

Pterosaur teeth from the Southern Neuquén Basin (Patagonia, Argentina): New insights on the reconstruction of ornithocheiriform dental anatomy
ALESSIO CIAFFI and FLAVIO BELLARDINI
Ciaffi, A. and Bellardini, F. 2024. Pterosaur teeth from the Southern Neuquén Basin (Patagonia, Argentina): New insights on the reconstruction of ornithocheiriform dental anatomy. Acta Palaeontologica Polonica 69 (1): 73–86.
The pterosaur fossil record of Argentina is increasing in recent times, both in the number of localities and their temporal range. The new materials are found in levels that span from the Late Triassic to the Late Cretaceous in age. In this contribution we describe twenty isolated pterosaur teeth from the Cerro de los Leones locality (Picún Leufú, Neuquén Province), where the Lohan Cura Formation (Albian) widely crops out. A new, detailed analysis of these remains allowed us to detect a set of morphological features, such as the enamel texture, cross-section shape, crown curvature, and absence of sharp carinae, which suggests close affinities with Ornithocheiriformes. Moreover, with the large number of tooth specimens, a discrimination of three morphotypes based on the crown curvature, the basal cross-section shape, and the presence of carinae was carried out. We performed extensive morphological comparisons between different members of Ornithocheiriformes with the aim to test the hypothesis that the presence of these different morphotypes reflects distinct anatomical positions of the teeth in the jaws . Thus, considering the heterodont dentition of ornithocheiriforms, the tooth morphological classification proposed here represents a new methodology, not only to identify isolated ornithocheiriform teeth, but also to assign them to the rostral, medial, or the posterior portion of the tooth row. Finally, the evidence of ornithocheiriform pterosaurs in Cerro de los Leones has paleobiogeographical implications, allowing us to correlate it with other Albian faunas from South America, expanding our knowledge of the distribution of Early Cretaceous pterosaurs across southwestern Gondwana.
Key words: Anhangueridae, Cretaceous, Gondwana, Neuquén Basin, tooth morphology, enamel, heterodonty.
A. Ciaffi [al.ciaffi@gmail.com], Circonvallazione Ostiense, Italy.
F. Bellardini [flaviobellardini@gmail.com; ORCID: https://orcid.org/0000-0003-1616-8548 ], 2 CONICET (Consejo Nacional de Investigaciones Científicas y Técnicas); Instituto de Investigación en Paleobiología y Geología (IIPG). Av. Roca 1242, R8332EXZ, General Roca, Río Negro, Argentina; Museo Carlos Ameghino (MPCA), Belgrano 1700, Paraje Pichi Ruca (predio Marabunta), Cipolletti R8324, Río Negro, Argentina; and Universidad Nacional de Río Negro (UNRN), Isidro Lobo 516, (8332) General Roca, Río Negro, Argentina.
Received 11 November 2023, accepted 21 February 2024, published online 22 March 2024.
Copyright © 2024 A. Ciaffi and F. Bellardini. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Pterosaurs were the first vertebrates to evolve powered flight and their fossil remains are found on every continent (Barrett et al. 2008; Kellner et al. 2019; Reguero et al. 2022), from the Upper Triassic to the Upper Cretaceous (e.g., Long and Murry 1995; Dalla Vecchia 2003; Upchurch et al. 2015; Martínez et al. 2022). Among the different adaptations to the flying lifestyle of this group, an extended invasive pneumatic system (Claessens et al. 2009) and hollow bones with extremely thin cortices (Elgin and Hone 2013) make pterosaur skeletal material notoriously fragile and difficult to preserve. This explains their scarcity in the fossil record, when compared with other taxa (Kellner 1994). In Gondwana, the majority of the outcrops that yielded pterosaur material is Cretaceous in age (Martínez et al. 2015). However, the study of Gondwanan pterosaur diversity is subject to sampling biases and is further challenged by complex historical and socioeconomic aspects of some localities of the Southern Hemisphere (Raja et al. 2021). In fact, despite being well represented in the Northern hemisphere, with most of the findings being from Europe (Barrett et al. 2008), Asia (Wang et al. 2005), and North America (Padian 1984; Kellner and Langston 1996), the Gondwanan Early Cretaceous pterosaur fossil record is comparatively limited. Indeed, the pterosaur evidence from Africa (Knoll 2000; Blackburn 2002) and Australia (Molnar and Thulborn 1980, 2007; Pentland and Poropat 2023) is scarce and fragmentary. Nevertheless, South America yielded a great variety of pterosaur materials (Codorniú and Gianechini 2016; Alarcón-Muñoz et al. 2020, 2022), exemplified by the abundant and diversified record from the Romualdo Formation and the Crato Formation (Lower Cretaceous) of northeastern Brazil (Price 1971; Kellner et al. 2013). In central Argentina, hundreds of specimens belonging to a single species, the filter-feeding Pterodaustro guinazui (Codorniú and Chiappe 2004; Codorniú et al. 2013, 2018), have been collected in the San Luis Province (Aptian). In Patagonia, the Early Cretaceous pterosaur record is represented by a proximal portion of a femur from La Amarga Formation (Hauterivian–Aptian; Montanelli 1987), a fragmentary radius and ulna from the Río Belgrano Formation (Barremian; Kellner et al. 2003; Codorniú and Gianechini 2016) and a proximal portion of an ulna from the Lohan Cura Formation (Albian; Bellardini and Codorniú 2019). Moreover, Martinelli et al. (2007) tentatively assigned to Pterosauria two isolated teeth found in the Cerro de los Leones locality (Lohan Cura Formation), suggesting similarities to the Brazilian pterosaur from genus Anhanguera. Because of its rarity in the fossil record, even fragmentary and isolated pterosaur material is relevant to reconstruct the evolutionary history of this morphologically diversified and geographically dispersed group (Kellner et al. 2003). The occurrence of the pterodactyloid clade Ornithocheiriformes (Andres 2021) is well known from Lower Cretaceous deposits, but most of the evidence consists of isolated teeth from the marine, estuarine and lacustrine outcrops of Brazil (Elias et al. 2007; Rodrigues and Kellner 2010), France (Vullo and Neraudeau 2009), Morocco (Kellner and Mader 1997; Wellnhofer and Buffetaut 1999; Knoll 2000), New South Wales (Brougham et al. 2017), Russia (Averianov 2007; Solonin et al. 2021), Spain (Sánchez-Hernández et al. 2007; Vullo et al. 2009), Tunisia (Martill et al. 2018), and Ukraine (Sokolskyi 2023). These records provide important data to reconstruct the paleobiogeographical distribution of ornithocheiriform pterosaurs and their morphological and taxonomic diversification. However, to date, the anatomical and taxonomic attribution of isolated ornithocheiriform teeth is a challenging task, lacking a specific approach to discriminate not only the interspecific, but also the intraspecific dental diversification, especially taking into account their heterodont condition (Frey et al. 2003). The aim of this contribution is to describe eighteen isolated teeth from the Lohan Cura Formation (Albian) outcrop at the Cerro de los Leones locality (Picún Leufú, Neuquén Province), as well as redescribe two isolated tooth specimens reported by Martinelli et al. (2007). Considering the poor record of fossil teeth from Argentina, the description of this evidence provides new data on the pterosaur faunal composition of the southern Neuquén Basin during the Albian, improving our knowledge on the paleoecology and the paleobiogeography of Early Cretaceous pterosaurs from southwestern Gondwana. Finally, a new anatomical and taxonomic approach to identify isolated ornithocheiriform teeth is proposed.
Institutional abbreviations.—AODF, Age of Dinosaurs Fossil, Winton, Queensland, Australia; FSAC, Faculté des Sciences Ain Chock, Université Hassan II, Casablanca, Morocco; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China; LINHM, Long Island Natural History Museum, New York, USA; LRF, Australian Opal Centre, Lightning Ridge, Australia; MA, “Musée d’Angoulême”, Angoulême, France; FDB, “Font-de-Benon”, FVA, “Fouras-Vauban”, RND, “Les Renardières” collections; MCF-PVPH, Museo Municipal “Carmen Funes”, Plaza Huincul, Argentina; MHNS, Museum of Natural History Sintra, Sintra, Portugal; MN, Museu Nacional/Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil; NNPM, National Natural History Museum of National Academy of Sciences of Ukraine, Kyiv, Ukraine; NSM, Division of Vertebrate Paleontology, National Science Museum, Tokyo, Japan; QM, Queensland Museum, Brisbane, Australia; RH, Mémoire de la terre, Museum Tataouine, C207 Tataouine, Tunisia; RSU GE, Ryazan State University, Department of Geography, Ecology and Nature Management (DGENM), Ryazan, Russian Federation; SMNK PAL, Staatliches Museum für Naturkunde Karlsruhe, Germany; UFMA, Universidade Federal do Maranhão, São Luís, Brazil; URC, Universidade Estadual Paulista, Rio Claro, Brazil; ZIN PH, Paleoherpetological collection of the Zoological Institute of the Russian Academy of Sciences, St. Petersburg, Russian Federation.
Geological setting
The specimens described here were collected during different fieldworks carried out from 1997 to 2016 by the Museo Argentino de Ciencias Naturales “Bernardino Rivadavia” of Buenos Aires (Martinelli et al. 2022) and the Museo Municipal “Carmen Funes” of Plaza Huincul (Coria 2022) in the Cerro de los Leones locality, 11 km southwest of the city of Picún Leufú (Fig. 1).
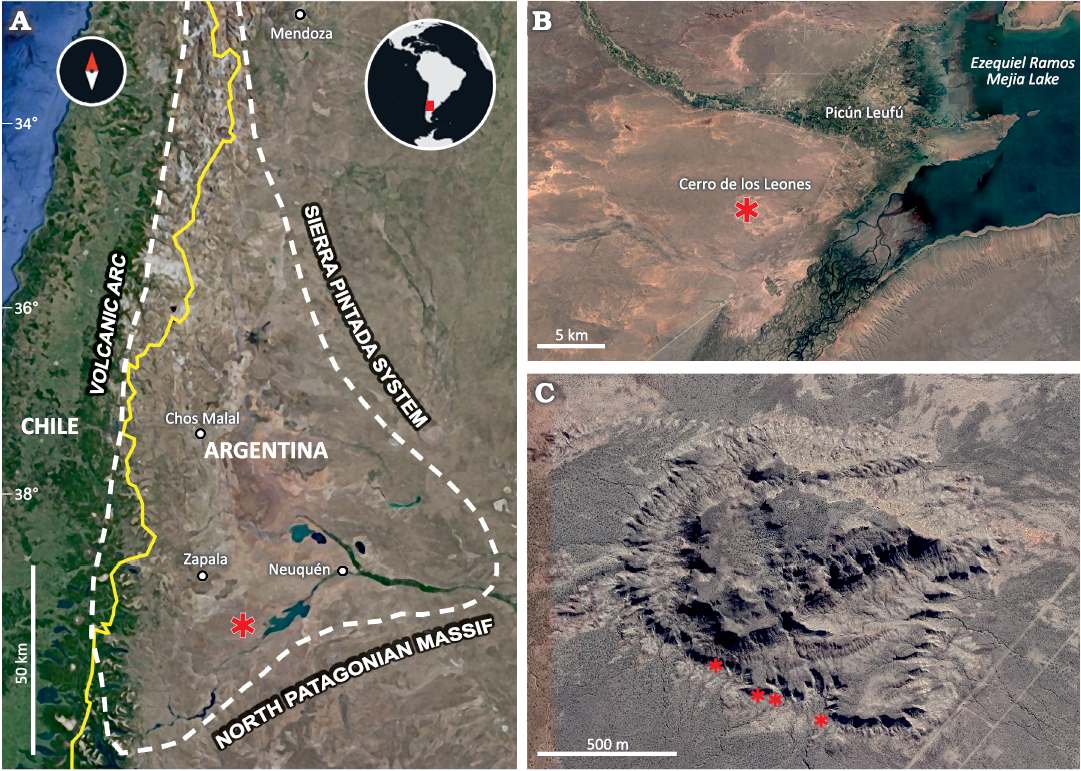
Fig. 1. Geographic position of Cerro de los Leones in the Neuquén Basin. A. Map of the Neuquén Basin, showing position of the studied area (asterisk); white dashed line indicates the boundary of the Neuquén Basin; yellow line indicates the boundary between Chile and Argentina. Satellite views of the Cerro de los Leones area showing relation to Picún Leufú (B) and locations (asterisks) of provenance of vertebrate fossils (images from Google Earth).
This area contains outcrops of a thick sequence of fluvial sediments referred to the upper section of the Lohan Cura Formation, the Cullín Grande Member (Albian; Leanza and Hugo 1997, 2011; Leanza 2003), which is composed principally of mudstones with intercalations of thin layers of sandstones and evaporitic episodes characterized by pelite-gypsum-limestone cycles, suggesting a lagoonal depositional environment evolved under alternating dry/humid climatic conditions for this sequence (Martinelli et al. 2007). Moreover, following Martinelli et al. (2007), four main fossiliferous layers at the Cerro de los Leones were detected, of which the first three have yielded most of the fossil specimens recorded from this area, including sauropod and theropod dinosaurs, pterosaurs, turtles, crocodiles, fresh-water mollusks, gastropods, fish, and plants (Bonaparte 1999; Bonaparte et al. 2006; Martinelli et al. 2007; Bellardini and Cerda 2017; Bellardini and Codorniú 2019; Bellardini et al. 2022). The sedimentological analysis suggests a distal floodplain depositional environment for these layers, which are characterized by paleosol, pond, swamp, and sheet flood deposits (Martinelli et al. 2007). The teeth described here were found isolated and scattered in proximity of Fossiliferous Layer N°1 (sensu Martinelli et al. 2007; Fig. 2).
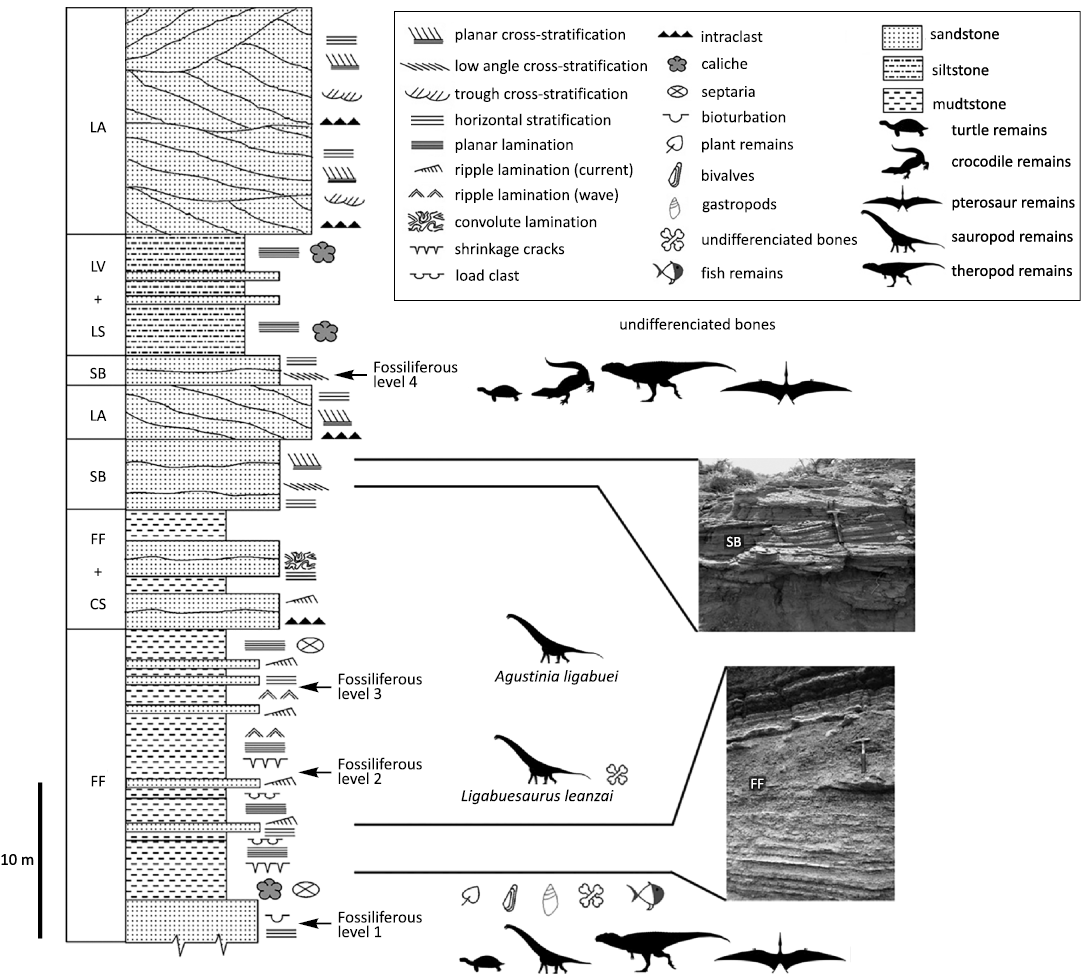
Fig. 2. Stratigraphic column of the Cullín Grande Member (Lohan Cura Formation), Albian, Lower Cretaceous, Cerro de Los Leones, Patagonia, Argentina. Architectural element codes follow Miall 1996: CS, crevasse channel; FF, floodplain fines; LA, lateral accretion; LS, laminated sand sheets; LV, levee; SB, sandy bedforms. Modified from Martinelli et al. 2007 with silhouettes from www.phylopic.org: pterosaur (Anhanguera santanae) silhouette credit of Leon P.A.M. Claessens, Patrick M. O’Connor, David M. Unwin; sauropod (Dreadnoughtus schrani) silhouette by Scott Hartman; theropod (Giganotosaurus carolinii) silhouette by Tasman Dixon; crocodile silhouette by B. Kimmel. Anhanguera santanae and Dreadnoughtus schani silhouettes license: https://creativecommons.org/licenses/by-sa/3.0/.
Material and methods
The material described in this contribution consists of twenty isolated teeth housed in the Museo Municipal “Carmen Funes” of Plaza Huincul, Neuquén Province, Argentina. The general dental description follows the terminology proposed by Hendrickx et al. (2015) for theropod dinosaur specimens. Considering that ornithocheiriform tooth morphology does not strictly follow dinosaur dentition, the term “carina” is here referred to the apicobasal constriction of the mesial and distal crown surfaces that does not correspond exactly to the sharp and well delimited ridge described by Hendrickx et al. (2015). In this context, some narrow but soft apicobasal edges of the specimens have not been considered carinae, in order to distinguish them from more relevant characters for morphological description and comparison. To date, the phylogenetic assessment of Pterosauria is widely debated (Kellner 2003; Lü et al. 2009; Unwin 2003); despite this, the focus of this contribution is not to explore the phylogenetic relationships between pterosaur clades, and we follow the systematic proposal by Andres (2021). The fossil material was observed using a LEICA MZ6 microscope and measured using a Black caliper. Photographs of the specimens were taken using the 5 mpx Macro digital camera of a Samsung Galaxy A31 smartphone. Specimen tables of the teeth have been drawn with a Wacom Intuos Pro drawing tablet and using the software Adobe Photoshop CC 2015.
Systematic paleontology
Pterosauria Kaup, 1834
Pterodactyloidea Plieninger, 1901
Lanceodontia Andres et al., 2014
Ornithocheiriformes Andres, 2021
Figs. 3–6.
Material.—Twenty isolated teeth: MCF-PVPH-879-4, 879-6, 879-8, 879-10 (Fig. 4); MCF-PVPH-739-2, 743, 879-9, 880-1, 880-2, 880-3, and 880-4 (Fig. 5); MCF-PVPH-741, 875, 879-1, 879-2, 879-3, 879-5, 879-7, 879-11, and 880-5 (Fig. 6). All from the Cretaceous Cerro de los Leones locality, Albian, Lower Cretaceous, Picún Leufú, Argentina.
Description.—All specimens here described are represented by isolated tooth crowns with reabsorbed or lacking roots (Fig. 3). In this sense, the apical wear facets, when present, and the lack of the roots do not seem to have been caused by weathering events and/or taphonomic processes, suggesting that the teeth could have been lost in life. The tooth-crowns are conidont (cone-shaped, sensu Hendrickx et al. 2015), apicobasally elongated and slightly recurved. The cross-sections are diversified in shape, subcircular, oval or elliptical. Taking into account the general morphology of the teeth, we identify three morphotypes, which are defined on the basis of three morphological features (Table 1): tooth crown curvature, shape of the crown cross-section, and presence of carinae.

Fig. 3. Isolated pterosaur teeth from the Cerro de los Leones locality, Albian, Lower Cretaceous, Picún Leufú, Argentina. Morphotype 1 (H, J, L, N), Morphotype 2 (A, C, M, P, Q, R, S), and Morphotype 3 (B, D, E, G, I, K, O, T). A. MCF-PVPH-739-2. B. MCF-PVPH-741. C. MCF-PVPH-743. D. MCFPVPH-875. E. MCF-PVPH-879-1. F. MCF-PVPH-879-2. G. MCF-PVPH-879-3. H. MCF-PVPH-879-4. I. MCF-PVPH-879-5. J. MCF-PVPH-879-6. K. MCF-PVPH-879-7. L. MCF-PVPH-879-8. M. MCF-PVPH-879-9. N. MCF-PVPH-879-10. O. MCF-PVPH-879-11. P. MCF-PVPH-880-1. Q. MCFPVPH-880-2. R. MCF-PVPH-880-3. S. MCF-PVPH-880-4. T. MCF-PVPH-880-5. In labial (A–G, I, K, O–Q, S, T), lingual (H, M, N, R), and mesial (J, L) views.
Table 1. Most inclusive morphological characterization of the Cerro de Los Leones teeth, divided in three morphotypes.
| |
Crown curvature |
Cross-section shape |
Carinae |
Teeth |
|
Morphotype 1 |
more lingual than distal |
subcircular |
absent |
MCF-PVPH-879-4, 879-6, 879-8, 879-10 |
|
Morphotype 2 |
more lingual than distal |
elliptical |
present |
MCF-PVPH-739-2, 743, 879-9, 880-1, 880-2, 880-3, 880-4 |
|
Morphotype 3 |
more distal than lingual |
oval |
absent |
MCF-PVPH-741, 875, 879-1, 879-2, 879-3, 879-5, 978-7, 879-11, 880-5 |
Morphotype 1.—MCF-PVPH-879-4, 879-6, 879-8, 879-10 (Fig. 4). These tooth crowns are apicobasally elongated and strongly lingually recurved. The basal cross-section is subcircular, and weakly labiolingually compressed. All specimens lack carinae. In MCF-PVPH-879-4, 879-8, and 879-10 the enamel layer is fragmentary, exposing the underlying dentine layer. The dentine surface is mainly smooth, but several transversal and longitudinal fractures are present, especially on the lingual and apical surfaces. The preserved enamel surface is slightly wrinkled, showing a set of anastomosed, apicobasally extended ridges and grooves, which are more pronounced basally and progressively weaker apically. On the other hand, MCF-PVPH-879-6 preserves small fragments of enamel on the labial and lingual surfaces, resulting in an almost smooth surface. The specimens where the apical portion is preserved (MCF-PVPH-879-6 and 879-8) show wear facets in apical, labial or mesial position. In MCF-PVPH-879-8 the apical wear facet is wide and rounded, extending basally to the half of the crown (Fig. 4).
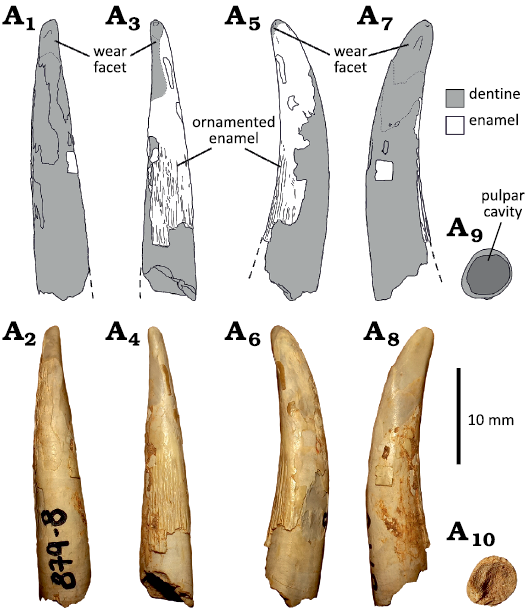
Fig. 4. Isolated tooth (Morphotype 1) from the Cerro de los Leones locality, Albian, Lower Cretaceous, Picún Leufú, Argentina. MCF-PVPH-879-8 in labial (A1, A2), lingual (A3, A4), mesial (A5, A6), and distal (A7, A8) views. The basal cross section (A9, A10) is visible due to the natural breaking of the crown.
Morphotype 2.—MCF-PVPH-739-2, 743, 879-9, 880-1, 880-2, 880-3, and 880-4 (Fig. 5). These tooth crowns are slightly lingually recurved and almost straight in mesial view. The basal cross-section is elliptical, with a labiolingual compression. The dentine surface is almost smooth. MCF-PVPH-739-2 is incomplete, lacking part of the basal and apical portion, whereas small and smooth patches of enamel are preserved on the labial surface of the crown. Conversely, the surface of MCF-PVPH-743 is slightly wrinkled on the lingual side of the crown base, whereas in MCF-PVPH-880-1 it has several longitudinal grooves that fade apically (Fig. 5). The crowns show different wear facets, of which the apical is rounded and wide, whereas the mesial and distal ones are narrower and apicobasally extended. All the specimens, except MCF-PVPH-879-9, have both distal and mesial carinae, without denticles. MCF-PVPH-879-9 is incomplete, lacking part of the basal and apical portions. Moreover, it seems to have only one distal carina; however, taking into account that the crown is heavily weathered, without most of the dentine and the enamel surface, especially on the mesial surface, the loss of the mesial carina due to diagenetic process should be taken into consideration.
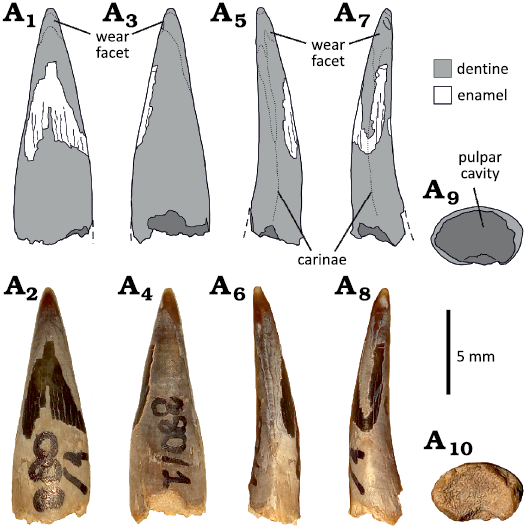
Fig. 5. Isolated tooth (Morphotype 2) from the Cerro de los Leones locality, Albian, Lower Cretaceous, Picún Leufú, Argentina. MCF-PVPH-880-1 in labial (A1, A2), lingual (A3, A4), distal (A5, A6), and mesial (A7, A8) views. The basal cross section (A9, A10) is visible due to the natural breaking of the crown.
Morphotype 3.—MCF-PVPH-741, 875, 879-1, 879-2, 879-3, 879-5, 879-7, 879-11, and 880-5 (Fig. 6). These tooth crowns are more recurved distally than lingually, with oval, labiolingually compressed cross-sections, and smooth dentine surfaces. The apical wear facet is rounded, worn at the apex from multiple directions, especially mesially and distally. All the specimens lack carinae. MCF-PVPH-875 preserves two portions of smooth enamel on the labial and distal surfaces, whereas in MCF-PVPH-879-2 the enamel is well preserved, especially on the basal half of the crown, and wrinkled, showing several apicobasally oriented and anastomosed ridges and grooves, slightly deeper on the lingual surface. A similar condition is observed on the preserved enamel of MCF-PVPH-879-1, 879-5, and 880-5, whereas in MCF-PVPH-879-3 the enamel is apically worn, with apicobasally oriented grooves on the basal half of the crown. The crowns of MCF-PVPH-875, 879-2, and 879-3 feature labial wear on their apexes and mesial and distal wear that visibly rounds their apices.
MCF-PVPH-741 shows a wide and smooth groove, slightly deeper basally and apicobasally extended to the apical third of the crown.
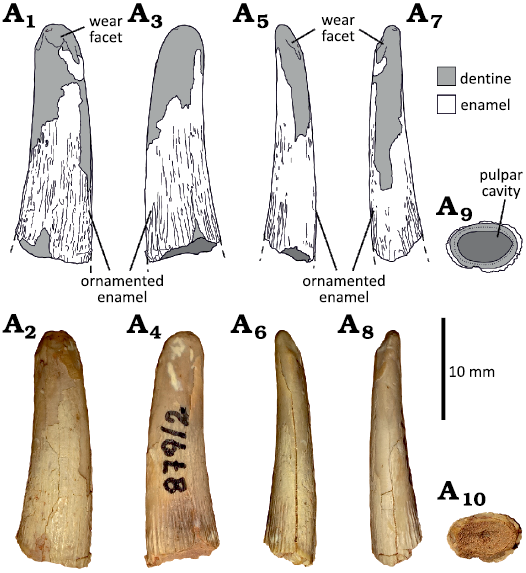
Fig. 6. Isolated tooth (Morphotype 3) from the Cretaceous Cerro de los Leones locality, Picún Leufú, Argentina. MCF-PVPH-879-2 in labial (A1, A2), lingual (A3, A4), distal (A5, A6), and mesial (A7, A8) views. The basal cross section (A9, A10) is visible due to the natural breaking of the crown.
Discussion
Comparisons
Morphotype 1.—The main morphology of these teeth slightly resembles the teeth of different fishes reported from Middle Jurassic to Upper Cretaceous sediments of the Neuquén Basin, such as the actinopterygian groups Pachycormiformes and Aspidorhynchiformes (Gouiric-Cavalli 2015; Gouiric-Cavalli and Arratia 2021). The teeth of the Patagonian pachycormiformid Kaykay lafken have a well ornamented collar enameloid with apicobasally oriented ridges, which are more prominent basally (Gouiric-Cavalli and Arratia 2021). However, the anterior procumbent teeth of Kaykay are typically under 10 mm in crown height and their cross-section is circular, while the teeth from Cerro de los Leones are bigger and show a compressed subcircular cross-section. Similarly, the internal and middle predentary tooth rows of the aspidorhynchiform fish order have conical teeth, but their size is considerably smaller than any of the specimens described in this contribution, which also lack the diagnostic accessory cusp seen in this clade (Gouiric-Cavalli 2015). Moreover, MCF-PVPH-879-4, 879-6, 879-8, and 879-10 differ from plesiosaur teeth for the apicobasally fluted surface texture on the lingual side of the crown, with flutes often bifurcating towards the base (O’Gorman and Varela 2010). On the other hand, the enamel texture, the crown inclination, and the cross-section shape of the specimens strongly resemble ornithocheiriform teeth, as observed in the anhanguerids Anhanguera (Kellner and Tomida 2000; Fastnacht 2001; Pinheiro and Rodrigues 2017), Cearadactylus (Vila Nova et al. 2014), Ludodactylus (Frey et al. 2003), Guidraco (Wang et al. 2012), Siroccopteryx (Mader and Kellner 1999), the hamipterid Hamipterus (Wang et al. 2014), and the targaryendraconian Barbosania (Elgin and Frey 2011). In particular, the teeth share their morphological traits with the most rostral teeth (those of the anterior “rosette”) of these pterosaurs, which are the largest teeth of their dentition. MCF-PVPH-879-8 is also very similar to the ornithocheiriform isolated teeth from Morocco BSP 1993 IX 618 and 621 (Wellnhofer and Buffetaut 1999). In this sense, the authors describe the smooth and rounded distal surface of the latter Moroccan teeth as a carina, however we prefer to reconsider this feature, describing the specimens as being without carinae but with a convex lingual surface and an almost straight labial surface. This condition is widespread among similar isolated teeth of Ornithocheiriformes, such as the Russian specimen RSU DGE 2020 RO MP 35 (Solonin et al. 2021) and the Tunisian specimen RH45 (Martill et al. 2018).
Morphotype 2.—The unserrated carinae of all the specimens are not as sharp as those observed in crocodyliforms such as Eutretauranosuchus (Smith et al. 2010), or in spinosaurine theropod dinosaurs (Richter et al. 2012), but more closely resemble some titanosaur sauropod teeth (García and Cerda 2010). However, titanosaur teeth are straight and have well developed wear facets labially and lingually (García and Cerda 2010), while the Cerro de los Leones teeth are more recurved and have rounded tips. The sharpness of the carinae is even less pronounced than in titanosaur teeth, in which the labial and lingual surfaces contact to form a sharp edge.
MCF-PVPH-880-3 shares with spinosaurine theropod dinosaurs a slight labiolingual compression of the crown, a gentle lingual curvature, unserrated mesial and distal carinae and deep apicobasally oriented ridges (Richter et al. 2012). However, it differs in the less pronounced carinae, the combination of lingual and distal inclination and a labiolingual compression of the crown, conditions not regarded in the tooth morphology of Spinosaurinae. Instead, all the specimens (MCF-PVPH-739-2, 743, 879-9, 880-1, 880-2, 880-3, 880-4) are morphologically similar to isolated ornithocheiriform pterosaur teeth such as UFMA 1.20.166 (Elias et al. 2007), BSP 1993 IX 314 (Wellnhofer and Buffetaut 1999), and FSAC KK 17001 (Martill et al. 2018). This tooth morphology is also observed in Anhanguera piscator NSM-PV 19892 (Kellner and Tomida 2000) and Anhanguera sp. SMNK 2302 PAL (Fastnacht 2001; Pinheiro and Rodrigues 2017), where the teeth are preserved in situ. In particular, the pterosaur teeth from Cerro de los Leones share the elliptical cross-section and the lingual inclination of the crown with the 5th and 6th teeth of Anhanguera, which are smaller, more labiolingually compressed and straighter than the contiguous teeth. Moreover, these conditions are shared with the last premaxillary teeth and the first maxillary teeth (i.e., after the 6th tooth) of Cearadactylus atrox (Vila Nova et al. 2014), the fourth tooth of the Ferrodraco lentoni (Pentland et al. 2022), the 6th tooth of Ludodactylus sibbicki (Frey et al. 2003), the 6th tooth of the upper jaw and the 4th tooth of the lower jaw of Guidraco venator (Wang et al. 2012), and the 4th tooth of the Siroccopteryx moroccensis holotype (Mader and Kellner 1999). Outside of Anhangueria, the pterosaur teeth from Cerro de los Leones share the main morphology with the several specimens of Hamipterus tianshanensis (Wang et al. 2014). MCF-PVPH-880-4 is incomplete, lacking the apical extremity. The base of the crown shows an oval, wide, and apicobasally extended smooth surface, which we interpreted as a tooth-to-tooth wear surface due to the eruption tooth.
Morphotype 3.—The main morphology of these teeth from Cerro de los Leones (MCF-PVPH-741, 875, 879-1, 879-2, 879-3, 879-5, 879-7, 879-11, 880-5), with oval cross-section, distally inclined crown, and without carinae, is shared with teeth posterior to the “rosette” of Anhanguera (Kellner and Tomida 2000; Fastnacht 2001; Pinheiro and Rodrigues 2017), which are gradually smaller and more distally inclined than more rostral teeth. A similar morphology is seen in the lower teeth of Mythunga camara (Molnar and Thulborn 2007), the 5th and 6th teeth of Ferrodraco lentoni (Pentland et al. 2022), the 9th lower, 10th upper, and posterior teeth of Guidraco venator (Wang et al. 2012), Ludodactylus sibbicki (Frey et al. 2003), Liaoningopterus gui (Rodrigues et al. 2015), and Hamipterus tianshanensis (Wang et al. 2014). The isolated tooth LINHM 007 (Kellner and Mader 1997) from Morocco also shares the same anatomical features. Furthermore, the groove observed in MCF-PVPH-741 can be interpreted as a tooth-to-tooth wear facet due to the growth of a replacement tooth, reflecting the tooth replacement pattern seen in other toothed pterosaurs, such as Anhanguera piscator (Kellner and Tomida 2000) and CSRL 12692/12713 (Dalla Vecchia 1993). Indeed, both specimens are reported to have several alveoli hosting two teeth, one functional and one growing tooth; this condition would suggest that there was a period of coexistence between older tooth and replacement tooth during the growth of the latter. Finally, Fastnacht (2008) observed that the teeth were shed when their replacement had reached about 60% of the full-grown height, similarly to extension of the posterior groove in MCF-PVPH-741.
Taxonomic attribution
The isolated teeth from Cerro de los Leones share several morphological features with the targaryendraconian Barbosania (Elgin and Frey 2011) and known pterosaur taxa of two more inclusive clades: Anhangueridae and Hamipteridae. Therefore, considering these morphological comparisons, together with the isolated and fragmentary conditions, the tooth specimens here described are considered indeterminate members of Ornithocheiriformes. More exclusive taxonomic attributions are difficult due to the lack of known diagnostic features in ornithocheiriform isolated teeth, as Solonin et al. (2021) recently noticed. The morphological variability of the teeth from Cerro de los Leones reflects the heterodont dentition of other ornithocheiriforms, which varies in cross-section shape, curvature, and size of the tooth crowns across the tooth row (Wellnhofer 1985, 1987). Moreover, ornithocheiriforms share a rostral “rosette” (Kellner and Tomida 2000) with more elongated, lingually and distally recurved teeth with respect to more posterior teeth (post-“rosette” and posterior teeth). These teeth are weakly compressed and the enamel has longitudinal, anastomosed, and deep grooves, as seen in Anhanguera piscator, Mythunga camara, and Guidraco venator (Kellner and Tomida 2000; Molnar and Thulborn 2007; Wang et al. 2012). Posteriorly to the “rosette”, the teeth are smaller, more labiolingually compressed and slightly lingually inclined (Mader and Kellner 1999; Kellner and Tomida 2000; Frey et al. 2003; Vila Nova et al. 2014). The most posterior ornithocheiriform teeth are progressively smaller, more sparse, more distally curved, and more labiolingually compressed, as observed in A. piscator and G. venator (Kellner and Tomida 2000; Frey et al. 2003; Wang et al. 2012). Moreover, the posterior dentition of some ornithocheiriforms lacks the wrinkled enamel (Wellnhofer 1985), as exemplified by the isolated tooth described by Fletcher and Salisbury (2010). Indeed, as observed by Pentland et al. (2022), the differences of the enamel surface between anterior and posterior ornithocheiriform teeth, such as A. piscator (Kellner and Tomida 2000), would be more concordant with a differential wear pattern than with morphological interspecific variations. In this sense, the tooth apex is often smoother than the basal portion, and smaller posterior teeth are often smoother than the bigger anterior teeth. However, in the Chinese Liaoningopterus (Rodrigues et al. 2015) all teeth are smooth, proving that distinguishing ornithocheiriforms by their teeth alone is currently challenging (Solonin et al. 2021).
Finally, the assignment of the teeth from Cerro de los Leones to Ornithocheiriformes is concordant with Martinelli et al. (2007) and Bellardini and Codorniú (2019), which adduce more taxonomic affinities with anhanguerids than other pterosaurs for the teeth MCF-PVPH-741 and MCF-PVPH-743, and the ulna MCF-PVPH-881, respectively.
Implications on ornithocheiriform dental reconstruction
The definition of the three dental morphotypes here recognized, allows us to compare them to the matching heterodont dentition of ornithocheiriforms. Indeed, ornithocheiriform pterosaurs bear rostral “rosette” teeth consistent with Morphotype 1, post-“rosette” teeth that correspond to Morphotype 2, and posterior teeth representing the Morphotype 3 (Fig. 7).
Taking into account the interspecific, intraspecific and ontogenic variation of tooth morphology within Ornithocheiriformes (Vullo and Neraudeau 2009), the limits of this partition are tentative and more focused to identify functional areas than to strictly define specific tooth positions. Moreover, it should be taken into consideration that tooth morphology often varies gradually across the tooth row. Finally, in order to allow morphological and anatomical attributions of isolated ornithocheiriform teeth, a new approach using the following morphological data matrix is here proposed (Table 2; Fig. 8).
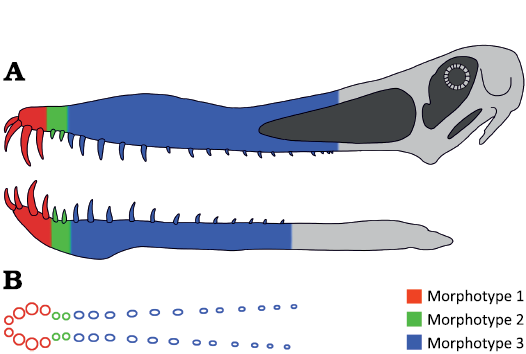
Fig. 7. Tooth morphotype distribution in ornithocheiriform upper and lower jaw (A) with associated crown size and cross-section variation (B). Example based on Anhanguera piscator (Kellner and Tomida 2000).

Fig. 8. Life reconstruction of the Cerro de los Leones pterosaur, displaying the morphological diversity of its dentition based on the observed distribution of the morphotypes. Illustration by Alessio Ciaffi.
Table 2. Morphological characterization of ornithocheiriform dentition to aid identification of isolated teeth, where each morphotype is assigned to a mouth area.
| |
Crown |
Cross-section shape |
Position |
|
Morphotype 1 |
more lingual than distal |
subcircular |
“rosette” |
|
Morphotype 2 |
more lingual than distal |
elliptical |
post-“rosette” |
|
Morphotype 3 |
more distal than lingual |
oval |
posterior |
Ornithocheiriform dental partition
Following the resulting three morphotypes, isolated ornithocheiriform teeth can be assigned to a specific dental area (i.e., “rosette”, post-“rosette”, posterior tooth row). To test this new approach, and the consistency of the chosen morphological features and the resulting morphotypes of the double-entry matrix, we analyze worldwide known evidence of isolated ornithocheiriform teeth in the pterosaur fossil record.
The isolated tooth crowns from the Cretaceous Kem Kem beds (Morocco) were assigned to “Ornithocheiridae” by Wellnhofer and Buffetaut (1999), which recognized four morphotypes. However, the specimens (BSP 1993 IX 314, 590–596; BSP 1993 IX 332, 608–617) of two morphotypes (morphotypes I and III) of their contribution share strong labiolingual compression, often smooth enamel and distally inclined crown, as the new Morphotype 3 here proposed. Following the new approach, they are therefore considered posterior teeth. The specimens (BSP 1993 IX 314, 597–607) they referred to as morphotype II have carinae and an almost mesiodistally straight and labiolingually inclined crown, as the new Morphotype 2 attributed to the post-“rosette” area. With respect to the isolated crowns (BSP 1993 IX 618–621) of the morphotype IV defined by the authors (Wellnhofer and Buffetaut 1999), they have a subcircular cross-section, strong lingual inclination of the crown and a wrinkled enamel, as the new Morphotype 1. Furthermore, Wellnhofer and Buffetaut (1999) report the presence of a distal carina on the teeth of their last morphotype, however we consider this feature a faint border, not as defined as a carina, caused by the asymmetrical shape of the crown. This feature is better observed in cross-section and is shared by other ornithocheiriform teeth (LRF 3142, Brougham et al. 2017; RSU DGE 2020 RO MP 36, SS106 5, SS106 6, Solonin et al. 2021; RH45, Martill et al. 2018).
The isolated tooth LINHM 007 from the Cretaceous of the Ksar es Souk Province (Morocco), which Kellner and Mader (1997) described as being very similar to those of anhanguerids from the Lower Cretaceous Santana Formation of northeastern Brazil, is consistent with Morphotype 3 and therefore recognized to belong to the ornithocheiriform posterior portion of the tooth row.
The complete isolated tooth FSAC KK 17001 from the Kem Kem beds of eastern Morocco mentioned by Martill et al. (2018) is lingually recurved and almost straight from mesial view, has an elliptical cross-section and preserved carinae. The same features are shared by an unnamed isolated tooth assigned to “Ornithocheiridae” by Benton et al. (2000), coming from the Chenini Formation of the Tataouine region in southern Tunisia. These features correspond to the Morphotype 2.
The isolated tooth RH45 from the Albian, Lower Cretaceous of Oum ed Diab Member of the Aïn el Guettar Formation of southern Tunisia (Martill et al. 2018) is gently lingually recurved, ornamented and has a subcircular cross-section. Moreover, the authors proposed that the tooth could be from the anterior part of the jaws, likely from alveolar pair 3 to 4 if from the upper dentition. This information further supports its identification as a Morphotype 1 tooth.
The isolated crown MA FVA 2 from Fouras-Vauban, France (Vullo and Neraudeau 2009) is more recurved distally than lingually, with oval cross-section shape, as described in Morphotype 3. Two other crowns from the same contribution, MA FDB 2 and MA RND 1, are weakly curved labiolingually, but are not recurved distally. This assigns them to Morphotype 2.
The isolated tooth NNPM2588-62 from the Albian, Lower Cretaceous of Burim Formation (Ukraine), assigned by Sokolskyi (2023) to Anhangueria, shows an oval cross-section and is posteriorly recurved, sharing the apical wear facets with some Morphotype 3 teeth as well.
Solonin et al. (2021) describe several isolated teeth from the Upper Cretaceous Dmitrov Formation of Malyy Prolom (Russia). RSU DGE 2020 RO MP 35 shows subcircular cross-section and distal curvature, features shared with Morphotype 1, and referred to the anterior “rosette”. The straight, labiolingually compressed crown with carinae of RSU DGE 2020 RO MP 37 is consistent with the small teeth of Morphotype 2, like MCF-PVPH-880-2. On the other hand, RSU DGE 2020 RO MP 36, SS106 5 and SS106 6 are oval in cross-section and posteriorly recurved, and are considered as some of the first, larger, posterior teeth of Morphotype 3.
Averianov (2007) describes several isolated teeth from the Lower Creaceous of Russia and Uzbekistan, of which ZIN PH 45/44 shows a lingually recurved crown with elliptical cross-section and carinae, morphological features of Morphotype 2. On the other hand, ZIN PH 15/43, ZIN PH 41/43 and ZIN PH 49/44 have distally inclined and labiolingually compressed crowns, as in Morphotype 3.
The isolated teeth LRF 759 and LRF 3142 from the Lower Cretaceous Griman Creek Formation (Australia) were referred to Anhangueridae by Brougham et al. (2017). These crowns are labiolingually compressed, distally inclined, and show a mostly smooth surface, corresponding with Morphotype 3. While LRF 759 lacks carinae, the authors attribute to LRF 3142 mesial and distal carinae. We prefer to refer to them as narrow edges, since the distal edge transitions smoothly from being acute on the apical half of the crown to more curved on the basal portion of the crown (Brougham et al. 2017).
Elias et al. (2007) report several isolated teeth from the Cretaceous of the São Luís-Grajaú Basin (Brazil), of which UFMA 1.20.130, 1.20.1744, 1.20.178-1, 1.20.130 and URC R.77-1, R.77-2, R.77-3, R.77-4, R.77-5 show distally recurved crowns with oval cross-sections, characters of Morphotype 3, suggesting a posterior position in the tooth row. On the other hand, the lingually inclined and symmetrically compressed crown of UFMA 1.20.166 is consistent with Morphotype 2.
Paleobiological implications
The new pterosaur evidence from Cerro de los Leones, with different tooth sizes and morphologies, suggests a more abundant and diversified ornithocheiriform fauna than known to date. Moreover, morphological analysis and comparisons allow us to reconstruct part of the tooth anatomy of ornithocheiriforms, suggesting some degree of partition in their dentition. The resulting anatomical attribution of isolated ornithocheiriform teeth allows the approximation of the cranial size of the owner of one specimen. In this context, assuming that the Cerro de los Leones pterosaurs had similar cranial proportions to those of the complete skull of the related A. piscator described by Kellner and Tomida (2000), an approximate estimation of the minimum skull length of the owner of MCF-PVPH-879-8 can be explored. The morphology of MCF-PVPH-879-8 identifies it as a Morphotype 1 tooth, attributed to the anterior “rosette”. In A. piscator (Kellner and Tomida 2000) the third tooth is the largest, as it often is in ornithocheiriforms. Therefore, assuming MCF-PVPH-879-8 to have been the third, largest tooth, a minimum skull size estimation can be carried on. The third tooth crown height is 61 mm in NSM-PV 19892, whereas in MCF-PVPH-879-8 it is 27 mm. The cranial length of NSM-PV 19892 measured by Kellner and Tomida (2000) is approximately 625 mm. Scaling the Cerro de los Leones tooth MCF-PVPH-879-8 with the size of the in situ tooth of A. piscator, the resulting estimated cranial length is 276 mm. Considering that Aureliano et al. (2014) estimated the wingspan of the same A. piscator specimen to be at least 4 m, a scaled wingspan for MCF-PVPH-879-8 would be approximately 2 m. This value is consistent with the wingspan estimation obtained by Bellardini and Codorniú (2019) on the pterodactyloid ulna uncovered in the same locality (3.2 m). Moreover, the incomplete tooth crown MCF-PVPH-879-10, here referred to Morphotype 1, is larger than MCF-PVPH-879-8, suggesting the presence of larger pterosaur specimens in Cerro de los Leones. This result, however, can be further discussed only with new fossil data. Skull morphology in ornithocheiriforms such as Anhanguera was variable and changed throughout ontogeny (Bantim et al. 2015; Duque et al. 2022), making any size estimation tentative. Nevertheless, an approximate size range estimation represents important biometric information for the paleoecosystem reconstruction of Cerro de los Leones. Detailed knowledge on the body-size of the southern Neuquén Basin pterosaurs allows us to consider what kind of paleoenvironmental conditions and feeding resources were available during the Albian to provide nutrients to populations with different small, medium or large flying reptiles.
Paleobiogeographical and paleoecological implications
To date, evidence of Ornithocheirifomes in Argentina is composed of a distal portion of a left ulna from the Río Belgrano Formation (Barremian) of the Santa Cruz Province (MACN-Pv SC3617d, ex MACN-SC-3617), which was described as closely related to Anhangueridae by Kellner et al. (2003), and a proximal half of an ulna from the Lohan Cura Formation (Albian) of the Neuquén Province (Bellardini and Codorniú 2019). The latter ulna specimen and the teeth of this contribution, all from Cerro de los Leones, represent a recent addition to the record of Ornithocheiriformes for the Neuquén Basin. This evidence suggests that ornithocheiriforms continued inhabiting Patagonia after the Barremian and continued being part of faunal assemblages at least up to the Albian. Therefore, these pterosaurs were potentially contemporary to many of their Gondwanan relatives (Upchurch et al. 2015). Moreover, the ulna from the Río Belgrano Formation was deposited in a nearshore environment (Kellner et al. 2003), while the new Cerro de los Leones record is part of a fluvial depositional sequence (Martinelli et al. 2007). This information reveals that, at least during the Albian, this pterosaur clade had extended its distribution from coastal, marine environments of Argentina to continental bodies of water.
The fossil record of Ornithocheiriformes is often linked to shallow-water, marine, fluvial and lagoonal depositional environments (Upchurch et al. 2015), as inferred for the Lohan Cura Formation cropping out at the Cerro de los Leones locality (Leanza and Hugo 2011; Martinelli et al. 2007). In this sense, different authors correlate the occurrence of pterosaurs in these paleoenvironments with a diet consisting primarily of fish and other aquatic organisms; this assumption is also supported by several cranial and dental morphological features of ornithocheiriforms (Zhou et al. 2017; Bestwick et al. 2018, 2020). Indeed, Ornithocheiriformes shares the heterodont condition with different extant and extinct vertebrates, such as congrid fishes (Congridae) and spinosaurid dinosaurs (Vullo et al. 2016), which are adapted to feed on evasive prey in aquatic environments. Indeed, they show a premaxillary and dentary “rosette” with enlarged, apicobasally elongated, slightly distally inclined and conical teeth, which, together with anteroposteriorly long and laterally compressed jaws, confer a high biting speed and a low drag during predation (Vullo et al. 2016). Ornithocheiriform pterosaurs, similarly to what was pointed out for spinosaurid dinosaurs by Vullo et al. (2016), also share with anguilliform fishes a notch posterior to the “rosette”, bearing smaller teeth. These features combined improve the prey capture kinematics in anguilliforms (Eagderi and Adriaens 2010) and could have represented an analogous advantage for ornithocheiriforms. Although reconstruction of pterosaur diet is not the aim of this contribution, the new data from Cerro de los Leones can contribute to understanding the morphological diversification of ornithocheiriform dental anatomy, which is closely related to the feeding strategies of these flying reptiles. Finally, considering the pterosaur, fish, and mollusk fossil remains from Cerro de los Leones (Martinelli et al. 2007), new, more complete, well preserved specimens will improve our knowledge of the trophic relationships between the pterosaurs and the freshwater fauna in the southern Neuquén Basin during the Albian.
Conclusions
The isolated pterosaur teeth from the Albian, fluvial outcrops of the Cerro de los Leones locality (Patagonia, Argentina) are here referred to different individuals of indeterminate specimens of Ornithocheiriformes. These dental remains, together with the postcranial evidence from the same locality (Bellardini and Codorniú 2019), suggest a more abundant and diversified ornithocheiriform fauna in the south of Neuquén Basin than previously known, at least during the Albian. Moreover, a morphological analysis was carried out and three inclusive morphotypes were distinguished. The comparisons of the new evidence from Cerro de los Leones with more complete and better preserved anhanguerian and targaryendraconian specimens, suggests that ornithocheiriform pterosaurs had a partitioned heterodont dentition, where three main dental regions are recorded: a most anterior, rostral “rosette” region, with a tooth morphology consistent with Morphotype 1; a post-“rosette” region, with a tooth morphology corresponding with Morphotype 2; a posterior region, where the teeth fit with Morphotype 3. In this contribution we also suggest a new approach to carry out a preliminary taxonomic attribution and an anatomical assignment of isolated teeth to a functional area of the heterodont ornithocheiriform tooth row. The understanding of ornithocheiriform dental morphology, together with information on the depositional environment of the Lohan Cura Formation and the diversified fossil record from Cerro de los Leones locality (Martinelli et al. 2007), has paleoecological implications, suggesting a piscivorous adaptation for the flying reptiles that inhabited the southern Neuquén Basin during the Albian. Finally, considering the scarce Ornithocheiriformes fossil record for Argentina, the new evidence from Cerro de los Leones improves our knowledge of the taxonomic diversification and paleobiogeographic evolution of the Lower Cretaceous pterosaurs in the Southwest of Gondwana.
Acknowledgements
The authors wish to thank Rodolfo A. Coria and Ludmila Coria (both MCF-PVPH) for allowing access to the paleontological collection of the Museo Municipal “Carmen Funes” of Plaza Huincul, Raffaele Sardella (Dipartimento di Scienze della Terra, Sapienza Università di Roma, Rome, Italy) for encouraging the project, and Agustin G. Martinelli (Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires, Argentina) for providing access to the fossil material under his care and updating the collection number of a specimen of this contribution. The authors thank Rodrigo Pêgas (Laboratório de Paleontologia de Vertebrados e Comportamento Animal, Universidade Federal do ABC, São Bernardo do Campo, Brazil) and an anonymous reviewer for their helpful insights and reviews which improved the quality of the manuscript. This research was financed by the grant of Sapienza Università di Roma: Borsa Tesi Estero n. 18/2021 (AC). This manuscript was accomplished as part of the M.Sc. Thesis project, Sapienza Università di Roma (AC) directed by Raffaele Sardella.
References
Alarcón-Muñoz, J., Codorniú, L., González, E., Suárez, M.E., Suárez, M., Vicencio-Campos, O., Soto-Acuña, S., Kaluza, J., Vargas, A.O., and Rubilar-Rogers, D. 2022. A new locality with ctenochasmatid pterosaurs (Pterosauria: Pterodactyloidea) in the Atacama Desert, Northern Chile. Cretaceous Research 135: 105173. Crossref
Alarcón-Muñoz, J., Soto-Acuña, S., Codorniú, L., Rubilar-Rogers, D., Sallaberry, M., and Suárez, M. 2020. New ctenochasmatid pterosaur record for Gondwana: discovery in the Lower Cretaceous continental deposits of the Atacama Desert, northern Chile. Cretaceous Research 110: 104378. Crossref
Andres, B. 2021. Phylogenetic systematics of Quetzalcoatlus Lawson 1975 (Pterodactyloidea: Azhdarchoidea). Journal of Vertebrate Paleontology 41: 203–2017. Crossref
Andres, B., Clark, J., and Xu, X. 2014. The earliest pterodactyloid and the origin of the group. Current Biology 24: 1011–1016. Crossref
Aureliano, T., Ghilardi, A.M., Duque, R.R., and Barreto, A.M. 2014. On the occurrence of Pterosauria in Exu, Pernambuco (Lower Cretaceous Romualdo Formation, Araripe Basin) Northeastern Brazil. Estudos Geológicos 24 (2): 15–27. Crossref
Averianov, A.O. 2007. Mid-cretaceous Ornithocheirids (Pterosauria, Ornithocheiridae) from Russia and Uzbekistan. Paleontological Journal 41: 79–86. Crossref
Bantim, R.A.M., Saraiva, A.A.F., and Sayão, J.M. 2015. Skull variation and the shape of the sagittal premaxillary crest in anhanguerid pterosaurs (Pterosauria, Pterodactyloidea) from the Araripe Basin, Northeast Brazil. Historical Biology 27: 656–664. Crossref
Barrett, P.M., Butler, R.J., Edwards, N.P., and Milner, A.R. 2008. Pterosaur distribution in time and space: an atlas. Zitteliana 28: 61–108.
Bellardini, F. and Cerda, I.A. 2017. Bone histology sheds light on the nature of the “dermal armor” of the enigmatic sauropod dinosaur Agustinia ligabuei Bonaparte, 1999. The Science of Nature 104: art. 1. Crossref
Bellardini, F. and Codorniú, L. 2019. First pterosaur post-cranial remains from the Lower Cretaceous Lohan Cura Formation (Albian) of Patagonia, Argentina. Ameghiniana 56: 116–134. Crossref
Bellardini, F., Coria, R.A., Pino, D.A., Windholz, G.J., Baiano, M.A., and Martinelli, A.G. 2022. Osteology and phylogenetic relationships of Ligabuesaurus leanzai (Dinosauria: Sauropoda) from the Early Cretaceous of the Neuquén Basin, Patagonia, Argentina. Zoological Journal of the Linnean Society 196: 1333–1393. Crossref
Benton, M.J., Bouaziz, S., Buffetaut, E., Martill, D., Ouaja, M., Soussi, M., and Trueman, C. 2000. Dinosaurs and other fossil vertebrates from fluvial deposits in the Lower Cretaceous of southern Tunisia. Palaeogeography, Palaeoclimatology, Palaeoecology 157: 227–246. Crossref
Bestwick, J., Unwin, D.M., Butler, R.J., and Purnell, M.A. 2020. Dietary diversity and evolution of the earliest flying vertebrates revealed by dental microwear texture analysis. 2020. Nature Communications 11: 5293. Crossref
Bestwick, J., Unwin, D.M., Butler, R.J., Henderson, D.M., and Purnell, M.A. 2018. Pterosaur dietary hypotheses: a review of ideas and approaches. Biological Reviews 93: 2021–2048. Crossref
Blackburn, D. 2002. Two Early Cretaceous pterosaurs from Africa. Journal of Vertebrate Paleontology 22: 37A.
Bonaparte, J.F. 1999. An armoured sauropod from the Aptian of northern Patagonia, Argentina. National Science Museum Monographs 15: 1–12.
Bonaparte, J.F., Gonzaléz Riga, B.J., Apesteguía, S., 2006. Ligabuesaurus leanzai gen. et sp. nov. (Dinosauria, Sauropoda), a new titanosaur from the Lohan Cura Formation (Aptian, Lower Cretaceous) of Neuquén, Patagonia, Argentina. Cretaceous Research 27: 364–376. Crossref
Brougham, T., Smith E.T., and Bell, P.R. 2017. Isolated teeth of Anhangueria (Pterosauria: Pterodactyloidea) from the Lower Cretaceous of Lightning Ridge, New South Wales, Australia. PeerJ 5: e3256. Crossref
Claessens, L.P.A., O’Conner, P.M., and Unwin, U.M. 2009. Respiratory Evolution Facilitated the Origin of Pterosaur Flight and Aerial Gigantism. PLoS ONE 4 (2): e4497. Crossref
Codorniú, L. and Chiappe, L.M. 2004. Early juvenile pterosaurs (Pterodactyloidea: Pterodaustro guinazui) from the Lower Cretaceous of central Argentina. Canadian Journal of Earth Sciences 41: 9–18. Crossref
Codorniú, L. and Gianechini, F.A. 2016. The flying reptiles from Argentina: An overview. Revista del Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Special Volume 6: 85–95.
Codorniú, L., Chiappe, L.M., and Cid, F.D. 2013. First occurrence of stomach stones in pterosaurs. Journal of Vertebrate Paleontology 33: 647–654. Crossref
Codorniú, L., Chiappe, L.M., and Rivarola, D. 2018. Neonate morphology and development in pterosaurs: evidence from a ctenochasmatid embryo from the Early Cretaceous of Argentina. Geological Society, London, Special Publications 455: 83–94. Crossref
Coria, R. A. 2022. Pequeña historia de la paleoherpetología en el museo Carmen Funes de Plaza Huincul (Neuquén, Argentina): Hechos y protagonistas. Publicación Electrónica de la Asociación Paleontológica Argentina 22: 326–334. Crossref
Dalla Vecchia, F.M. 1993. Cearadactylus? ligabuei, nov. sp., a new Early Cretaceous (Aptian) pterosaur from Chapada do Araripe (Northeastern Brazil). Bollettino della Società Paleontologica Italiana 32: 401–409.
Dalla Vecchia, F.M. 2003. A review of the Triassic pterosaur fossil record. Rivista del Museo Civico di Scienze Naturali “E. Caffi” di Bergamo 22: 13–29.
Duque, R.R.C., Pinheiro, F.L., and Barreto, A.M.F. 2022. The ontogenetic growth of Anhangueridae (Pterosauria, Pterodactyloidea) premaxillary crests as revealed by a crestless Anhanguera specimen. Journal of Vertebrate Paleontology 42 (1): e2116984. Crossref
Eagderi, S. and Adriaens, D. 2010. Head morphology of the duckbill eel, Hoplunnis punctata (Regan, 1915; Nettastomatidae: Anguilliformes) in relation to jaw elongation. Zoology 113: 148–157. Crossref
Elgin, R.A. and Frey, E. 2011. A new ornithocheirid, Barbosania gracilirostris gen. et sp. nov. (Pterosauria, Pterodactyloidea) from the Santana Formation (Cretaceous) of NE Brazil. Swiss Journal of Palaeontology 130: 259–275. Crossref
Elgin, R.A. and Hone D.W.E. 2013. Pneumatization of an immature azhdarchoid pterosaur. Cretaceous Research 45: 16–24. Crossref
Elias, F., Bertini, R., and Medeiros, M. 2007. Pterosaur teeth from the Laje do Coringa, middle Cretaceous, São Luís-Grajaú basin, Maranhão state, Northern-Northeastern Brazil. Revista Brasileira de Geociências 37: 668–676. Crossref
Fastnacht, M. 2001. First record of Coloborhynchus (Pterosauria) from the Santana Formation (Lower Cretaceous) of the Chapada do Araripe, Brazil. Paläontologische Zeitschrift 75: 23–36. Crossref
Fastnacht, M. 2008. Tooth replacement pattern of Coloborhynchus robustus (Pterosauria) from the Lower Cretaceous of Brazil. Journal of Morphology 269: 32–48. Crossref
Fletcher, T.L. and Salisbury, S.W. 2010. New pterosaur fossils from the Early Cretaceous (Albian) of Queensland, Australia. Journal of Vertebrate Paleontology 30: 1747–1759. Crossref
Frey, E., Martill, D., and Buchy, M. 2003. A new crested ornithocheirid from the Lower Cretaceous of northeastern Brazil and the unusual death of an unusual pterosaur. Geological Society Special Publication 217: 55–63. Crossref
García, R.A. and Cerda, I. 2010. Dentición de titanosaurios (Dinosauria, Sauropoda) del Cretácico Superior de la provincia de Río Negro, Argentina: morfología, inserción y reemplazo. Ameghiniana 47: 45–60. Crossref
Gouiric-Cavalli, S. 2015. Jonoichthys challwa gen. et sp. nov., a new Aspidorhynchiform (Osteichthyes, Neopterygii, Teleosteomorpha) from the marine Upper Jurassic sediments of Argentina, with comments about paleobiogeography of Jurassic aspidorhynchids. Comptes Rendus de l’Académie des sciences 14 (4): 291–304. Crossref
Gouiric-Cavalli, S. and Arratia, G. 2021. A new †Pachycormiformes (Actinopterygii) from the Upper Jurassic of Gondwana sheds light on the evolutionary history of the group, Journal of Systematic Palaeontology 19: 1517–1550. Crossref
Hendrickx, C., Mateus, O., and Araújo, R. 2015. A proposed terminology of theropod teeth (Dinosauria, Saurischia), Journal of Vertebrate Paleontology 35 (5): e982797. Crossref
Kellner, W.A. 1994. Remarks on pterosaur taphonomy and paleoecology. Acta Geologica Leopoldensia 39: 175–189.
Kellner, A.W.A. 2003. Pterosaur phylogeny and comments on the evolutionary history of the group. In: E. Buffetaut and J.-M. Mazin (eds.), Evolution and Palaeobiology of Pterosaurs. Geological Society, London, Special Publications 217: 105–137. Crossref
Kellner, A.W.A. and Langston, W., Jr. 1996. Cranial remains of Quetzalcoatlus (Pterosauria, Azhdarchidae) from Late Cretaceous sediments of Big Bend National Park, Texas. Journal of Vertebrate Paleontology 16: 222–231. Crossref
Kellner, A.W.A. and Mader B.J. 1997 Archosaur teeth from the Cretaceous of Morocco. Journal of Paleontology 71: 525–527. Crossref
Kellner, A.W.A. and Tomida, Y. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaurfauna from the Santana Formation (Aptian–Albian), northeastern Brazil. National Science Museum Monographs 17: 1–135. National Science Museum, Tokyo.
Kellner, A.W.A., Campos, D.A., Sayão, J.M., Saraiva, A.A., Rodrigues, T., Oliveira, G., Cruz, L.A., Costa F.R., Silva H.P., and Ferreira, J.S. 2013. The largest flying reptile from Gondwana: a new specimen of Tropeognathus cf. T. mesembrinus Wellnhofer, 1987 (Pterodactyloidea, Anhangueridae) and other large pterosaurs from the Romualdo Formation, Lower Cretaceous, Brazil. Anais da Academia Brasileira de Ciências 85: 113–135. Crossref
Kellner, A.W.A., Aguirre-Urreta, M.B., and Ramos, V.A. 2003. On the pterosaur remains from the Río Belgrano Formation (Barremian), Patagonian Andes of Argentina. Anais da Academia Brasileira de Ciências 75 (4): 487–495. Crossref
Kellner, A.W.A., Caldwell, M.W., Holgado, B., Dalla Vecchia, F.M., Nohra, R., Sayão, J.M., and Currie, P.J. 2019. First complete pterosaur from the Afro-Arabian continent: insight into pterodactyloid diversity. Scientific Reports 9: 17875. Crossref
Knoll, F. 2000. Pterosaurs from the Lower Cretaceous (?Berriasian) of Anoual, Morocco. Annales de Paléontologie 86 (3): 157–164. Crossref
Leanza, H.A. 2003. Las sedimentitas huitrinianas y rayosianas (Cretácico Inferior) en el ámbito central y meridional de la Cuenca Neuquina, Argentina. Servicio Geológico Minero Argentino, Serie Contribuciones Técnicas-Geología, Buenos Aires 2: 131.
Leanza, H.A. and Hugo, C.A. 1997. Hoja Geológica 3969-III Picún Leufú, provincias del Neuquén y Río Negro. Boletín Instituto de Geología y Recursos Naturales, SEGEMAR 218: 1–135.
Leanza, H.A. and Hugo, C.A. 2011. Las Formaciones La Amarga y Lohan Cura (Cretácico temprano) en el Depocentro de Picún Leufú. In: H.A. Leanza, C. Arregui, O. Carbone, J.C. Danieli and J.M. Vallés (eds.), Geología y Recursos Naturales de la Provincia del Neuquén. Relatorio del 18º Congreso Geológico Argentino, 223–230. Asociación Geológica Argentina, Neuquén.
Long, R.A. and Murry, P.A. 1995. Late Triassic (Carnian and Norian) tetrapods from the southwestern United States. Bulletin of the New Mexico Museum of Natural History and Science 4: 1–254.
Lü J., Unwin DM, Xingsheng J., Liu Y., and Ji Q. 2009. Evidence for modular evolution in a long-tailed pterosaur with a pterodactyloid skull. Proceedings of the Royal Society B: Biological Sciences 277: 383–389. Crossref
Mader, B. and Kellner, A.W.A. 1999. A new anhanguerid pterosaur from the Cretaceous of Morocco. Boletim do Museu Nacional, Geologia, Nova Série 45: 1–11.
Martill, D.M., Ibrahim, N., and Bouaziz, S. 2018. A giant pterosaur in the Early Cretaceous (Albian) of Tunisia. Journal of African Earth Sciences 147: 331–337. Crossref
Martinelli, A.G., Agnolin, F.L., Ezcurra, M.D., Isasi, M.P., and Novas, F.E. 2022. El Museo Argentino de Ciencias Naturales “Bernardino Rivadavia” y los aportes a la paleoherpetología argentina. Publicación Electrónica de la Asociación Paleontológica Argentina 22: 188–207. Crossref
Martinelli, A.G., Garrido, A.C., Forasiepi, A.M., Paz. E.R., and Gurovich Y. 2007. Notes on fossil remains from the Early Cretaceous Lohan Cura Formation, Neuquén Province, Argentina, Gondwana Research 11: 537–552. Crossref
Martínez, R.N., Andres, B., Apaldetti, C., and Cerda, I.A. 2022. The dawn of the flying reptiles: first Triassic record in the southern hemisphere. Papers in Palaeontology 8: e1424. Crossref
Martínez, R.N., Apaldetti, C., Correa, G., Colombi, C.E., Fernández, E., Malnis, P.S., Praderio, A., Abelín, D., Benegas, L.G., Aguilar-Cameo, A., and Alcober, O.A. 2015. A new Late Triassic vertebrate assemblage from northwestern Argentina. Ameghiniana 52: 379–390. Crossref
Miall, A.D. 1996. The Geology of Fluvial Deposits. Sedimentary Facies, Basin Analysis, and Petroleum Geology. 582 pp. Springer, New York.
Molnar, R.E. and Thulborn, R.A. 1980. First pterosaur from Australia. Nature 288: 361. Crossref
Molnar, R.E. and Thulborn, R.A. 2007. An incomplete pterosaur skull from the Cretaceous of North-Central Queensland, Australia. Arquivos do Museu National, Río de Janeiro 65: 461–470.
Montanelli, S.B. 1987. Presencia de Pterosauria (Reptilia) en la Formación La Amarga (Hauteriviano–Barremiano) Neuquén, Argentina. Ameghiniana 24: 109–113.
O’Gorman, J.P. and Varela, A.N. 2010. The oldest lower Upper Cretaceous plesiosaurs (Reptilia, Sauropterygia) from southern Patagonia, Argentina. Ameghiniana 47: 447–459. Crossref
Padian, K. 1984. A large pterodactyloid pterosaur from the Two Medicine Formation (Campanian) of Montana. Journal of Vertebrate Paleontology 4: 516–524. Crossref
Pentland, A.H. and Poropat, S.F. 2023. A review of the Jurassic and Cretaceous Gondwanan pterosaur record. Gondwana Research 119: 341–383. Crossref
Pentland, A.H., Poropat, S.F., White, M.A., Rigby, S.L., Bevitt, J.J., Duncan, R.J., Sloan, T., Elliott, R.A., Elliott, H.A., Elliott, J.A., and Elliott, D.A. 2022. The osteology of Ferrodraco lentoni, an anhanguerid pterosaur from the mid-Cretaceous of Australia. Journal of Vertebrate Paleontology 41 (5): e2038182. Crossref
Pinheiro, F.L. and Rodrigues, T. 2017. Anhanguera taxonomy revisited: is our understanding of Santana Group pterosaur diversity biased by poor biological and stratigraphic control? PeerJ 5: e3285. Crossref
Price, L.I. 1971. A presença de Pterosauria no Cretáceo inferior da Chapada do Araripe, Brasil. Anais da Academia Brasileira de Ciências 43: 451–461.
Raja, N.B., Dunne, E.M., Matiwane, A., Khan, T.M., Nätscher, P.S., Ghilardi, A.M., and Chattopadhyay, D. 2021. Colonial history and global economics distort our understanding of deep-time biodiversity. Nature Ecology & Evolution 6: 145–154. Crossref
Richter, U., Mudroch, A., and Buckley, L.G. 2012. Isolated theropod teeth from the Kem Kem Beds (Early Cenomanian) near Taouz, Morocco. Paläontologische Zeitschrift 87: 291–309. Crossref
Reguero, M.A., Gasparini, Z., Olivero, E.B., Coria, R.A., Fernández, M.S., O’ Gorman, J.P., Gouiric-Cavalli, S., Hospitaleche, C.A., Bona, P., Iglesias, A., Gelfo, J.N., Raffi, M.E., Moly, J.J., Santillana, S.N., and Cárdenas, M. 2022. Late Campanian–early Maastrichtian vertebrates from the James Ross Basin, West Antarctica: updated synthesis, biostratigraphy, and paleobiogeography. Anais da Academia Brasileira de Ciências 94: e20211142. Crossref
Rodrigues T. and Kellner, A.W.A. 2010. Note on the pterosaur material described by Woodward from the Reconcavo Basin, Lower Cretaceous, Brazil. Revista Brasileira de Paleontologia 13 (2): 159–164. Crossref
Rodrigues, T., Jiang, S., Cheng, X., Wang, X., and Kellner, A.W.A. 2015. A new toothed pteranodontoid (Pterosauria, Pterodactyloidea) from the Jiufotang Formation (Lower Cretaceous, Aptian) of China and comments on Liaoningopterus gui Wang and Zhou, 2003. Historical Biology 27: 782–795. Crossref
Sánchez-Hernández, B., Benton, M.J., Naish, D. 2007. Dinosaurs and other fossil vertebrates from the Late Jurassic and Early Cretaceous of the Galve area, NE Spain. Palaeogeography, Palaeoclimatology, Palaeoecology 249: 180–215. Crossref
Smith, D.K., Allen, E.R., Sanders, R.K., and Stadtman, K.L. 2010. A new specimen of Eutretauranosuchus (Crocodyliformes; Goniopholididae) from Dry Mesa, Colorado. Journal of Vertebrate Paleontology 30: 1466–1477. Crossref
Sokolskyi, T. 2023. First occurrence of pterosaurs in Ukraine from freythe Albian (Lower Cretaceous) Burim Formation, Kaniv Natural Reserve. Journal of Vertebrate Paleontology 43 (1): e2238000 Crossref
Solonin, S.V., Martill, D.M., Smith, R.E., and Vodorezov, A.V. 2021. First occurrence of ornithocheirid pterosaur teeth in the Dmitrov Formation (Santonian) of Ryazan Oblast, Russia. Cretaceous Research 127: 104943. Crossref
Unwin, D.M. 2003. On the phylogeny and evolutionary history of pterosaurs. Geological Society, London, Special Publications 217: 139–190. Crossref
Upchurch, P., Andres, B., Butler, R.J., and Barrett, P.M. 2015. An analysis of pterosaurian biogeography: implications for the evolutionary history and fossil record quality of the first flying vertebrates. Historical Biology 27: 697–717. Crossref
Vila Nova, B.C., Sayão, J.M., Neumann, V.H.M.L., and Kellner, A.W.A. 2014. Redescription of Cearadactylus atrox (Pterosauria, Pterodactyloidea) from the Early Cretaceous Romualdo Formation (Santana Group) of the Araripe Basin, Brazil. Journal of Vertebrate Paleontology 34: 126–134. Crossref
Vullo, R. and Neraudeau, D. 2009. Pterosaur remains from the Cenomanian (Late Cretaceous) paralic deposits of Charentes, western France. Journal of Vertebrate Paleontology 29: 277–282. Crossref
Vullo, R., Allain, R., and Cavin, L. 2016. Convergent evolution of jaws between spinosaurid dinosaurs and pike conger eels. Acta Palaeontologica Polonica 61: 825–828. Crossref
Vullo, R., Buscalioni, A., Marugan-Lobon, J., and Moratalla, J.J. 2009. First pterosaur remains from the Early Cretaceous Lagerstatte of Las Hoyas, Spain: palaeoecological significance. Geological Magazine 146: 931–936. Crossref
Wang, X., Kellner, A.W.A., Zhou, Z., and Campos, D. 2005. Pterosaur diversity and faunal turnover in Cretaceous terrestrial ecosystems in China. Nature 437: 875–879. Crossref
Wang, X., Kellner, A.W.A., Jiang, S., and Cheng. 2012. New toothed flying reptile from Asia: close similarities between early Cretaceous pterosaur faunas from China and Brazil. Naturwissenschaften 99 (4): 249–257. Crossref
Wang, X., Kellner, A.W.A., Jiang, S., Wang, Q., Ma, Y., Paidoula, Y., Cheng, X., Rodrigues, T., Meng, X., Zhang, J., Li, N., and Zhou, Z. 2014. Sexually dimorphic tridimensionally preserved pterosaurs and their eggs from China. Current Biology 24: 1323–1330. Crossref
Wellnhofer P. 1985. Neue pterosaurier aus der Santana-Formation (Apt) der Chapada do Araripe, Brasilien. Palaeontographica. Abteilung A 187: 105–182.
Wellnhofer, P. 1987. New Crested Pterosaurs from the Lower Cretaceous of Brazil. Mitteilungen der Bayerischen Staatssammlung fiir Paleontologie und historische Geologie 27: 175–186.
Wellnhofer, P. and Buffetaut, E. 1999. Pterosaur remains from the Cretaceous of Morocco. Palaeontologie Zeitschrift 73: 133–142. Crossref
Zhou, C.F., Gao, K.Q., Yi H., Xue, J., Li, Q., and Fox, R.C. 2017. Earliest filter-feeding pterosaur from the Jurassic of China and ecological evolution of Pterodactyloidea. Royal Society Open Science 4 (2): 160672. Crossref
Acta Palaeontol. Pol. 69 (1): 73–86, 2024
https://doi.org/10.4202/app.01122.2023