ VPRA 4326 but not CMNH 50957 since the CMNH block had not been prepared at the time of Colbert’s description. Unlike other skull elements, histological study of the hyoid has recently been demonstrated to provide a record of growth in extant archosaurs (Griffin and Colleary 2021). Histological analysis of hyoids would be a much more reliable method of determining the age and maturity of an isolated skull than relying on size and morphology, due to Coelophysis having highly variable growth trajectories (Griffin and Nesbitt 2016; Barta et al. 2022). Preliminary data (JEB personal observations) from histology of cervical rib and hyoid samples from CMNH 50957 indicate this individual was immature, with one visible line of arrested growth in the cervical rib, two in the hyoid, and no external fundamental system. Histology from both elements supports the morphological assessment of CMNH 50957 as a young, skeletally immature individual. Given its similar morphology to CMNH 50957, MCZ VPRA 4326 is inferred to be immature.
Synonymy with Megapnosaurus.—CMNH 50957 and MCZ VPRA 4326 have some anatomy in common with NHMZ QG 165, a disarticulated “juvenile” skull of M. rhodesiensis described by Bristowe and Raath (2004). NHMZ QG 165 was used to dispute the validity of Megapnosaurus by Bristowe and Raath (2004), but was not compared to any Coelophysis bauri skulls of comparable ontogenetic age, since the smaller known skulls of C. bauri had only been briefly mentioned without a useful description in previously published literature (e.g., Colbert 1989). Bristowe and Raath (2004) instead compared NHMZ QG 165 to isolated skull elements and CM 31374, which they described as “subadult”, and indeed appears to be more skeletally mature than the skulls described in this study, but less mature than large C. bauri specimens such as CMNH 11882, AMNH FARB 7224, YPM VP 41196, and MCZ VPRA 4327.
The jugal, lacrimal, and tooth row were all considered relevant parts of the cranial anatomy for distinguishing Coelophysis from Megapnosaurus by Bristowe and Raath (2004). All of these elements are preserved in both CMNH 50957 and MCZ VPRA 4326. The jugal of CMNH 50957 and MCZ VPRA 4326 have a forked posterior process and a tapered anterior process, as in NHMZ QG 165, but unlike larger C. bauri skulls which have a less pronounced fork (CM 31374, CMNH 11882) or none at all (AMNH FARB 7224). The jugal is excluded from the border of the antorbital fenestra in CMNH 50957 and MCZ VPRA 4326, as in NHMZ QG 165 and mature individuals of both Coelophysis and Megapnosaurus (Bristowe and Raath 2004). This is also the case in Panguraptor lufengensis (You et al. 2014) and “Syntarus” kayentakatae (Rowe 1989) so this is most likely a coelophysoid synapomorphy. The lacrimal footplate possibly differs between Coelophysis and Megapnosaurus: in MCZ VPRA 4326 and CMNH 50955 the transition from the footplate to the bar is tapered and nearly symmetrical, whereas in NHMZ QG 165 it is asymmetrical and has a near-right angle on the anterior side, as in the holotype of Panguraptor lufengensis (You et al. 2014). Since this does not seem to be ontogenetically variable in C. bauri, it may be of taxonomic significance. The “crest” noted on the nasals of Megapnosaurus by Bristowe and Raath (2004) is not present on either specimen of Coelophysis. If it is indeed a genuine feature and not an artifact of taphonomy, then this also differs between the two genera.
Similarities between the two C. bauri skulls described here, NHMZ QG 165, and putatively mature skulls of M. rhodesiensis further support the great similarity between the two genera noted by various authors (Raath 1969, 1977; Paul 1988, 1993; Colbert 1989; Downs 2000; Bristowe and Raath 2004; McDavid and Bugos 2022). As Bristowe and Raath (2004) noted, individuals assigned to M. rhodesiensis are generally smaller than those assigned to C. bauri. Notably, most C. bauri individuals are skeletally immature (Barta et al. 2022); the same may be true of M. rhodesiensis. We hypothesize that some features considered to differentiate the two species may be ontogenetically variable and do not represent interspecific differences. Among these, the similar morphology of MCZ VPRA 4326 and putatively mature specimens of M. rhodesiensis in the mandibular fenestra, quadratojugal, and quadrate are particularly salient. Tykoski (2005) reached a similar conclusion for early theropods more generally, though he did not include C. bauri in his study and treated the larger specimens of M. rhodesiensis as mature. Based on anatomical differences that are consistent throughout ontogeny, we consider Megapnosaurus and Coelophysis separate genera, but caution that ontogenetically variable cranial features are not reliable for distinguishing taxa, and we encourage future workers to carefully evaluate traits that are potentially ontogenetically variable before using them to delineate taxa.
Acknowledgements
The authors would like to thank Chris Griffin (Princeton University, Princeton, New Jersey, USA) for his assistance with literature review, a helpful review of our manuscript, preliminary data on hyoid and cervical rib histology, and providing photographs of relevant specimens, Adam Marsh (US National Park Service, Petrified Forest, Arizona, USA) for a helpful review and providing relevant specimen photographs, Lee Hall (Museum of the Rockies, Bozeman, Montana, USA) for his help with literature review and starting this project, journal editor Daniel Barta (Acta Palaeontologica Polonica and Oklahoma State University College of Osteopathic Medicine at Cherokee Nation) for being accommodating of our long revision time, Darin Croft (Case Western Reserve University, Cleveland, Ohio, USA), Caitlin Colleary (CMNH), Scott Hartman (University of Wisconsin-Madison, USA), and Mickey Mortimer (Maple Valley, Washington, USA) for their advice and helpful conversations throughout this research, Amanda McGee (CMNH), Christina Byrd (MCZ), Carl Mehling (AMNH), Roger Benson (AMNH), Dan Brinkman (YPM), Vanessa Rhue (YPM), Bhart-Anjan Bhullar (YPM), and David Evans (ROM) for collections access, Justin Andell (Stockton University) for introducing the authors to each other, Hailey Majewski (CMNH) for surface-scanning comparative material, Robert Carr (CMNH) for photography, Bernhard Zipfel (Wits University, Johannesburg, South Africa) for useful information on South African specimens, SauriaZoicIllus for the life reconstruction on our SVP poster, Tyler Greenfield (Webster City, Iowa, USA), Lori Sinay (Altadena, California, USA), and the staff at the New York Public Library for providing copies of literature, and the late Dale Zelinski (CMNH) for his meticulous work preparing the CMNH Coelophysis block.
Author’s contributions
Study Conception and Design: JEB and SNM; Data Collection: JEB. and SNM; Analysis: SNM and JEB; Manuscript writing: SNM. and JEB; Illustration: SNM. Both authors reviewed and approved the final manuscript. An early version of this project was presented as a Case Western Reserve University Capstone project by JEB in 2021, and this work was presented in preliminary form as a poster by both authors at the 2023 Society of Vertebrate Paleontology Annual Meeting.
References
Bakker, R.T. 1986. The Dinosaur Heresies. 481 pp. William Morrow and Company, Inc., New York.
Bakker, R.T., Williams, M., and Currie, P. 1988. Nanotyrannus, a new genus of pygmy tyrannosaur, from the Latest Cretaceous of Montana. Hunteria 1 (5): 1–30.
Barta, D.E., Griffin, C.T., and Norell, M.A. 2022. Osteohistology of a Triassic dinosaur population reveals highly variable growth trajectories typified early dinosaur ontogeny. Scientific Reports 12 (1): article 17321. Crossref
Barta, D.E., Nesbitt, S.J., and Norell, M.A. 2018. The evolution of the manus of early theropod dinosaurs is characterized by high inter- and intraspecific variation. Journal of Anatomy 232: 80–104. Crossref
Bever, G.S. and Norell, M.A. 2009. The perinate skull of Byronosaurus (Troodontidae) with observations on the cranial ontogeny of paravian theropods. American Museum Novitates 3657: 1–52. Crossref
Bhullar, B.-A.S., Marugán-Lobón, J., Racimo, F., Bever, G.S., Rowe, T.B., Norell, M.A., and Abzhanov, A. 2012. Birds have paedomorphic dinosaur skulls. Nature 487: 223–226. Crossref
Bristowe, A. and Raath, M.A. 2004. A juvenile coelophysoid skull from the Early Jurassic of Zimbabwe, and the synonymy of Coelophysis and Syntarsus. Palaeontologia Africana 40: 31–41.
Bristowe, A., Parrott, A., Hack, J., Pencharz, M., and Raath, M. 2004. A non-destructive investigation of the skull of the small theropod dinosaur, Coelophysis rhodesiensis, using CT scans and rapid prototyping. Palaeontologia Africana 40: 159–163.
Carpenter, K. and Alf, K. 1994. Global distribution of dinosaur eggs, nests, and babies. In: K. Carpenter, K.F. Hirsch, and J.R. Horner (eds.), Dinosaur Eggs and Babies, 15–30. Cambridge University Press, Cambridge.
Carr, T.D. 1999. Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria). Journal of Vertebrate Paleontology 19: 497–520. Crossref
Carr, T.D. 2010. A taxonomic assessment of the type series of Albertosaurus sarcophagus and the identity of Tyrannosauridae (Dinosauria, Coelurosauria) in the Albertosaurus bonebed from the Horseshoe Canyon Formation (Campanian–Maastrichtian, Late Cretaceous). Canadian Journal of Earth Sciences 47: 1213–1226. Crossref
Carr, T.D. 2020. A high-resolution growth series of Tyrannosaurus rex obtained from multiple lines of evidence. PeerJ 8: e9192. Crossref
Carr, T.D. and Williamson, T.E. 2004. Diversity of late Maastrichtian Tyrannosauridae (Dinosauria: Theropoda) from western North America. Zoological Journal of the Linnean Society 142: 479–523. Crossref
Choiniere, J.N., Clark, J.M., Forster, C.A., Norell, M.A., Eberth, D.A., Erickson, G.M., Chu, H., and Xu, X. 2013. A juvenile specimen of a new coelurosaur (Dinosauria: Theropoda) from the Middle–Late Jurassic Shishugou Formation of Xinjiang, People’s Republic of China. Journal of Systematic Palaeontology 12: 177–215. Crossref
Chure, D.J. 1998. On the orbit of theropod dinosaurs. Gaia 15: 233–240.
Colbert, E.H. 1989. The Triassic dinosaur Coelophysis. Museum of Northern Arizona Bulletin 57: 1–160.
Colbert, E.H. 1990. Variation in Coelophysis bauri. In: K. Carpenter and P.J. Currie (eds.), Dinosaur Systematics: Perspectives and Approaches, 81–90. Cambridge University Press, Cambridge. Crossref
Coombs, W.P. 1982. Juvenile specimens of the ornithischian dinosaur Psittacosaurus. Palaeontology 25: 89–107.
Cope, E.D. 1887. The dinosaurian genus Coelurus. The American Naturalist 21: 367–369. Crossref
Cope, E.D. 1889. On a new genus of Triassic Dinosauria. The American Naturalist 23: 626. Crossref
Currie, P.J. 2003. Cranial anatomy of tyrannosaurid dinosaurs from the Late Cretaceous of Alberta, Canada. Acta Palaeontologica Polonica 48: 191–226.
Currie, P.J. and Dong, Z. 2001. New information on Shanshanosaurus huoyanshanensis, a juvenile tyrannosaurid (Theropoda, Dinosauria) from the Late Cretaceous of China. Canadian Journal of Earth Sciences 38: 1729–1737. Crossref
D’Amore, D.C. and Blumenschine, R.J. 2009. Komodo monitor (Varanus komodoensis) feeding behavior and dental function reflected through tooth marks on bone surfaces, and the application to ziphodont paleobiology. Paleobiology 35: 525–552. Crossref
Downs, A. 2000. Coelophysis bauri and Syntarus rhodesiensis compared, with comments on the preparation and preservation of fossils from the Ghost Ranch Coelophysis Quarry. New Mexico Museum of Natural History and Science Bulletin 17: 33–37.
Emerson, S.B. and Bramble, D.M. 1993. Scaling, Allometry, and Skull Design. In: J. Hanken and B.K. Hall (eds.), The Skull: Functional and Evolutionary Mechanisms. Vol. 3, 384–416. University of Chicago Press, Chicago.
Gilmore, C.W. 1946. New carnivorous dinosaur from the Lance formation of Montana. Smithsonian Miscellaneous Collections 106 (13): 1–19.
Godefroit, P., Currie, P.J., Hong, L., Yong, S.C., and Dong, Z.-M. 2008. A new species of Velociraptor (Dinosauria: Dromaeosauridae) from the Upper Cretaceous of northern China. Journal of Vertebrate Paleontology 28: 432–438. Crossref
Goldsmith, E.R., Nesbitt, S.J., and Stocker, M.R. 2023. Comparison of cranial and postcranial ontogeny in a size series of “Redondasaurus” (Archosauriformes: Phytosauria). In: D. Ehret and E. Fulwood (eds.), 83rd Annual Meeting Conference, Program Guide, 185–186. Society of Vertebrate Paleontology, Cinncinati.
Griffin, C. and Colleary, C. 2021. The osteohistology of hyoid elements preserves a record of growth in archosaurs. In: 81st Annual Virtual Meeting Conference Program, 128–129. Society of Vertebrate Paleontology.
Griffin, C.T. and Nesbitt, S.J. 2016. Anomalously high variation in postnatal development is ancestral for dinosaurs but lost in birds. Proceedings of the National Academy of Sciences 113: 14757–14762. Crossref
Griffin, C.T., Stocker, M.R., Colleary, C., Stefanic, C.M., Lessner, E.J., Riegler, M., Formoso, K., Koeller, K., and Nesbitt, S.J. 2021. Assessing ontogenetic maturity in extinct saurian reptiles. Biological Reviews 96 (2): 470–525. Crossref
Harris, J.D. and Downs, A. 2002. A drepanosaurid pectoral girdle from the Ghost Ranch (Whitaker) Coelophysis Quarry (Chinle Group, Rock Point Formation, Rhaetian), New Mexico. Journal of Vertebrate Paleontology 22: 70–75. Crossref
Heckert, A.B., Lucas, S.G., Rinehart, L.F., and Hunt, A.P. 2008. A new genus and species of sphenodontian from the Ghost Ranch Coelophysis Quarry (upper Triassic: Apachean), Rock Point Formation, New Mexico, USA. Palaeontology 51 (4), 827–845. Crossref
Hendrickx, C., Mateus, O., and Araújo, R. 2015. A proposed terminology of theropod teeth (Dinosauria, Saurischia). Journal of Vertebrate Paleontology 35 (5): e982797. Crossref
Holliday, C.M. and Witmer, L.M. 2008. Cranial kinesis in dinosaurs: intracranial joints, protractor muscles, and their significance for cranial evolution and function in diapsids. Journal of Vertebrate Paleontology 28: 1073–1088. Crossref
Hone, D.W.E., Farke, A.A., and Wedel, M.J. 2016. Ontogeny and the fossil record: What, if anything, is an adult dinosaur? Biology Letters 12 (2): 20150947. Crossref
Hu, D., Clarke, J.A., Eliason, C.M., Qiu, R., Li, Q., Shawkey, M.D., Zhao, C., D’Alba, L., Jiang, J., and Xu, X. 2018. A bony-crested Jurassic dinosaur with evidence of iridescent plumage highlights complexity in early paravian evolution. Nature Communications 9 (1): article 1. Crossref
Hunt, A.P. and Lucas, S.G. 1989. Late Triassic vertebrate localities in New Mexico. In: A.P. Hunt and S.G. Lucas (eds.), Dawn of the Age of Dinosaurs in the American Southwest, 72–101. New Mexico Museum of Natural History, Albuquerque.
Hunt, A.P. and Lucas, S.G. 1991. Rioarribasaurus, a new name for a Late Triassic dinosaur from New Mexico (USA). Paläontologische Zeitschrift 65: 191–198. Crossref
Hunt, A.P. and Lucas, S.G. 1993. A new phytosaur (Reptilia: Archosauria) genus from the Uppermost Triassic of the Western United States and its biochronological significance. New Mexico Museum of Natural History and Science Bulletin 3: 193–196.
Lallensack, J., Teschner, E., Pabst, B., and Sander, P.M. 2021. New skulls of the basal sauropodomorph Plateosaurus trossingensis from Frick, Switzerland: is there more than one species? Acta Palaeontologica Polonica 66: 1–28. Crossref
Li, Z., Zhou, Z., and Clarke, J.A. 2018. Convergent evolution of a mobile bony tongue in flighted dinosaurs and pterosaurs. PLOS ONE 13 (6): e0198078. Crossref
Lucas, S., Sullivan, R., Hunt, A., and Heckert, A. 2005. The saga of Coelophysis. In: S.G. Lucas, K.E. Zeigler, V.W. Lueth, and D.E. Owen (eds.), New Mexico Geological Society Field Conference Guidebook 56—Geology of the Chama Basin, 37–38. New Mexico Geological Society, Socorro.
Marsh, A.D. and Parker, W.G. 2020. New dinosauromorph specimens from Petrified Forest National Park and a global biostratigraphic review of Triassic dinosauromorph body fossils. PaleoBios 37: 1–56. Crossref
Marsh, A.D. and Rowe, T.B. 2020. A comprehensive anatomical and phylogenetic evaluation of Dilophosaurus wetherilli (Dinosauria, Theropoda) with descriptions of new specimens from the Kayenta Formation of northern Arizona. Journal of Paleontology 94 (S78): 1–103. Crossref
Marsh, O.C. 1884. Principal characters of American Jurassic dinosaurs: Part VIII, The order Theropoda. American Journal of Science s3-27 (160): 329–340. Crossref
McDavid, S.N. and Bugos, J.E. 2022. Taxonomic notes on Megapnosaurus and “Syntarsus” (Theropoda: Coelophysidae). The Mosasaur 12: 1–5.
Munyikwa, D. and Raath, M.A. 1999. Further material of the ceratosaurian dinosaur Syntarsus from the Elliot Formation (Early Jurassic) of South Africa. Palaeontologia Africana 35: 55–59.
Nesbitt, S.J. 2007. The anatomy of Effigia okeeffeae (Archosauria, Suchia), theropod-like convergence, and the distribution of related taxa. Bulletin of the American Museum of Natural History 302: 1–84. Crossref
Nesbitt, S.J. 2011. The early evolution of archosaurs: relationships and the origin of major clades. Bulletin of the American Museum of Natural History 352: 1–292. Crossref
Nesbitt, S.J. and Sues, H.-D. 2021. The osteology of the early-diverging dinosaur Daemonosaurus chauliodus (Archosauria: Dinosauria) from the Coelophysis Quarry (Triassic: Rhaetian) of New Mexico and its relationships to other early dinosaurs. Zoological Journal of the Linnean Society 191: 150–179. Crossref
Nesbitt, S.J., Irmis, R.B., and Parker, W.G. 2007. A critical re‐evaluation of the Late Triassic dinosaur taxa of North America. Journal of Systematic Palaeontology 5: 209–243. Crossref
Nesbitt, S.J., Turner, A.H., Erickson, G.M., and Norell, M.A. 2006. Prey choice and cannibalistic behaviour in the theropod Coelophysis. Biology Letters 2: 611–614. Crossref
Padian, K. 1986. On the type material of Coelophysis Cope (Saurischia: Theropoda) and a new specimen from the Petrified Forest of Arizona (Late Triassic: Chinle Formation). In: K. Padian (ed.), The Beginning of the Age of Dinosaurs, 45–60. Cambridge University Press, Cambridge.
Paul, G.S. 1988. Predatory Dinosaurs of the World: A Complete Illustrated Guide. 464 pp. Simon and Schuster, New York.
Paul, G.S. 1993. Are Syntarsus and the Whitaker quarry theropod the same genus? New Mexico Museum of Natural History and Science Bulletin 3: 397–402.
Pritchard, A.C. and Nesbitt, S.J. 2017. A bird-like skull in a Triassic diapsid reptile increases heterogeneity of the morphological and phylogenetic radiation of Diapsida. Royal Society Open Science 4 (10): 170499. Crossref
Raath, M.A. 1969. A new Coelurosaurian dinosaur from the Forest Sandstone of Rhodesia. Arnoldia (Rhodesia) 4 (28): 1–25.
Raath, M.A. 1977. The Anatomy of the Triassic Theropod Syntarsus rhodesiensis (Saurischia : Podokesauridae) and a Consideration of its Biology. 233 pp. Ph.D. Thesis, Rhodes University, Grahamstown.
Ratsimbaholison, N.O., Felice, R.N., and O’Connor, P. M. 2016. Ontogenetic changes in the craniomandibular skeleton of abelisaurid dinosaur Majungasaurus crenatissimus from the Late Cretaceous of Madagascar. Acta Palaeontologica Polonica 61: 281–292. Crossref
Rauhut, O.W.M. and Fechner, R. 2005. Early development of the facial region in a non-avian theropod dinosaur. Proceedings of the Royal Society B: Biological Sciences 272: 1179–1183. Crossref
Rieppel, O. 1979. A functional interpretation of the varanid dentition (Reptilia, Lacertilia, Varanidae). Gegenbaurs Morphologisches Jahrbuch 125: 797–817.
Rinehart, L.F., Lucas, S.G., Heckert, A.B., Spielmann, J.A., and Celesky, M.D. 2009. The paleobiology of Coelophysis bauri (Cope) from the Upper Triassic (Apachean) Whitaker quarry, New Mexico, with detailed analysis of a single quarry block. Bulletin of the New Mexico Museum of Natural History and Science 45: 1–269.
Rowe, T. 1989. A new species of the theropod dinosaur Syntarsus from the Early Jurassic Kayenta Formation of Arizona. Journal of Vertebrate Paleontology 9: 125–136. Crossref
Rozhdestvensky, A.K. 1965. Growth changes in Asian dinosaurs and some problems of their taxonomy (D.A. Russell, translation). [Original in Russian]. Paleontologičeskij žurnal 1965 (3): 95–109.
Russell, D.A. 1970. Tyrannosaurs from the Late Cretaceous of western Canada. Publications in Palaentology 1: 1–34.
Russell, D.A. and Dong, Z.-M. 1993. A nearly complete skeleton of a new troodontid dinosaur from the Early Cretaceous of the Ordos Basin, Inner Mongolia, People’s Republic of China. Canadian Journal of Earth Sciences 30 (10): 2163–2173. Crossref
Salgado, L., Coria, R.A., and Chiappe, L.M. 2005. Osteology of the sauropod embryos from the Upper Cretaceous of Patagonia. Acta Palaeontologica Polonica 50: 79–92.
Schaeffer, B. 1967. Late Triassic fishes from the western United States. Bulletin of the American Museum of Natural History 135: 287–342.
Schwartz, H.L. and Gillette, D.D. 1994. Geology and taphonomy of the Coelophysis Quarry, Upper Triassic Chinle Formation, Ghost Ranch, New Mexico. Journal of Paleontology 68: 1118–1130. Crossref
Smith, D.K. and Merrill, A. 2006. Facial variation in Coelophysis bauri and the status of Megapnosaurus (Syntarsus). New Mexico Museum of Natural History and Science Bulletin 37: 592–598.
Sues, H.-D., Nesbitt, S.J., Berman, D.S., and Henrici, A.C. 2011. A late-surviving basal theropod dinosaur from the latest Triassic of North America. Proceedings of the Royal Society B: Biological Sciences 278: 3459–3464. Crossref
Therrien, F. and Fastovsky, D.E. 2000. Paleoenvironments of early theropods, Chinle Formation (Late Triassic), Petrified Forest National Park, Arizona. Palaios 15: 194–211. Crossref
Tsuihiji, T., Watabe, M., Tsogtbaatar, K., Tsubamoto, T., Barsbold, R., Suzuki, S., Lee, A.H., Ridgely, R.C., Kawahara, Y., and Witmer, L.M. 2011. Cranial osteology of a juvenile specimen of Tarbosaurus bataar (Theropoda, Tyrannosauridae) from the Nemegt Formation (Upper Cretaceous) of Bugin Tsav, Mongolia. Journal of Vertebrate Paleontology 31: 497–517. Crossref
Tykoski, R.S. 1998. The Osteology of Syntarsus kayentakatae and Its Implications for Ceratosaurid Phylogeny. 217 pp. Ms. Thesis, University of Texas at Austin, Austin.
Tykoski, R.S. 2005. Anatomy, Ontogeny, and Phylogeny of Coelophysoid Theropods. 572 pp. Ph.D. Thesis, University of Texas at Austin, Austin.
Tykoski, R.S., Forster, C.A., Rowe, T., Sampson, S.D., and Munyikwa, D. (2002). A furcula in the coelophysid theropod Syntarsus. Journal of Vertebrate Paleontology 22: 728–733. Crossref
Voris, J.T., Zelenitsky, D.K., Therrien, F., Ridgely, R.C., Currie, P.J., and Witmer, L.M. 2021. Two exceptionally preserved juvenile specimens of Gorgosaurus libratus (Tyrannosauridae, Albertosaurinae) provide new insight into the timing of ontogenetic changes in tyrannosaurids. Journal of Vertebrate Paleontology 41 (6): e2041651. Crossref
Whiteside, J.H., Lindström, S., Irmis, R.B., Glasspool, I.J., Schaller, M.F., Dunlavey, M., Nesbitt, S.J., Smith, N.D., and Turner, A.H. 2015. Extreme ecosystem instability suppressed tropical dinosaur dominance for 30 million years. Proceedings of the National Academy of Sciences 112: 7909–7913. Crossref
Woodward, H.N., Tremaine, K., Williams, S.A., Zanno, L.E., Horner, J.R., and Myhrvold, N. 2020. Growing up Tyrannosaurus rex: osteohistology refutes the pygmy “Nanotyrannus” and supports ontogenetic niche partitioning in juvenile Tyrannosaurus. Science Advances 6 (1): eaax6250. Crossref
You, H.-L., Azuma, Y., Wang, T., Wang, Y.-M., and Dong, Z.-M. 2014. The first well-preserved coelophysoid theropod dinosaur from Asia. Zootaxa 3873 (3): article 3. Crossref
Zeigler, K.E., Kelley, S., and Geissman, J.W. 2008. Revisions to stratigraphic nomenclature of the Upper Triassic Chinle Group in New Mexico: new insights from geologic mapping, sedimentology, and magnetostratigraphic/paleomagnetic data. Rocky Mountain Geology 43 (2): 121–141. Crossref
Acta Palaeontol. Pol. 69 (4): 549–563, 2024
https://doi.org/10.4202/app.01085.2023
Serration status and serrations per mm of teeth in selected CMNH Coelophysis bauri skulls. The lengths of teeth measured (mm) represent the longest uninterrupted row of serrations on each tooth surface/carina, and are often shorter than the entire length of the erupted tooth. * tooth count from mesial to distal, does not represent tooth position and does not include empty alveoli.
|
Tooth number* |
CMNH 50957 right |
CMNH 50955 left |
CMNH 50955 right |
CMNH 11882 right |
||||
|
distal carina |
mesial carina |
distal carina |
mesial carina |
distal carina |
mesial carina |
distal carina |
mesial carina |
|
|
(serrations/mm) |
||||||||
|
Maxilla |
||||||||
|
1 |
20/1.77 mm |
6/0.88 mm |
14/1.54 mm |
8/1.26 mm |
|
none visible |
14/1.54 mm |
8/1.26 mm |
|
2 |
15/1.51 mm |
|
|
22/2.38 mm |
|
|
|
22/2.38 mm |
|
3 |
14/1.23 mm |
4/0.40 mm |
17/2.70 mm |
|
|
|
17/2.70 mm |
|
|
4 |
|
|
18/2.07 mm |
|
|
22/2.98 mm |
18/2.07 mm |
|
|
5 |
16/1.4 mm |
7/0.77 mm |
11/1.97 mm |
12/1.89 mm |
17/2.77 mm |
|
11/1.97 mm |
12/1.89 mm |
|
6 |
8/0.91 mm |
|
|
|
18/3.27 mm |
28/4.34 mm |
|
|
|
7 |
8/0.78 mm |
|
14/2.46 mm |
22/3.91 mm |
20/3.90 mm |
28/4.73 mm |
14/2.46 mm |
22/3.91 mm |
|
8 |
|
|
|
|
|
23/3.57 mm |
|
|
|
9 |
|
|
42/8.01 mm |
20/3.01 mm |
|
|
42/8.01 mm |
20/3.01 mm |
|
10 |
|
|
|
|
|
9/1.70 mm |
|
|
|
11 |
9/0.78 mm |
|
36/6.96 mm |
25/4.32 mm |
|
|
36/6.96 mm |
25/4.32 mm |
|
12 |
|
|
|
8/1.41 mm |
|
50/8.03 mm |
|
8/1.41 mm |
|
13 |
|
|
|
21/3.41 mm |
|
17/3.55 mm |
|
21/3.41 mm |
|
14 |
|
|
|
|
|
|
|
|
|
15 |
|
|
|
|
11/1.91 mm |
|
|
|
|
16 |
|
|
12/1.78 mm |
9/1.19 mm |
17/2.67 mm |
|
12/1.78 mm |
9/1.19 mm |
|
17 |
|
|
5/0.78 mm |
10/1.23 mm |
|
|
5/0.78 mm |
10/1.23 mm |
|
18 |
|
|
19/2.63 mm |
|
|
|
19/2.63 mm |
|
|
19 |
|
|
|
|
|
|
|
|
|
20 |
|
|
|
|
|
|
|
|
|
Total count maxilla |
n = 7 |
n = 3 |
n = 10 |
n = 10 |
n = 5 |
n = 7 |
n = 10 |
n = 10 |
|
Average serration/mm maxilla teeth |
10.7 |
8.6 |
6.6 |
6.8 |
5.8 |
6.1 |
7.7 |
8.1 |
|
Dentary |
||||||||
|
1 |
17/1.54 mm |
13/1.11 mm |
no serrations visible |
no exposed right dentary teeth in CMNH 50955 |
|
|
||
|
2 |
|
11/1.22 mm |
|
|
||||
|
3 |
|
|
5/0.53 mm |
|
||||
|
4 |
|
|
|
|
||||
|
5 |
|
|
24/2.60 mm |
|
||||
|
6 |
|
|
|
|
||||
|
7 |
|
|
|
12/0.89 mm |
||||
|
8 |
|
|
|
|
||||
|
Total count dentary |
n = 1 |
n = 2 |
n = 0 |
n = 0 |
n = 0 |
n = 0 |
n = 2 |
n = 1 |
|
Average serration/mm dentary teeth |
11.0 |
10.4 |
|
|
|
|
9.2 |
13.5 |
|
Average serrations per mm |
10.2 |
6.4 |
9.2 |
|||||
|
Total teeth in sample |
n = 9 |
n = 23 |
n = 16 |
|||||