
A new species of the equisetalean plant Equicalastrobus from the Middle Triassic of Argentina
Jano Nehuén Procopio Rodríguez, Josefina Bodnar, and Marisol Beltrán
Procopio Rodríguez, J.N., Bodnar, J., and Beltrán, M. 2024. A new species of the equisetalean plant Equicalastrobus from the Middle Triassic of Argentina. Acta Palaeontologica Polonica 69 (2): 303–313.
We present a morphological-systematic study of new fossil specimens of equisetaleans (horsetails) from the Cortaderita Formation, Sorocayense Group, Middle Triassic, Cuyana Basin, southwest of the San Juan Province, Argentina. In this unit, nine fossiliferous strata with plant remains have been recognized (EF4–12). The fossils studied come from EF4, located in the lower member of the formation, and correspond to impressions-compressions. The fossils are assigned to the family Equisetaceae and the genus Equicalastrobus, being the oldest record of the genus and the first record of this taxon for Argentina and Gondwana. The samples are closed and open strobili composed of peltate sporangiophores, with glabrous hexagonal discs and with an umbo on its external face which projects, acquiring a leaf shape. Some specimens are found in organic connection with leaf sheaths and axis fragments, allowing the whole plant to be reconstructed.
Key words: Sphenophytes, Equisetales, Equisetaceae, Triassic, Cuyana Basin, strobili, sporangiophores.
Jano Nehuén Procopio Rodríguez [procopio.jano@gmail.com; ORCID: https://orcid.org/0009-0000-9823-5702 ], Josefina Bodnar [jbodnar@fcnym.unlp.edu.ar; ORCID: https://orcid.org/0000-0001-7707-396X ], División de Paleobotánica, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Paseo del Bosque s/n, B1900FWA La Plata, Buenos Aires, Argentina. Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
Marisol Beltrán [meduself@hotmail.com; ORCID: https://orcid.org/0000-0003-3562-7201 ], Instituto de Investigaciones en Biodiversidad y Medioambiente (INIBIOMA), CCT CONICET-Patagonia Norte, Av. De Los Pioneros 2350, 8400, San Carlos de Bariloche, Río Negro, Argentina. Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
Received 28 December 2023, accepted 25 April 2024, published online 14 June 2024.
Copyright © 2024 J.N. Procopio Rodríguez et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Equisetales (horsetails) are a group of plants with a worldwide distribution since the Paleozoic (Boureau 1964; Cúneo and Escapa 2006; Taylor et al. 2009; Elgorriaga et al. 2015; Gnaedinger et al. 2020; Villalva et al. 2023). They represent the most diverse and long-lived order within the sphenophytes. Within the families of this group, the Archaeocalamitaceae and Calamitaceae, reached their peak during the Carboniferous and became extinct towards the end of the Permian, while Neocalamitaceae, Schizoneuraceae, Phyllothecaceae, and Equisetaceae began their diversification in the Carboniferous/Permian and continued to the Mesozoic Era (Stewart and Rothwell 1993).
The family Equisetaceae includes the only extant genus Equisetum Linnaeus, 1753 (Boureau 1964), which consists of 15–18 species (Hauke 1963, 1978; Smith et al. 2006; Christenhusz et al. 2019). Fossil representatives, which are found in disarticulated form and do not have all the characters to be assigned to the genus Equisetum, are assigned to the genus Equisetites Sternberg, 1833 (Boureau 1964). Recently, Zhang et al. (2021) proposed that the genera Kraaiaostachys, Viridistachys, and Equicalastrobus should be treated within the family Equisetaceae.
During the Mesozoic, the presence of plants of the family Equisetaceae was reported worldwide, which are represented by rhizomes, leaves, articulated axes, and reproductive structures, the latter being of great importance when posing evolutionary hypotheses of the group (e.g., Taylor et al. 2009). For the Triassic of Argentina, there are numerous citations of impressions and impressions-compressions of axes, referring to the genus Equisetites, but fossil remains of reproductive structures are very scarce, and include, for example Equisetites quindecindentata Menéndez, 1958, Neocalamostachys arrondoi Brea & Artabe, 1999, Equisetostachys sp. (Coturel et al. 2016), Equisetites sp. (Cariglino et al. 2018), and Equisetites lateralis Phillips, 1829 (Gnaedinger et al. 2023).
In this work, we describe strobili assigned to a new fossil species of the genus Equicalastrobus Grauvogel-Stamm & Ash, 1999 (family Equisetaceae) from the Middle Triassic Cortaderita Formation (Sorocayense Group) in the San Juan Province, central-western Argentina, providing new morphological information on the Argentinean Triassic Equisetales.
Institutional abbreviations.—PBSJ, Colección Palaeobotánica of the Instituto y Museo de Ciencias Naturales de San Juan, San Juan, Argentina.
Other abbreviations.—EF, fossiliferous strata.
Nomenclatural acts.—The nomenclatural act included in this work has been registered in Plant Fossil Names Registry (PFNR); urn:lsid:plantfossilnames.org:act:3302 (PFN003302).
Geological setting
The Cuyana (or Cuyo) Basin (western-central Argentina), together with the Ischigualasto-Villa Unión and Marayes-El Carrizal basins, belongs to a series of extensional continental basins that developed during the early Mesozoic on the western edge of Pangaea (Ramos and Kay 1991; López Gamundí 1994). The sediments of the northern part of the Cuyana Basin, in the San Juan Province, are represented by the Sorocayense and Rincón Blanco groups (Spalletti 2001; Barredo 2012). This area is known as Rincón Blanco hemigraben (Barredo and Ramos 2010). According to Barredo and Ramos (2010), the Sorocayense Group accumulated along a passive margin of a hemigraben. On the contrary, the sediments of the Rincón Blanco Group were deposited in the active margin of the hemigraben, which was inferred from the recognition of thicker basal successions and greater volcanic activity (López Gamundí 1994; Barredo and Ramos 1997, 2010).
The Sorocayense Group (Mésigos 1953) crops out in the Barreal-Calingasta depocenter (Stipanicic 1972; López Gamundí 1994), and in the southern part it comprises three formations: Barreal Formation (Middle Triassic), Cortaderita Formation (Middle Triassic), and Cepeda Formation (Upper Triassic) (Groeber and Stipanicic 1953). Except for the Cepeda Formation, the other units of the Sorocayense Group present a continuous series of fossil plant bearing strata.
The Cortaderita Formation is divided into two members according to lithological differences (Bodnar et al. 2019). The lower member (Don Raúl Member) is formed by conglomerates and sandstones in yellowish cross-stratified lenticular bodies, interspersed with grayish massive bentonites and edafized siltstones, muddy sandstones, and greenish bentonites. These fine facies are highly bioturbated with traces of roots and host abundant plant fossil remains (Bodnar 2010; Bodnar et al. 2019). It has been concluded that this member has been deposited in an anastomosing fluvial system of medium-high sinuosity, with amalgamated gravel and sand channels, with well-developed floodplains (Bodnar et al. 2019). The upper member (La Emilia Member) is composed of sandstones with horizontal and cross-laminated, fine to coarse-grained, pink to violet in color, with numerous permineralized trunks, and gray siltstones and claystones with abundant tuffaceous clasts and numerous fossil plant impressions. It was proposed that the La Emilia Member corresponds to high-energy sandy braided fluvial systems (Spalletti 2001; Bodnar et al. 2019). Based on the stratigraphic correlation and the preserved paleofloras, it was inferred that the Don Raúl Member was deposited during the early Ladinian and the La Emilia Member during the late Ladinian (Bodnar et al. 2019).
Material and methods
Plant fossils from the Cortaderita Formation (Middle Triassic, Sorocayense Group) were collected in the La Tinta Creek, 8 km east of the city of Barreal, located in the Southern Precordillera, San Juan Province, west-central Argentina, between W 69°26’12.2” S 31°37’45.8” and W 69°23’52.1” S 31°39’00.1” (Fig. 1A, B).
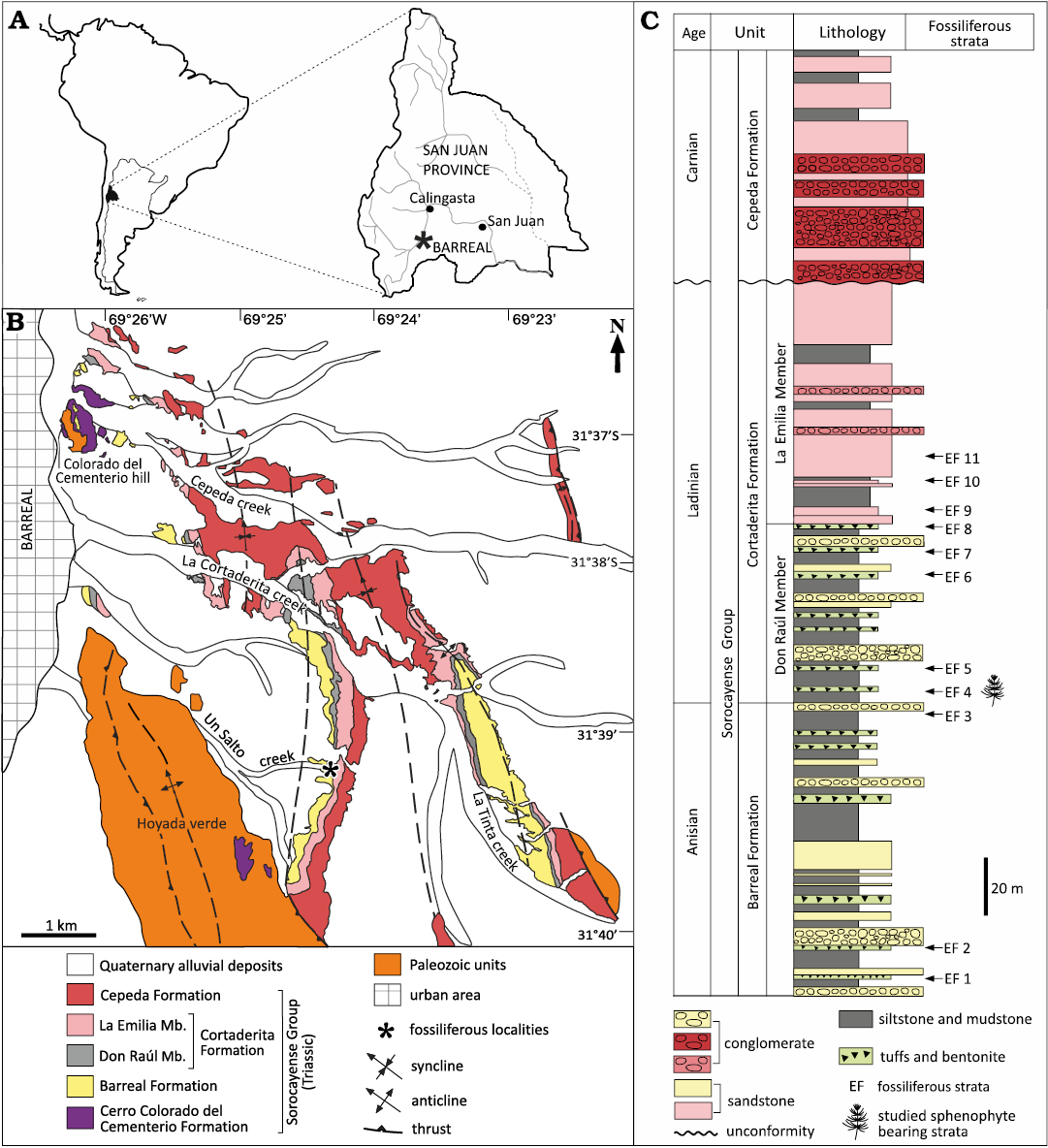
Fig. 1. Geographic and stratigraphic provenance of the studied fossils. A. Geographic location of the Barreal area, San Juan Province, Argentina. B. Geological map of the Barreal depocenter. Modified from Bodnar et al. (2019). Abbreviations: Mb., Member. C. Stratigraphic column of the Sorocayense Group in the Barreal area, with the stratigraphic position of recognized fossiliferous strata (EF). Modified from Bodnar et al. (2018).
In the Cortaderita Formation, five fossiliferous strata (EF) were identified in the Don Raúl Member (EF4–8), and four in the La Emilia Member (EF9–12). The fossil remains studied here come from the stratum EF4 (Fig. 1C) and consist of impressions-compressions of axes, branches, leaves, strobili, and isolated sporangiophores of sphenophytes, which are preserved in tuffaceous and bentonitic gray claystones, siltstones, and silty sandstones (Fig. 1C).
The impressions-compressions were cleaned mechanically with the help of chisels, needles, and a pneumatic pencil. The specimens were studied through a Leica DM 2500 stereo microscope and measured through photographs with ImageJ (Schneider et al. 2012). Photographs of the specimens were taken with Canon EOS Rebel T3i and Canon SX420IS cameras. Details of the samples were photographed with the Leica DC 150 system and the Canon Powershot S40 and Leica DMC2900 cameras.
The descriptions applied the terminology of Font Quer (1982). In the nomenclatural treatment of the fossils, the International Code of Nomenclature for Algae, Fungi, and Plants was followed (Turland et al. 2018). For the reconstruction of the plants, the different parts were linked according to the criteria of Anderson and Anderson (1985). To estimate plant height, the following equations proposed by Niklas (1994) were used:
“Nonwoody and woody” species:
log10H = 1.81 + 0.70(log10D) – 0.13 (log10D)2
“Nonwoody” species:
log10H = 2.51 + 1.41(log10D) + 0.03(log10D)2
where H is the predicted fossil plant height for a known (or specified) axis diameter D (all measurements in m).
The specimens are deposited in the Colección Paleobotánica of the Instituto y Museo de Ciencias Naturales de San Juan, San Juan, Argentina.
Systematic palaeobotany
Class Equisetopsida C. Agardh, 1825
Order Equisetales DC., 1813 ex Bercht. & J. Presl, 1820
Family Equisetaceae Michx ex DC., 1804
Genus Equicalastrobus Grauvogel-Stamm & Ash, 1999
Type species: Equicalastrobus chinleana (Daugherty, 1941) Grauvogel-Stamm & Ash, 1999, Upper Triassic Chinle Formation, Petrified Forest National Park, Arizona, USA.
Equicalastrobus glabratus sp. nov.
Figs. 2–4.
PFNR: urn:lsid:plantfossilnames.org:act:3302 (PFN003302).
Etymology: The specific epithet refers to the absence of trichomes on the margins of the sporangiophore discs.
Type material: Holotype, a complete closed strobilus, PBSJ 1562 (Fig. 2A). Paratype, a complete open strobilus, PBSJ 1539 (Fig. 3A) from the type locality and horizon.
Type locality: Un Salto Creek, Barreal, San Juan Province, Argentina.
Type horizon: EF4, Don Raúl Member, Cortaderita Formation, Sorocayense Group; Ladinian, Middle Triassic.
Material.—42 impressions-compressions of closed and open strobili: PBSJ 1481, 1484, 1487, 1489, 1490, 1493–1495, 1501, 1505, 1506, 1508, 1521, 1522, 1534, 1537, 1539, 1540, 1553, 1555, 1559, 1664, 1576, 1579, 1588, 1589, 1593, 1597, 1598, 1603, 1604, 1605, 1607, 1608, 1609, 1610, 1619, 1621, 1623, 1626, 1736, 1737. All from the type locality and horizon.
Diagnosis.—Cylindrical strobili formed by a main axis on which sporangiophores are arranged in whorls. Sporangiophores consist of hexagonal peltate discs without trichomes (= glabrous). Central umbo, hexagonal, slightly elevated on the external surface of the disc. From the umbo emerges a leaf-shaped projection, with longitudinal striations, ending in a sharp tip directed towards the apex of the strobilus. Base of the strobilus with a leaf sheath protecting it. Branched axes with marked longitudinal ribs, continuous through the node. Robust first-order axes. Thinner second-order axes emerging from the nodes of the first-order axes and bearing leaf-sheaths composed of reduced leaves, fused at the base and free at the apex.
Description.—The studied material corresponds to impressions-compressions of closed and open strobili (Figs. 2A, 3A1, B). The better preserved closed strobilus is cylindrical, compact, 22 mm long by 6 mm in diameter, and is formed by 18 sporangiophores with whorled arrangement, of which only the discs can be seen (Fig. 2A). The discs are peltate, hexagonal, smooth or glabrous, 2.0–2.6 mm in diameter, and present on their external face the mark of a hexagonal umbo, 0.9–1.9 mm in diameter (Fig. 2A, B). The umbo has a leaf-shaped projection, 0.9–1.1 mm wide at its base by 3.3–3.9 mm long, pointed, with longitudinal striae (Fig. 2C, D2), and directed towards the apex of the strobilus (Fig. 2A). At the base of the strobilus, there are leaves 4.7 mm wide by 3.1 mm long fused forming a leaf sheath (Figs. 2A, 4A). The strobilus is found in organic attachment with a small axis fragment 3.2 mm wide by 8.3 mm long, with 5 longitudinal ribs (Figs. 2A, 4A).
The best preserved open strobilus is cylindrical, loose, 38 mm long by 11 mm wide, and is formed by 16 sporangiophore discs with regularly whorled arrangement, with a separation distance 1.7–2.2 mm between whorls. The discs are glabrous, 2.0–3.3 mm in diameter (Figs. 3A, B, 5C). The umbo marks are hexagonal, 0.9–1.7 mm in diameter (Fig. 3A). The umbo has a leaf-shaped projection, with longitudinal striations, 1.4–7.5 mm long by 0.7–1.4 mm wide (Fig. 3A2, B). Other specimens consist of scattered sporangiophore discs 2.0–2.4 mm in diameter (Figs. 2C, D1, 3C), with a hexagonal umbo 0.9–1.4 mm in diameter, with a projection of 1.8–3 mm long by 0.9 mm wide, and sometimes showing traces of probable sporangia up to 1.5 mm long (Fig. 2C).

Fig. 2. Closed strobilus and isolated sporangiophores of the equisetalean plant Equicalastrobus glabratus sp. nov. from Un Salto Creek, Barreal, San Juan Province, Argentina (Ladinian, Middle Triassic). A. PBSJ 1562, holotype, closed strobilus in organic attachment with a leaf sheath and an axis fragment (white arrow). Black arrow indicates the projections of the umbo at the margins of the strobilus. B. PBSJ 1593, sporangiophore discs with a central hexagonal umbo (black arrow). C. PBSJ 1626, laterally arranged sporangiophore with marked umbo projection (black arrow) and possible sporangia (white arrow). D. PBSJ 1604, fragment of a strobilus with two well-preserved whorls with sporangiophores with a central umbo (white arrow) and a marked projection (black arrow) (D1, D2).
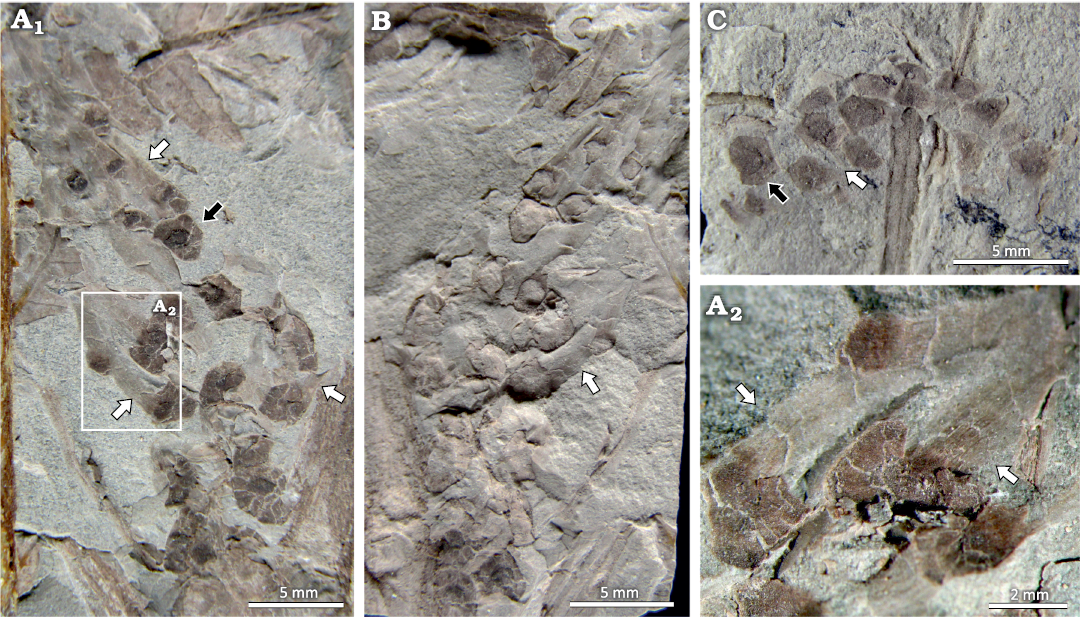
Fig. 3. Open strobili of the equisetalean plant Equicalastrobus glabratus sp. nov. from Un Salto Creek, Barreal, San Juan Province, Argentina (Ladinian, Middle Triassic). A. PBSJ 1539, paratype, open strobilus with well-preserved sporangiophore disc (black arrow) and marked umbo projections (white arrow), with longitudinal striae on projections (white arrow) (A1, A2). B. PBSJ 1540, counterpart of material PBSJ 1539 with longitudinally striated projections (white arrow). C. PBSJ 1736, open strobilus with sporangiophore discs with hexagonal umbo (black arrow) and umbo projections (white arrow).
In the same slabs bearing the strobili, fragmented and entire branched axes were found (Fig. 4C, E–G). The first-order axes are 8–12 mm wide, with well-marked (Fig. 4C) or slightly marked (Fig. 4E) nodes. The surface of the internodes shows distinct 4–5 longitudinal ribs, separated by 0.8–1.1 mm, which are continuous as they pass through the node (Fig. 4C, G). No leaf sheaths were found associated with the first-order axes. The second-order axes are 2–4 mm wide, with marked nodes and internodes 9 mm long (Fig. 4B, E). The surface of the internodes shows four distinct longitudinal ribs, separated by 0.5–0.6 mm (Fig. 4A, B, E). Leaf-sheaths are 4 mm long by 4 mm wide and are preserved attached to the nodes, with eight poorly preserved leaves, fused at the base and free at the apex (Fig. 4B). A leaf sheath was also found attached to a 5 mm wide node, which has 15 leaves, each one 2.6 mm long by 0.7 mm wide, fused at the base and free at the apex (Fig. 4D).
Remarks.—The fossils here studied are assigned to Equicalastrobus because they share the main diagnostic characteristics with that taxon, i.e., cylindrical strobilus bearing peltate sporangiophores consisting of a stalk and a hexagonal disk with a single forward-directed leaf-like umbo narrow projection in its outer surface (Grauvogel-Stamm and Ash 1999). The presence of projections on the umbo of the sporangiophores allows the specimens here studied to be differentiated from the genus Equisetostachys Halle, 1908 (Fig. 5D1, D3, F, K), from Equisetites Sternberg, 1833, and Neocalamostachys Kon’no, 1962, present in the Triassic of Argentina and other regions. Other genera which are characterized by projections on the umbo are Kraaiostachys Anderson & Anderson, 2018, Viridistachys Anderson & Anderson, 2018, and Neocalamites meriani (Brongniart, 1828) Halle, 1908, but differ from the Cortaderita fossils due to the first one presents sub-circular sporangiophore discs with an attenuated conical umbo, the second one possesses hexagonal sporangiophore discs with a wider projection that covers the entire diameter of the disc umbo (Fig. 5H–J), while the last one is larger with hexagonal to polygonal sporangiophores with spiny appendages. Pott et al. (2008) described the reproductive structure associated with Neocalamites meriani, but it is not found in organic connection. The strobili assigned to Neocalamites broadly correspond to the genus Neocalamostachys Kon’no, 1962, and do not present spiny appendages, so the systematic assignment of said strobili should be reconsidered. Unfortunately, there is no more information about the spiny appendages of the specimen studied by Pott et al. (2008) for comparison. Until now, two species were described within Equicalastrobus, i.e., E. chinleana (Daugherty, 1941) Grauvogel-Stamm & Ash, 1999, from the Upper Triassic of the USA, and E. pusillus Zhang et al., 2021, from the Upper Triassic of China. Equicalastrobus glabratus sp. nov. is differentiated from E. chinleana (Daugherty, 1941) Grauvogel-Stamm & Ash, 1999, by the smaller strobili, smaller sporangiophores, and shorter and narrower umbo projections (Fig. 5E, F). On the other hand, the new species differs from E. pusillus by the markedly longer umbo projections (Fig. 5D1, D2, F, G). Equicalastrobus glabratus sp. nov. also is unlike the previously mentioned species by the absence of trichomes on the margins of the discs (Fig. 5E–G).
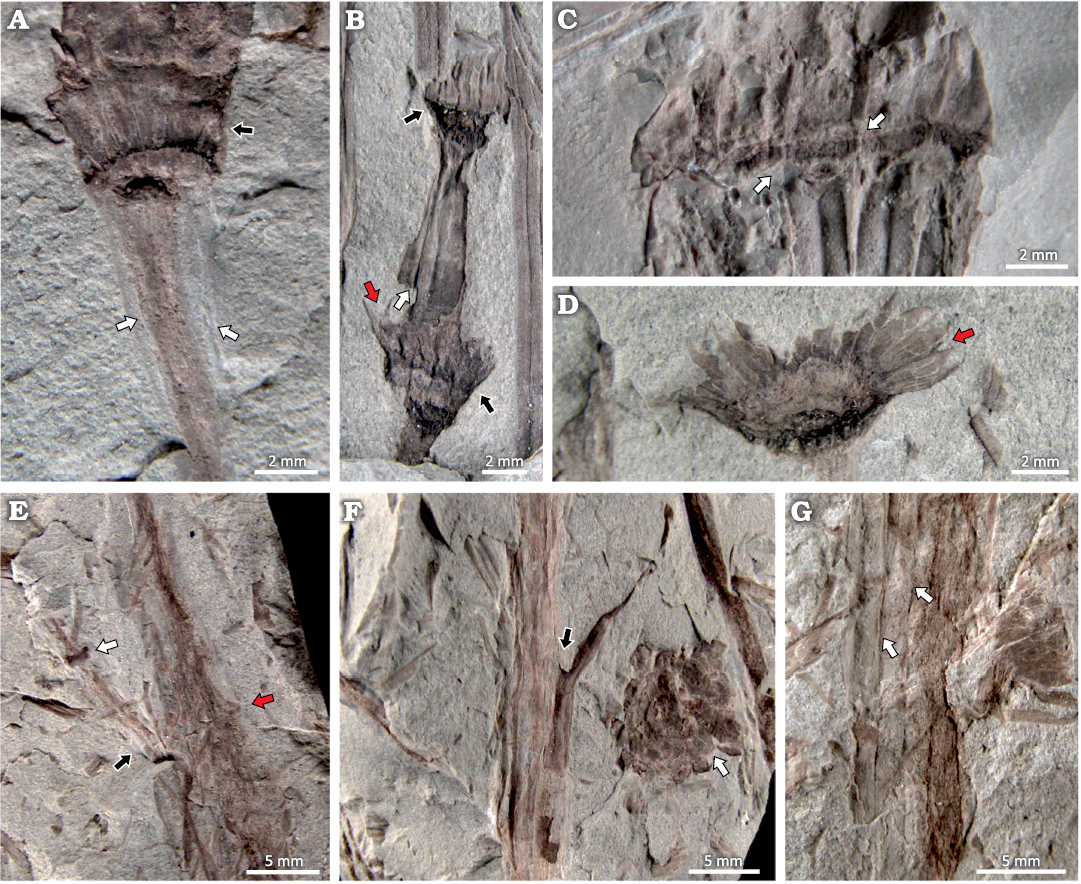
Fig. 4. Axis and leaf sheaths of the equisetalean plant Equicalastrobus glabratus sp. nov. from Un Salto Creek, Barreal, San Juan Province, Argentina (Ladinian, Middle Triassic). A. PBSJ 1562, detail of the axis with longitudinal ribs (white arrow) and a leaf sheath (black arrow), attached to the strobilus. B. PBSJ 1503, axis fragment with two nodes with leaf sheaths (black arrow), with leaves free at the apex (red arrow) and an internode with longitudinal ribs (white arrow). C. PBSJ 1481, detail of the ribs (white arrow) as they pass through the node. D. PBSJ 1506, detail of leaf sheath, with leaves free at the apex (red arrow). E. PBSJ 1598, first-order axis with a second-order branch (black arrow), a few marked nodes of the first-order axis (red arrow), and node of the second-order axis (white arrow). F. PBSJ 1487, first-order axis with a second-order branch (black arrow) next to a strobilus (white arrow). G. PBSJ 1607, detail of the longitudinal ribs (white arrow).
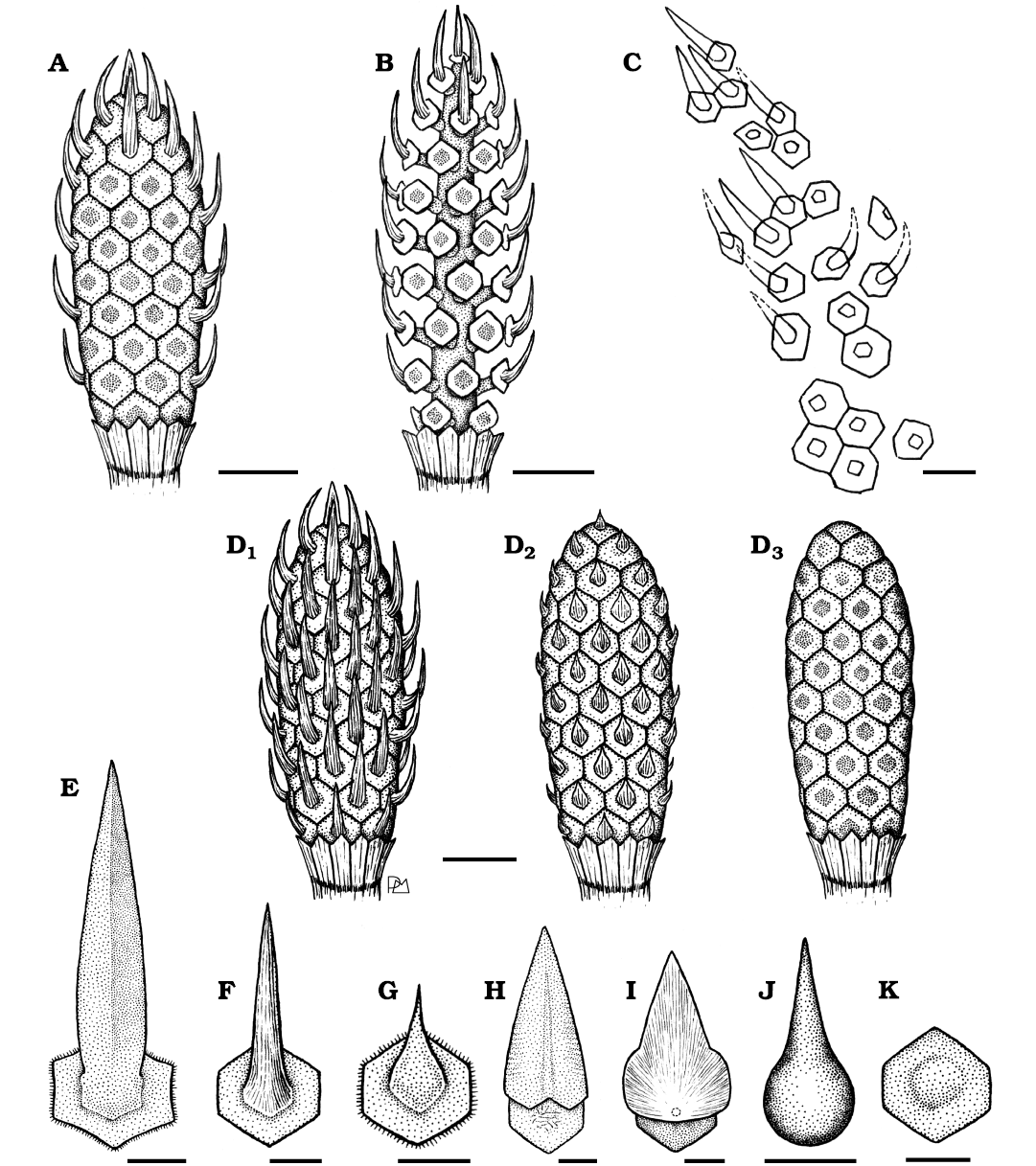
Fig. 5. Reconstruction of the equisetalean plant Equicalastrobus glabratus sp. nov. from Un Salto Creek, Barreal, San Juan Province, Argentina (Ladinian, Middle Triassic), and comparisons with other equisetalean strobili and isolated sporangiophores. Illustration of the closed strobilus (A, PBSJ 1562) and open strobilus (B, PBSJ 1539). C. Sketching of PBSJ 1539 where incomplete parts are interpreted with dotted lines. D. Reconstruction of strobilus of Equicalastrobus glabratus sp. nov. (D1) and comparison with a hypothetical reconstruction of Equicalastrobus pusillus Zhang et al., 2021 (D2) and with an Equisetostachys sp. strobilus (D3). E–K. Comparison of sporangiophores of species of different genera showing projections of the umbo and a disc of Equisetostachys. E. Equicalastrobus chinleana (Daugherty, 1941) Grauvogel-Stamm & Ash, 1999. F. Equicalastrobus glabratus sp. nov. G. Equicalastrobus pusillus Zhang et al., 2021. H. Viridistachys gypsensis Anderson & Anderson, 2018. I. Viridistachys moltenensis Anderson & Anderson, 2018. J. Kraaiostachys plaatkoppiensis Anderson & Anderson, 2018. K. Equisetostachys Halle, 1908. Scale bars 5 mm (A, B, D), 3 mm (C, E), 1 mm (F–K).
Regarding the axes and leaf sheaths, Equicalastrobus glabratus sp. nov. differs from the species of Equisetites and Equisetum by the presence of longitudinal ribs passing through the node. On the other hand, the new species is distinct from Schizoneura Schimper & Mougeot, 1844, by the leaf-sheath morphology, since in this genus they are larger and have laterally fused and flattened leaves, which are arranged in two opposite lobes (Boureau 1964), while the leaf sheaths in Equicalastrobus glabratus sp. nov. are characterized by reduced leaves fused at the base and free at the apex, and not arranged in lobes. Finally, Neocalamites presents robust branched axes, with Asterophyllites/Annularia-type unfused leaves (Boureau 1964), ruling out that the here studied axes can be assigned to that genus.
Stratigraphic and geographic range.—Ladinian, Middle Triassic; San Juan Province, Argentina.
Reconstruction of the whole plant
Anderson and Anderson (1985) proposed a scale to determine the degree of certainty in linking different fossil organs for the reconstruction of the entire plant. In this scale, it is proposed that the criteria for relating two or more organs are the following (from most reliable to least reliable): organic attachment, morphological similarity, kindred reinforcement, and mutual occurrence. The strobili of Equicalastrobus glabratus sp. nov. were found in organic attachment with the leaf sheaths and axis fragments. In turn, for the reproductive structures, a morphological similarity is established between dispersed sporangiophores and the complete closed and open strobili from different slabs. It is reconstructed as a herbaceous form with an estimated height of 48–82 cm (applying the “nonwoody” formula of Niklas 1994) or 60–97 cm (applying the “nonwoody and woody” formula of Niklas 1994), with branched axes, leaves fused into a leaf sheath and cylindrical strobili with peltate sporangiophores, with hexagonal discs that have a hexagonal umbo on their external face that projects forming an appendage similar to a leaf (Figs. 5A, B, D1, 6).
The scarce material in organic attachment does not allow us to identify whether the strobili are arranged in the first-order axes, in the second-order axes, or in both. In this reconstruction they are placed on the second-order axes, but the possibility that they are arranged in another way is not ruled out (Fig. 6).
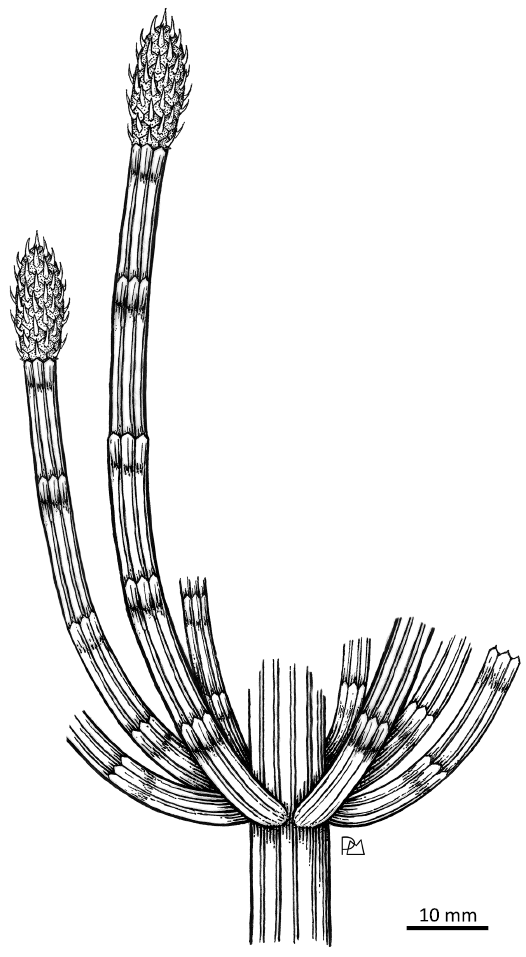
Fig. 6. The equisetalean plant Equicalastrobus glabratus sp. nov. from Un Salto Creek, Barreal, San Juan Province, Argentina (Ladinian, Middle Triassic), reconstruction of a part of the whole plant.
Discussion
Initially, the type material of Equicalastrobus was assigned by Daugherty (1941) to the lycopodialean strobilus Lycostrobus chinleana Daugherty, 1941. Years later, several authors reviewed the type material (Chaloner and Boureau 1967; Miller 1968; Bock 1969; Taylor 1981; Taylor and Taylor 1993; Retallack 1997) suggesting its doubtful systematic assignment, and indicating possible assignments to other lycophytes or conifers (Zhang et al. 2021). Stewart and Rothwell (1993) discussed its systematic allocation and related it to the equisetaleans. Grauvogel-Stamm and Ash (1999) reviewed the type material, confirmed its assignment to the order Equisetales, and erected the genus Equicalastrobus to include those strobili composed of peltate sporangiophores, with hexagonal discs with marginal trichomes, that present a hexagonal/rhomboidal umbo that is projected forming a structure similar to a “leaf”. These authors described the detailed morphology of the specimens and hypothesized that Equicalastrobus chinleana represents a transitional stage between Calamitaceae and Equisetaceae. On the other hand, Zhang et al. (2021) described the species of Equicalastrobus pusillus, which was found on the same slab along with Schizoneura; however, since they are not in an organic attachment, the authors considered that they should not be assigned to the same species. Zhang et al. (2021) also carried out a clustering analysis of the reproductive organs of fossil and extant species, resulting in Equicalastrobus being more closely related to the family Equisetaceae than to the Calamitaceae.
The strobili of Equicalastrobus glabratus sp. nov. were found in the same fossiliferous stratum next to articulated axes with whorled reduced leaves, fused at the base and free at the apex forming a sheath (Fig. 4D). The organic attachment between strobili and axis fragments indicates that these correspond to the vegetative part of Equicalastrobus, which until now was unknown. These axes and leaf sheaths are very similar to the axes of Equisetites and Equisetum, however, they differ from these by presenting longitudinal ribs parallel passing through the node. The fossil remains of Equicalastrobus found in the Cortaderita Formation represent the oldest record of the genus since the two species described so far come from the Upper Triassic.
The reproductive characters of Equicalastrobus glabratus sp. nov. indicate a close affiliation to the families Equisetaceae and Neocalamitaceae (= Apocalamitaceae), as both are characterized by strobili with hexagonal sporangiophores and without bracts, similar to those of the new species, but differ in the umbo projection which is absent in those families (De Candolle 1804; Boureau 1964; Stewart and Rothwell 1993; Simpson 2005). On the other hand, some vegetative characters of Equicalastrobus glabratus sp. nov. are shared by the family Neocalamitaceae (continuous ribs passing through the node) and by the family Equisetaceae (leaf sheaths with reduced leaves). According to Elgorriaga et al. (2018), the family Neocalamitaceae and Equisetaceae are two lineages grouped in a major clade. In contrast, the analysis of Zhang et al. (2021) proposed a major clade including the genera assigned to both Equisetaceae and Neocalamitaceae, but each family are not monophyletic (see Zhang et al. 2021: fig. 5). Based on the information from the both analyses, it could be interpreted that Equicalastrobus would be located basally in the family Equisetaceae but with characters, such as continuous ribs through the node, that are shared with the Neocalamitaceae. Unfortunately, few materials of this genus are known, so a deeper study of reproductive and vegetative structures from the Triassic, such as those of older ages, could provide more information about the lineage origins.
The assemblage of continental masses into a single supercontinent, Pangaea (Wing and Sues 1992), made up of the Laurasic and Gondwanan Phytogeographical Kingdoms (Artabe et al. 2003), led to widespread dispersal of many plant species during the Triassic (Artabe et al. 2003). One of the groups that achieved a cosmopolitan distribution was the family Equisetaceae, which comprises species of the only living genus, Equisetum, but it also includes extinct fossil genera such as Equisetites, Equisetostachys, Viridistachys, Kraaiostachys, and Equicalastrobus (Anderson and Anderson 2018; Zhang et al. 2021). Until now occurrences of Equicalastrobus were known from the USA and China (Grauvogel-Stamm and Ash 1999; Zhang et al. 2021) (Laurasian Phytogeographical Kingdom). In this work, the first record of the genus for Argentina and for the Gondwanan Kingdom indicates a cosmopolitan distribution of the genus Equicalastrobus and the family Equisetaceae.
Conclusions
The finding of impressions-compressions of reproductive structures assignable to the Equisetales represents the first fertile record of this order from the Cortaderita Formation. On the other hand, the discovery of the genus Equicalastrobus represents the oldest occurrence of the taxon worldwide and the first record of the genus for Argentina and Gondwana. These structures are found in organic attachment and close association with axes and leaf sheaths allowing the reconstruction of a whole plant of Equicalastrobus, until now unknown. The whole plant would have been herbaceous, branched, with axes with distinct nodes and internodes, whorled leaves fused at the base and free at the apex forming a leaf sheath.
Acknowledgements
We thank Eliana P. Coturel (Museo de La Plata, La Plata, Argentina), Adolfina Savoretti, (Centro Austral de Investigaciones Científicas del Consejo Nacional de Investigaciones Científicas y Técnicas, Ushuaia, Argentina), Carina Colombi and Juan M. Drovandi (both Centro de Investigaciones de la Geósfera y Biósfera, Consejo Nacional de Investigaciones Científicas y Técnicas-Universidad Nacional de San Juan, San Juan, Argentina), and Ari Iglesias (Instituto de Investigaciones en Biodiversidad y Medioambiente, Consejo Nacional de Investigaciones Científicas y Técnicas-Universidad Nacional del Comahue, Bariloche, Argentina), for their help in the field work. We also expressed acknowledgment to Eliana P. Coturel for her help in the discussion and interpretation specimens, Andrés Elgorriaga (University of Kansas, Lawrence, USA) for contributing with the systematic assignment, and to Paula Marcantoni (Museo de La Plata, La Plata, Argentina) for the illustrations. Finally, would like to thank to the reviewers, Evelyn Kustatcher (Museum of Nature South Tyrol, Bolzano, Italy), Christian Pott (Museum of Natural History and Planetarium, Munster, Germany), and Grzegorz Pacyna (Institute of Botany, Department of Taxonomy, Phytogeography and Palaeobotany, Gronostajowa, Kraków, Poland), whose comments contributed to the improvement of the work. This contribution was supported by ANPCyT (PICT2751) and UNLP (N807, N931).
References
Agardh, C.A. 1825. Equisetopsida. In: Aphorismi Botanici, 7. Literis Berlingianis, Lund.
Anderson, J.M. and Anderson, H.M. 1985. Palaeoflora of Southern Africa. Prodromus of South African Megafloras: Devonian to Lower Cretaceous. 423 pp. A.A. Balkema, Rotterdam.
Anderson, H.M., and Anderson. J.M. 2018. Molteno sphenophytes: Late Triassic biodiversity in southern Africa. Palaeontologia africana 53 (SI): 1–391
Artabe, A.E., Morel, E.M., and Spalletti, L.A. 2003. Caracterización de las provincias fitogeográficas triásicas del Gondwana extratropical. Ameghiniana 40: 387–405.
Barredo, S.P. 2012. Geodynamic and tectonostratigraphic study of a continental rift: the Triassic Cuyana Basin, Argentina. In: E. Sharkov (ed.), Tectonics—Recent Advances, 99–130. InTech, London.
Barredo, S.P. and Ramos, V.A. 1997. Los depósitos de Rincón Blanco (Precordillera de San Juan): un rift triásico. II Jornadas sobre Geología de Precordillera (San Juan), Actas 1: 130–135.
Barredo, S.P. and Ramos, V.A. 2010. Características tectónicas y tectosedimentarias del hemigraben Rincón Blanco, Cuenca Cuyana: una síntesis. Revista de la Asociación Geológica Argentina 66: 133–145.
Berchtold, F. and Presl, J.S. 1820. O Přirozenosti Rostlin. 322 pp. Jos Krause, Praga.
Bodnar, J. 2010. La paleoflora triásica de la Formación Cortaderita en la quebrada homónima, Cuenca de Barreal – Calingasta, provincia de San Juan, Argentina. 283 pp. Ph.D. Thesis, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata [available from http://sedici.unlp.edu.ar/handle/10915/4308]
Bodnar, J., Drovandi, J.M., Morel, E.M., and Ganuza, D.G. 2018. Middle Triassic dipterid ferns from west-central Argentina and their relationship to palaeoclimatic changes. Acta Palaeontologica Polonica 63: 397–416. Crossref
Bodnar, J., Iglesias, A., Colombi, C.E., and Drovandi, J.M. 2019. Stratigraphical, sedimentological and paleofloristic characterization of Sorocayense Group (Triassic) in Barreal Area, San Juan Province, Argentina. Andean Geology 46: 567–603. Crossref
Bock, W. 1969. The American Triassic flora and global distribution. Geological Center Series 3/4: 1–406.
Boureau, E. 1964. Traité de Paléobotanique 3, Sphenophyta, Noeggerathiophyta. 344 pp. Masson et Cie, Paris.
Brea, M. and Artabe, A.E. 1999. Apocalamitaceae (Sphenophyta) triásicas de la Formación Paramillo, Agua de la Zorra, provincia de Mendoza, Argentina. Ameghiniana 36: 389–400.
Brongniart, A. 1828. Prodrome d`une histoire des végétaux fossiles. VIII. 223 pp. Leurault, París. Crossref
Cariglino, B., Monti, M., and Zavattieri, A.M. 2018. A Middle Triassic macroflora from southwestern Gondwana (Mendoza, Argentina) with typical Northern Hemisphere elements: Biostratigraphic, palaeogeographic and palaeoenvironmental implications. Review of Palaeobotany and Palynology 257: 1–18. Crossref
Chaloner, W.G. and Boureau, E. 1967. Lycophyta. In: E. Boureau (ed.), Traité de Paleo-botanique. Vol. 2, 435–802. Masson et Cie., Paris.
Christenhusz, M.J., Bangiolo, L., Chase, M.W., Fay, M.F., Husby, C., Witkus, M., and Viruel, J. 2019. Phylogenetics, classification and typification of extant horsetails (Equisetum, Equisetaceae). Botanical Journal of the Linnean Society 189: 311–352. Crossref
Cúneo, N.R. and Escapa, I. 2006. The Equisetalean Genus Cruciaetheca nov. from the Lower Permian of Patagonia, Argentina. International Journal of Plant Sciences 167: 167–177. Crossref
Coturel, E.P., Morel, E.M., and Ganuza, D. 2016. Lycopodiopsids and equisetopsids from the Triassic of Quebrada de los Fósiles formation, San Rafael Basin, Argentina. Geobios 49: 167–176. Crossref
Daugherty, L.H. 1941. The Upper Triassic Flora of Arizona. Vol. 526. 108 pp. Carnegie Institution of Washington Publication, Washington DC.
De Candolle, A.P. 1804. Essai sur les propriétés médicales des plantes, comparées aves leurs formes extérieures et leur classification naturelle. 148 pp. Didot Jeune, Paris.
De Candolle, A.P. 1813. Catalogus plantarum horti botanici monspeliensis. 155 pp. Martel, Paris.
Elgorriaga, A., Escapa, I.H., Bomfleur, B., Cúneo, N.R., and Ottone, E.G. 2015. Reconstruction and phylogenetic significance of a new Equisetum Linnaeus species from the Lower Jurassic of Cerro Bayo (Chubut Province, Argentina). Ameghiniana. 52: 135–152. Crossref
Elgorriaga, A., Escapa, I.H., Rothwell, G.W., Tomescu, A.M., and Cúneo, N.R. 2018. Origin of Equisetum: Evolution of horsetails (Equisetales) within the major euphyllophyte clade Sphenopsida. American Journal of Botany 105: 1286–1303. Crossref
Font Quer, P. 1982. Diccionario de Botánica. 8th Reprint. 1244 pp. Editorial Labor, Barcelona.
Gnaedinger, S., Cariglino, B., Zavattieri, A.M., Monti, M., and Gutiérrez, P.R. 2020. Neoarthropitys gondwanaensis gen. nov. et sp. nov. from the Middle Triassic of Gondwana: an intermediate stage in the anatomical trend of Equisetalean stems. Review of Palaeobotany and Palynology 282: 104298. Crossref
Gnaedinger, S., Villalva, A.S., and Zavattieri, A.M. 2023. Triassic Equisetites lateralis Phillips with strobilus in organic connection from Patagonia of Argentina and endophytic oviposition insect scars. Review of Palaeobotany and Palynology 317: 104964. Crossref
Grauvogel-Stamm, L. and Ash, S.R. 1999. “Lycostrobus” chinleana, an equisetalean cone from the Upper Triassic of the southwestern United States and its phylogenetic implications. American Journal of Botany 86: 1391–1405. Crossref
Groeber, P. and Stipanicic, P.N. 1953. Triásico. In: P. Groeber, P.N. Stipanicic, and A.R.G. Mingramm (eds.), Mesozoico. Geografía de la República Argentina. Sociedad Argentina de Estudios Geográficos GAEA 2 (1): 13–131.
Halle, T.G. 1908. Zur Kenntnis der Mesozoischen equisetales Schwedens. Kungliska Svenska Vetenskapsademiens Handlingar 43: 3–42.
Hauke, R.L. 1963. A taxonomic monograph of the genus Equisetum, subgenus Hippochaete. Beih. Nova Hedwigia 8: 1–123. Crossref
Hauke, R.L. 1978. A taxonomic monograph of Equisetum subgenus Equisetum. Nova Hedwigia 30: 385–455. Crossref
Kon’no, E. 1962. Some species of Neocalamites and Equisetites in Japan and Korea. Science Reports, Tohoku University 5: 21–47.
Linnaeus C.V. 1753. Species plantarum, exhibentes plantas rite cognitas, ad genera relatas, cum differentiis specificis, nominibus trivialibus, synonymis selectis, locis natalibus, secundum systema sexuale digestas, Vol. 2, 561–1200. Laurentii Salvii, Stockholm. Crossref
López Gamundí, O. 1994. Facies distribution in an asymmetric half-graben: the northern Cuyo Basin (Triassic), western Argentina. In: XIV International Sedimentological Congress. Abstracts, 6–7. Petrobrás, Recife.
Miller, C.N. 1968. Lepidophytic affinities of genus Chinlea and Osmundites walkeri. American Journal of Botany 55:109–115. Crossref
Menéndez, C.A. 1958. Equisetites quimdecimdentata sp. nov. del Triásico Superior de Hilario, San Juan. Revista de la Asociación Geológica Argentina 13: 5–14.
Mésigos, M.G. 1953. El Paleozoico Superior de Barreal y su continuación austral, Sierra de Barreal, Provincia de San Juan. Revista de la Asociación Geológica Argentina 8: 65–109.
Niklas, K.J. 1994. Predicting the height of fossil plant remains: an allometric approach to an old problem. American Journal of Botany 81: 1235–1242. Crossref
Phillips, J. 1829. Illustrations of the Geology of Yorkshire: or, a Description of the Strata and Organic Remains of the Yorkshire Coast. xvi+ 192 pp. J. Murray, London. Crossref
Pott, C., Kerp, H., and Krings, M. 2008. Sphenophytes from the Carnian (Upper Triassic) of Lunz am See (Lower Austria). Jahrbuch der Geologischen Bundesanstalt 148: 183–199.
Ramos, V. and Kay, S. 1991. Triassic rifting and associated basalts in the Cuyo basin, central Argentina. In: R.S. Harmon and C.W. Rapela (eds.), Andean Magmatism and its Tectonic Setting. Geological Society of America, Special Paper 265: 79–91. Crossref
Retallack, G.J. 1997. Earliest Triassic origin of Isoetes and quillwort evolutionary radiation. Journal of Paleontology 71: 500–521. Crossref
Schimper, W.P. and Mougeot, A. 1844. Monographie des Plantes fossiles du Grés Bigarré de la Chaine des Vosges. 83 pp. G. Engelmann, Leipzig, Crossref
Schneider, C.A., Rasband, W., and Eliceiri, K.W. 2012. NIH Image to ImageJ: 25 years of image analysis. Nature Methods 9: 671–675. Crossref
Simpson, M.G. 2005. Plant Systematics. First Edition. 608 pp. Academic Press, Amsterdam. Crossref
Smith, A.R., Pryer, K.M., Schuettpelz, E., Korall, P., Schneider, H., and Wolf, P.G. 2006. A classification for extant ferns. Taxon 55: 705–731.
Spalletti, L.A. 2001. Modelo de sedimentación fluvial y lacustre en el margen pasivo de un hemigraben: el Triásico de la Precordillera occidental de San Juan, República Argentina. Revista de la Asociación Geológica Argentina 56: 189–210.
Sternberg, W.P. 1833. Versuch einer geognostichen botanischen. Darstellung der flora der Vorwelt 2: 1–80.
Stewart, W.R. and Rothwell, G.W. 1993. Paleobotany and the Evolution of Plants. 521 pp. Cambridge University Press, Cambridge.
Stipanicic, P.N. 1972. Cuenca triásica de Barreal. In: A.F. Leanza (ed.), Geología Regional Argentina, 537–566. Academia Nacional de Ciencias, Córdoba.
Taylor, T.N. 1981. Paleobotany. an Introduction to Fossil Plant Biology. 589 pp. McGraw-Hill, New York.
Taylor, T.N. and Taylor, E.L. 1993. The Biology and Evolution of Fossil Plants. 982 pp. Prentice Hall, Englewood Cliffs.
Taylor, E.L., Taylor, T. N., and Krings, M. 2009. Paleobotany: the Biology and Evolution of Fossil Plants. 1088 pp. Academic Press, Amsterdam.
Turland, N.J., Wiersema, J.H., Barrie, F.R., Greuter, W., Hawksworth, D.L., Herendeen, P.S., Knapp, S., Kusber, W.-H., Li, D.-Z., Marhold, K., May, T.W., McNeill, J., Monro, A.M., Prado, J., Price, M.J., and Smith, G.F. (eds.) 2018. International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017. Regnum Vegetabile 159, Koeltz Botanical Books, Glashütten. Crossref
Villalva, A.S., Gnaedinger S., and Zavattieri, A.M. 2023. Systematic and organ relationships of Neocalamites (Halle) Vladimirovicz, and Nododendron (Artabe and Zamuner) emend. from the Triassic of Patagonia. Palaeobiogeographic, palaeoenvironments and palaeoecology considerations. Review of Palaeobotany and Palynology 316: 104939. Crossref
Wing, S.L. and Sues, H.-D. 1992. Mesozoic and Early Cenozoic terrestrial ecosystems. In: A.K. Behrensmeyer, J.D. Damuth, W.A. DiMichele, R. Potts, H.-D. Sues, and S.L. Wing (eds.), Terrestrial Ecosystems Through Time, 327–416. The University of Chicago Press, Chicago.
Zhang, L., Wang, Y., Chen, H Y., Han, L., Zhang, Y.X., Li, W. J., and Yan, D.F. 2021. New fossil material of Equicalastrobus (Equisetales) and associated leaves from the Late Triassic of Baojishan basin, Gansu Province, China. Historical Biology 33: 1522–1533. Crossref
Acta Palaeontol. Pol. 69 (2): 303–313, 2024
https://doi.org/10.4202/app.01130.2023