
How not to disappear completely: new Stereospondyli fossils from the Rhaetian, Upper Triassic of Bonenburg, North Rhine-Westphalia and their implications for the Late Triassic extinction of Stereospondyli
ANDREA PRINO, FLORIAN WITZMANN, ACHIM H. SCHWERMANN, P. MARTIN SANDER, LAURENT GARBAY, and DOROTA KONIETZKO-MEIER
Prino, A., Witzmann, F., Schwermann, A.H., Sander, P.M., Garbay, L., and Konietzko-Meier D. 2024. How not to disappear completely: new Stereospondyli fossils from the Rhaetian, Upper Triassic of Bonenburg, North Rhine-Westphalia and their implications for the Late Triassic extinction of Stereospondyli. Acta Palaeontologica Polonica 69 (4): 687–712.
Temnospondyli appeared in the early Carboniferous, became extinct in the Early Cretaceous and reached high diversity especially during the Permian and Triassic. After the end-Permian mass extinction, almost only Stereospondyli survived (with the exception of the dissorophoid Micropholis). This clade radiated and gave rise to several successful groups: Plagiosauroidea, Trematosauroidea, Metoposauroidea, Capitosauria, and Brachyopoidea. While Brachyopoidea survived into the Early Cretaceous, the other groups were thought to have gradually disappeared during the Late Triassic, going extinct before the Rhaetian. This hypothesis was supported by the lack of unambiguously dated Rhaetian localities with Stereospondyli fossils. This gap was filled by the discovery of the Bonenburg clay pit in North Rhine-Westphalia (Germany). This locality has been unequivocally dated to the late middle Rhaetian and had yielded a temnospondyl fossil assigned to Capitosauria. Here we describe further Stereospondyli fossils from Bonenburg, including diagnostic bones such as a dentary, a pterygoid, a parietal, and eight other bones in different states of preservation. These fossils belong to at least two taxa of Capitosauria and one taxon of Plagiosauridae and represent the geologically youngest remains of both clades. The specimens are described morphologically, and for the long bones, a clavicle fragment, and an unidentified dermal bone, histological analysis was used to further confirm the morphological analysis. These results shed light on the extinction of two major groups of Stereospondyli, documenting that some non-brachyopoid temnospondyls survived until the end of the Triassic.
Key words: Temnospondyli, Capitosauroidea, Plagiosaurinae, bone histology, end-Triassic extinction, Rhaetian, Bonenburg.
Andrea Prino [andrea.prino@hu-berlin.de; ORCID: https://orcid.org/0009-0006-4620-2224 ], Comparative Zoology, Institute of Biology, Humboldt University of Berlin, Philippstrasse 12/13, 10115 Berlin, Germany. Museum für Naturkunde, Leibniz-Institut für Evolutions- und Biodiversitätsforschung, Invalidenstraße 43, 10115 Berlin, Germany. Bonn Institute of Organismic Biology (BIOB-V), Section Paleontology, University of Bonn, Nussallee 8, Bonn 53115, Germany.
Florian Witzmann [florian.witzmann@mfn.berlin; ORCID: https://orcid.org/0000-0002-0731-3259 ], Museum für Naturkunde, Leibniz-Institut für Evolutions- und Biodiversitätsforschung, Invalidenstraße 43, 10115 Berlin, Germany.
Achim H. Schwermann [achim.schwermann@lwl.org; ORCID: https://orcid.org/0000-0001-8928-8773 ], LWL-Museum für Naturkunde, Westfälisches Landesmuseum mit Planetarium, 48161 Münster, Germany.
P. Martin Sander [m.sander@uni-bonn.de; ORCID: https://orcid.org/0000-0003-4981-4307 ], Bonn Institute of Organismic Biology (BIOB-V), Section Paleontology, University of Bonn, Nussallee 8, Bonn 53115, Germany.
Laurent Garbay [garblaur@gmail.com; ORCID: https://orcid.org/0000-0001-6048-2468 ], Natural History Museum Luxembourg, Department of Paleontology, 25 rue Münster, 2160 Luxembourg City, Luxembourg.
Dorota Konietzko-Meier[dorota.konietzko-meier@smns-bw.de; ORCID: https://orcid.org/0000-0003-4932-7402 ], Bonn Institute of Organismic Biology (BIOB-V), Section Paleontology, University of Bonn, Nussallee 8, Bonn 53115, Germany; Stuttgart State Museum of Natural History, Rosenstein 1, 70191 Stuttgart, Germany.
Received 26 February 2024, accepted 2 November 2024, published online 23 December 2024.
Copyright © 2024 A. Prino et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Temnospondyli Zittel, 1888, were first described by Jaeger (1824, 1828) at the beginning of the 19th century and comprise a clade of ecologically highly diverse early tetrapods that occupied a variety of niches in aquatic, semiterrestrial and terrestrial habitats. Today, about 200 genera and 300 species of temnospondyls have been identified from all continents (Schoch 2013). The first representatives of the clade appeared in the early Carboniferous of Europe (Milner and Sequeira 1994; Werneburg et al. 2019), but they highly diversified only in the Permian, reaching a peak in the Triassic (Milner 1990). The most recent specimen found in the fossil record is the Australian Koolasuchus cleelandi, indicating the survival of the temnospondyls into the Early Cretaceous (Warren et al. 1997). Over the course of their long history, temnospondyls attained a high degree of diversity in terms of morphology, adaptations, and the number of species. The largest temnospondyls were up to 6 m long (Schoch and Milner 2000), while the smallest reached only a few centimetres in length (Boy 1972). The majority of temnospondyls have been found in freshwater sediments deposited in swamps, lakes, and rivers (Schoch and Milner 2000). However, some taxa are thought to have lived in more terrestrial environments (Pawley and Warren 2005), while others apparently adapted to marine environments, such as the trematosaurids (Kear et al. 2016).
During the Permian, the diversity of
temnospondyls increased on all continents (e.g., Pawley
and Warren 2006; Steyer et al. 2006; Marsicano
et al. 2021). After the end-Permian mass extinction, the most
severe ever (Erwin et al. 2002; Benton
2015), the only group of temnospondyls that survived (with the
exception of the dissorophoid Micropholis,
see Schoch and Rubidge 2005) was Stereospondyli
Zittel, 1888 (Schoch and Milner 2000) (Fig. 1). This
clade radiated during the Triassic, giving rise to Rhinesuchidae Watson,
1919, Lydekkerinidae Watson, 1919, Brachyopoidea Lydekker, 1885,
Plagiosauridae Abel, 1919, Trematosauroidea Säve-Söderbergh, 1935 (which
includes the Metoposauridae Watson, 1919), and Capitosauria Yates &
Warren, 2000 (Schoch and Milner 2000; Schoch
2013). Of these, only Brachyopoidea survived beyond the Triassic
into the Early Cretaceous (Warren et al. 1997).
Rhinesuchidae and Lydekkerinidae disappeared in the Early Triassic,
while Plagiosauridae, Trematosauridae, Metoposauridae, and
Capitosauria, were thought to have gradually disappeared before the
Rhaetian, during the Middle and Late Triassic, rather than going extinct
during the end-Triassic mass extinction (Milner 1993;
Lucas and Tanner 2004, 2015).
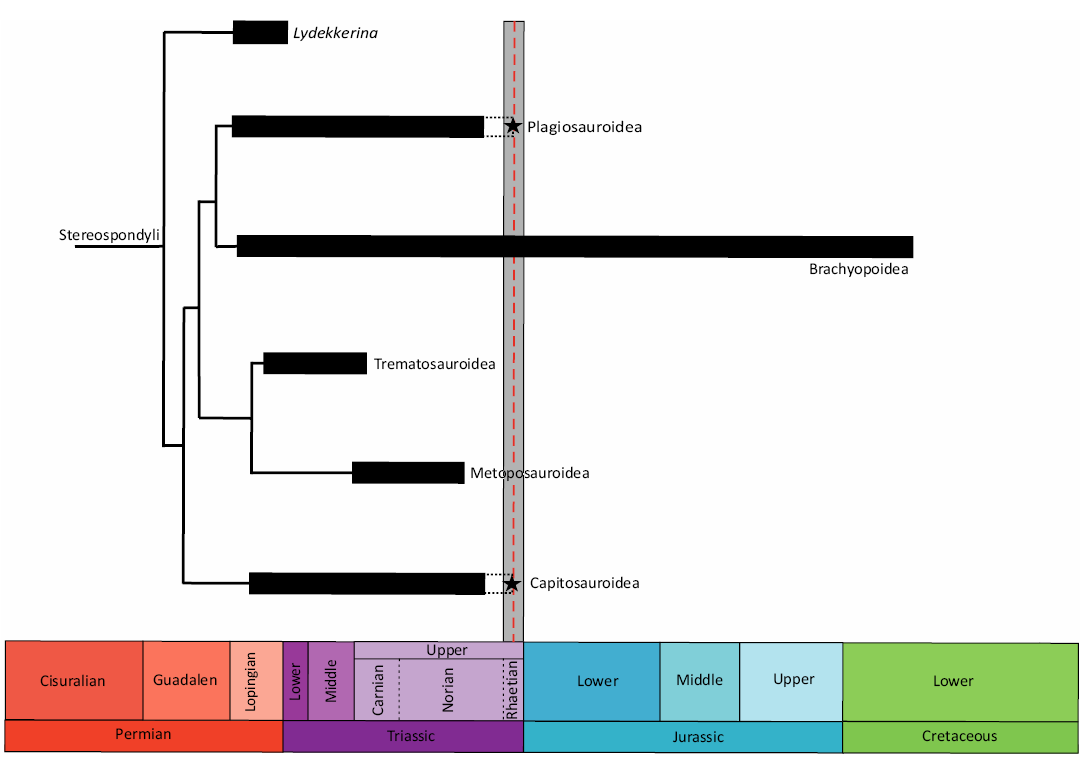
Fig. 1. Schematic diagram representing the time-calibrated phylogenetic tree of Stereospondyli. The black bars represent the known ranges of the stereospondyl clades based on the fossil record. The black dashed lines represent the uncertain presence of the taxa at that time. The grey vertical bar represents the duration of the Rhaetian, while the red dashed line represents the time to which Bonenburg was dated. The black stars represent the findings described in this paper. Note that there is no other certain record of non-brachyopoid Stereospondyli from the Rhaetian except for the fossils of Bonenburg.
The lack of well-dated potentially stereospondyl-bearing fossil localities from the Rhaetian, in contrast to their abundance during the rest of the Triassic, has always been an obstacle to testing the hypothesis of a gradual extinction of this clade (Milner 1993; Lucas and Tanner 2004, 2015).
Institutional abbreviations.—SMNS, Staatliches Museum für Naturkunde, Stuttgart, Germany; WMNM, LWL-Museum für Naturkunde, Münster, Germany.
Other abbreviations.—a, annulus; CEB, Central European Basin; ETEE, End Triassic Extinction Event; LAG, line of arrested growth; RLb, Rhaetipollis–Limbosporites palynozone b (see Fig. 3); vc, vascular canal; z, zone.
Distribution and last records of Temnospondyli in the Late Triassic
Records of all the major clades of the Stereospondyli have been found in fossil localities of Late Triassic age on almost all continents (Schoch 2013, 2014). Many such records predated the recognition of the exceptional duration of the Late Triassic Epoch of 35 million years (e.g., Ogg and Chen 2020), leading to a loss of temporal resolution. The Upper Triassic deposit with the greatest abundance of stereospondyl fossils in Europe is Krasiejów in Poland, rather imprecisely dated to the Carnian–Norian (Dzik and Sulej, 2007; Kowalski et al. 2019; Jewuła et al. 2019 and references therein). Here, a large number of specimens belonging to the metoposaurid Metoposaurus krasiejowensis have been discovered (Dzik et al. 2000; Dzik and Sulej 2007; Konietzko-Meier and Klein 2013). In addition to these finds, several remains of the capitosauroid Cyclotosaurus intermedius have been excavated (Sulej and Majer 2005). Another record of stereospondyl from the Upper Triassic of Europe is the skull of Cyclotosaurus buechneri from the Carnian of Bielefeld (Witzmann et al. 2016). The previously geologically most recent European stereospondyl record is Gerrothorax pulcherrimus, the plagiosaurid described from the Hogänäs Formation at the Bjuv coal mine in southern Sweden (Nilsson 1934, 1946). Palynological investigations have dated this formation to range from the middle to the late Rhaetian (Vajda and Wigforss-Lange 2009). Another Upper Triassic locality with stereospondyl fossils in close connection with Europe is the Fleming Fjord Formation in East Greenland (Jenkins et al. 2008; Marzola et al. 2017, 2018). The lacustrine deposits of the Fleming Fjord Formation were initially dated as early Rhaetian (Grasmück and Trümpy 1969), but recent work places them in the middle to late Norian (Clemmensen et al. 1998; Andrews et al. 2014). Also, from East Greenland, more precisely from the Kap Stewart Formation, are the putative stereospondyl remains mentioned in Milan et al. (2012), found in Rhaetian sediments. However, the fossils are not diagnostic due to their highly fragmentary preservation. Other European stereospondyl remains initially thought to be of Rhaetian age are those from a bone bed in the Algarve region of Portugal, described by Witzmann and Gassner (2008). Initially, the age assigned to the bone bed was Late Triassic to Early Jurassic, but this was revised to Carnian to Norian following the discovery of phytosaur material (Mateus et al. 2014) and of diagnostic temnospondyl material on which the new taxon Metoposaurus algarvensis was erected (Brusatte et al. 2015).
Cyclotosaurids and plagiosaurids have also been described from Lisowice in Poland (Dzik et al. 2008). The rocks exposed in the Lisowice clay pit were initially dated as Rhaetian, but the age assignment was later changed to Norian based on geological studies (Szulc et al. 2015). A similar stereospondyl fauna was also reported from Saint-Nicolas-de-Port, France (Buffetaut and Wouters 1986; Schoch and Milner 2014). Here, the age initially assigned also was Rhaetian, but the locality was subsequently re-dated as middle to late Norian (Buffetaut and Wouters 1986). One locality from Europe unequivocally dated as Rhaetian is Aust Cliff in south-west England, where bone beds containing fish and marine reptile fossils have been collected for centuries (Storrs 1994). Initially some of the fossils from these beds were attributed to Stereospondyli, but later the remains were identified as Severnichthys, a large holostean fish with teeth superficially similar to those of stereospondyls (Storrs 1994).
Also the United States has a rich record of Late Triassic stereospondyls, but again the dating of these localities is imprecise because of correlation problems of the continental red-beds of the Western Interior with marine sequences, and none of the finds can confidently be attributed to the Rhaetian (Lucas and Spielmann 2013). The metoposaurid Apachesaurus from the continental deposits of the Redonda Formation, New Mexico, has long been considered a Rhaetian stereospondyl record (Hunt 1993). The fauna of this formation has been used as the basis for the Apachean Land Vertebrate Faunachron (LVF), a biostratigraphic unit extending from the late Norian to the end of the Triassic (Lucas 2010, 2017). Biostratigraphy places the most recent occurrence of Apachesaurus in the late Rhaetian (Spielmann and Lucas 2012), but the Redonda Formation, and in general the entire Chinle Group, has uncertain magnetostratigraphic dating, making it difficult to confirm a Rhaetian age (Lucas and Spielmann 2013). Other metoposaurid taxa described from the Upper Triassic of the United States are Buettnererpeton and Anaschisma from the latest Carnian to early Norian Santa Rosa Formation, Dockum Group, Texas, and the middle Norian Popo Agie Formation, Wyoming, respectively (Gee and Kufner 2022; Kufner and Gee 2021).
Late Triassic brachyopoid fossils are known from Argentina (Marsicano 1993, 1999) and India (Sengupta 1995; Chakravorti and Sengupta 2023). Their remains are also known from post-Triassic strata in East Laurasia (Shishkin 1991; Maisch and Matzke 2005; Averianov et al. 2008) and Gondwana (Warren and Hutchinson 1983; Warren et al. 1997; Steyer and Damiani 2005). However, brachyopoids are globally absent from the Rhaetian fossil record, and totally absent from the European temnospondyl record.
There are hardly any unequivocal Rhaetian stereospondyls in the fossil record (Fig. 1). The only exception are the bones discovered at Bonenburg, which include the fossils discussed in this work as well as a humerus of Cyclotosaurus sp. described by Konietzko-Meier et al. (2019).
Geological setting
Bonenburg is a small village located in eastern North Rhine-Westphalia, Germany, close to the city of Warburg (Fig. 2A), in the Höxter District. The fossils come from the clay pit #III operated by the brick factory August Lücking GmbH & Co.KG, located 1.5 km northwest of the village. The clay pit is separated into two tectonic blocks by a fault crossing it from northwest to southeast (Fig. 2B). The northeastern part is characterized by Sinemurian sediments. In its southwestern part (51°33’45” N, 9°2’30” E), the pit exposes a continuous section from the upper Norian to the Hettangian (Figs. 2B, 3) (Sander et al. 2016; Wintrich et al. 2017; Sander and Wellnitz 2024). The upper Norian part of the section is represented by the greenish-grey marls of the Middle Keuper. No fossils have been reported from this unit. The Rhaetian succession is unusually thick and continuously exposed, as is the Triassic/Jurassic boundary (Fig. 3), making the Bonenburg pit one of the best outcrops in the epicontinental Rhaetian of Europe (Sander et al. 2016; Gravendyck et al. 2020; Sander and Wellnitz 2024).
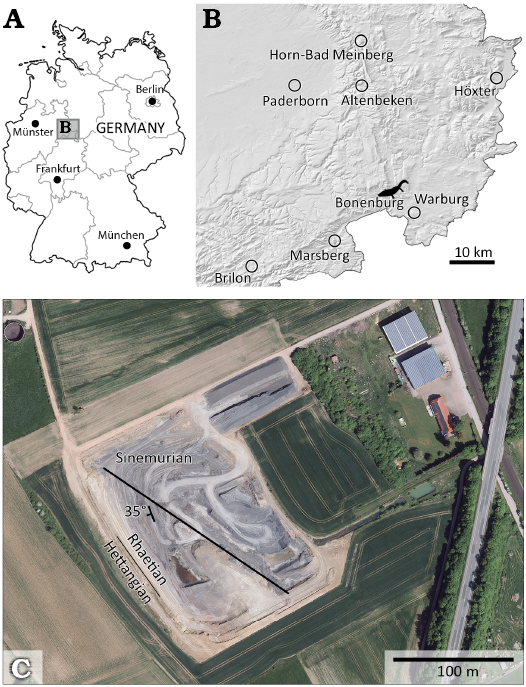
Fig. 2. Study area. A, B. Location of Bonenburg clay pit, North Rhine-Westphalia, Germany. C. Satellite image of Bonenburg clay pit. The northwest-southeast fault is indicated by the black line. The line between Rhaetian and Hettangian indicates the Triassic/Jurassic boundary. The photograph adapted from © GeoBasis-DE / BKG (2022) CC BY 4.0
The Rhaetian rocks in northern Germany are generally assigned to the Exter Formation and were laid down primarily in restricted marine conditions in the central part of the Central European Basin (CEB), known as the North German Basin (Barth et al. 2018).
The Exter Formation is subdivided into three members, beginning with the Postera Beds, followed by the Contorta Beds, and finally the Triletes Beds (Sander et al. 2016; Wintrich et al. 2017; Schobben et al. 2019; Gravendyck et al. 2020). The top of the Triletes Beds is the top of the Triassic. It is capped by the Jurassic marine transgression, represented by the fully marine Psilonotenton Formation. The mudstones with limestone interbeds of this formation contain a characteristic Hettangian ammonite fauna (Schobben et al. 2019). The Contorta Beds contain four bone beds which provided the majority of the rich and diverse vertebrate fauna from Bonenburg (Table 1 and SOM 1: table 1, Supplementary Online Material available at http://app.pan.pl/SOM/app69-Prino_etal_SOM.pdf). An exception to the bone bed origin is the articulated skeleton of the plesiosaur Rhaeticosaurus mertensi, which was found between Bone Beds 1b and 2a (Fig. 3; Wintrich et al. 2017).
Table 1. List of the fossils currently attributed to Temnospondyli from Bonenburg including the humerus WMNM P64371 that was the subject of a previous study (Konietzko-Meier et al. 2019). Specimens collected by Michael Mertens are marked by an asterisk.
|
Field N° |
Inventory N° |
Year |
Bonebed |
Taxon |
Body part |
Reference |
|
TE2 |
P97556* |
|
|
cf. Cyclotosaurus sp. |
left pterygoid fragment |
this paper |
|
P64371 |
|
2 |
cf. Cyclotosaurus sp. |
left humerus |
||
|
P97559* |
|
2b |
Capitosauroidea indet. |
right dentary |
this paper |
|
|
TE1 |
P97557* |
2018 |
2a |
Capitosauroidea indet. |
right angular in matrix |
this paper |
|
P97791 |
2018 |
2a |
Capitosauroidea indet. |
partial left clavicle |
this paper |
|
|
P98583 |
|
2a |
Capitosauroidea indet. |
partial interclavicle |
this paper |
|
|
TE3 |
P97555* |
|
3 |
Capitosauroidea indet. |
dermal bone fragment |
this paper |
|
P97790 |
|
2a |
Capitosauroidea indet. |
left femur |
this paper |
|
|
210 |
P97553 |
|
2a |
Capitosauroidea indet. |
right femur (proximal portion) |
this paper |
|
156 |
P97470 |
2017 |
2b |
Plagiosaurinae |
right parietal |
this paper |
|
333 |
P97554 |
2015 |
2a |
Plagiosaurinae |
interclavicle fragment |
this paper |
The fresh exposure in the clay pit and the organic-rich sediments allowed densely spaced palynological sampling from the Postera Beds into the Hettangian and high-resolution biostratigraphic dating of the rock units and bone beds (Gravendyck et al. 2020). The dating is based on the regional palynostratigraphic framework of the CEB (Barth et al. 2018; Gravendyck et al. 2020 and references therein) which is based primarily on drill hole sampling. Bone Bed 2 is datable with high precision as late middle Rhaetian (Gravendyck et al. 2020; Sander and Wellnitz 2024) because the bone bed is situated in the upper part of the middle subzone (RLb subzone of Gravendyck et al. 2020) of the Rhaetipollis–Limbosporites palynozone of the CEB (Fig. 3). The RLb subzone is middle Rhaetian in age.
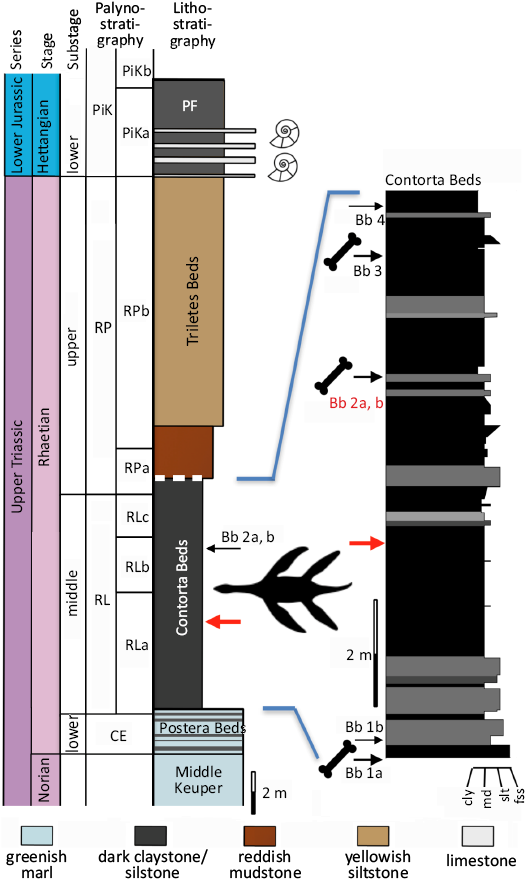
Fig. 3. Simplified stratigraphy of the Norian to Hettangian section exposed at the Bonenburg clay pit #III of August Lücking GmbH & Co. KG, North Rhine-Westphalia, Germany, with a special emphasis on the location of the bone beds and the plesiosaur skeleton in the section. The thick white dashed line on top of the Contorta Beds indicates truncation by a low-angle fault. Note that the reddish mudstones also belong to the Triletes Beds. Colors of the rock types in the main stratigraphic column approximate colors in fresh outcrop. Abbreviations: Bb, bone bed; cly, claystone; fss, fine-grained sandstone; md, mudstone; PF, Psilonotenton Formation; slt, siltstone. Palynozones: CE, Classopollis–Enzonalasporites palynozone; RL, Rhaetipollis–Limbosporites palynozone; RP, Riccisporites–Polypodiisporites palynozone; PiK, Pinuspollenites–Kraeuselisporites palynozone. Subzones are labelled alphabetically. Modified from Wintrich et al. (2017), palynostratigraphy from Gravendyck et al. (2020).
Interpolation from the geological time scale (Ogg and Chen 2020: duration of the Rhaetian from 205.7–201.36 Ma) suggests that the bone bed predates the end of the Triassic by 1.5–2 Ma. The Bonenburg section currently is probably the best-dated outcrop in the epicontinental European Rhaetian and “provides the most detailed record to date of vegetational changes leading up to the biotic crisis” (Gravendyck et al. 2020: 24). Beside stereospondyls, the fossil vertebrate fauna of Bonenburg consists of chondrichthyans, osteichthyans, ichthyosaurs, plesiosaurs, sphenodonts, putative choristoderes, phytosaurs, and synapsids (Sander et al. 2016; SOM 1: table 1).
Material and methods
The subject of the current study are 11 fossils of stereospondyls (Table 1). Some of them were recovered during excavations by the University of Bonn and the LWL-Museum für Naturkunde, others were discovered by the local collector Michael Mertens (Altenbeken, Germany). Due to their great scientific importance, M. Mertens’ finds were registered as “treasure trove finds”. This term refers to §18 of the Denkmalschutzgesetz NRW (Monument Protection Act of North Rhine-Westphalia), which states that fossils of special significance become the property of the state upon discovery. The discoverer, in this case M. Mertens, received a reward. All specimens are housed in the WMNM.
All specimens were examined and described from a morphological point of view, and some were also sectioned to observe their bone histology to aid identification.
Morphological methods.—The specimens were measured using a JOBI® vernier calliper 150 mm and a JOBI® calliper gauge 150 mm. Details of the measurements are given in the SOM. The nomenclature used in the morphological descriptions follows different publications, depending on the specimen studied. Taxonomic identifications have been made based on the literature or, in the case of fossils held in the collections of the SMNS, based on first-hand comparison by the first author.
Histological methods.—Some of the specimens were sectioned in order to study their bone histology. Long bones were sectioned at midshaft, or as close as possible to it; the dermal bones were sectioned as marked in the respective figures. The thin-sectioning procedure followed the one described by Lamm (2013) with one exception: instead of silicon carbide paper to grind the thin section, a slush of silicon carbide grinding powder was used with both grit sizes of 400 and 600. Furthermore, the sections were polished with a grit size of 800 and then covered with a coverslip. The microscope used for the observations of the sections was a LEICA DM LP polarizing light microscope (© 2022 Leica Microsystems). For overview images of the microanatomy, the sections were scanned with an EPSON PERFECTION V750 PRO flatbed scanner using a resolution of 3200 dpi. The nomenclature used in the descriptions of the histology follows de Buffrénil et al. (2021).
Systematic palaeontology
Temnospondyli Zittel, 1888
Stereospondyli Zittel, 1888
Capitosauria Yates & Warren, 2000
Capitosauroidea Säve-Söderbergh, 1935
Cyclotosauridae Shishkin, 1964
Cyclotosaurus Fraas, 1889
cf. Cyclotosaurus sp.
Figs. 4, 5.
Material.—WMNM P97556, a portion of the palatine ramus of the left pterygoid; WMNM P64371, a left humerus; all from clay pit #III of the August Lücking GmbH & Co. KG, northwest of the village of Bonenburg, North Rhine-Westphalia, Germany; Contorta Beds of the Exter Formation, Rhaetian, Triassic.
Description.—WMNM P97556 (Fig. 4; SOM 1: fig. 1): the pterygoid fragment is dorsoventrally flattened and weakly concave on the ventral side, while it is convex on the dorsal side. The medial edge, which forms the posterolateral border of the interpterygoid vacuity, is straight, with only the most proximal portion weakly medially recurved. On the other hand, the lateral edge, which contributes to the border of the subtemporal window, is weakly convex. This convexity, resulting from a lateral enlargement called “transverse flange” (Damiani 2001), probably switches to a concavity in the most proximal portion of the palatine ramus. The ventral surface of the bone is concave, but the concavity is accentuated in the medial portion, at around 1 cm from the edge, forming a sort of longitudinal groove (Fig. 4A1). The lateral-most portion of the ventral surface is rather flat but weakly ornamented (Fig. 4A1). The dorsal surface of the bone is smooth and convex along the longitudinal axis (Fig. 4A2). The ornamentation consists of a few longitudinal ridges, most evident in the medial-most portion of the ventral surface.
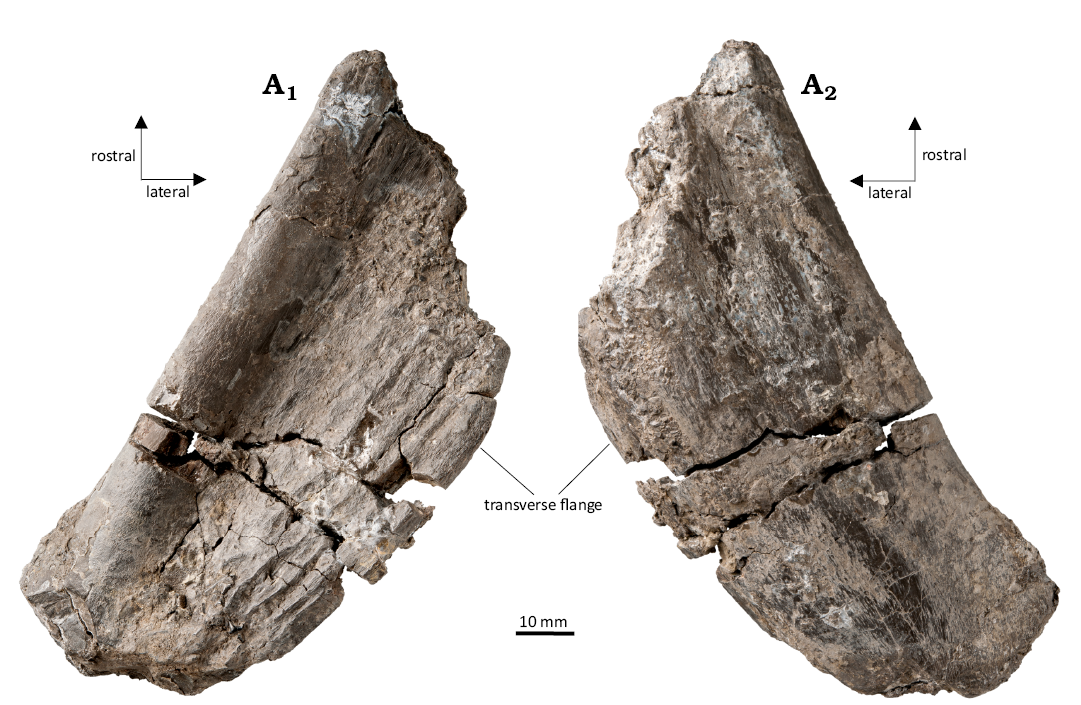
Fig. 4. Stereospondyl amphibian cf. Cyclotosaurus sp. from the Rhaetian Contorta Beds, Exter Formation, Bonenburg, North Rhine-Westphalia, Germany. WMNM P97556, portion of the palatine ramus of a left pterygoid in ventral (A1) and dorsal (A2) views.
WMNM P64371 (Fig. 5; SOM 1: figs. 2, 3): A detailed morphological and a preliminary histological description of this humerus was published by Konietzko-Meier et al. (2019). The humerus shows the typical cyclotosaur morphology, with a dorsoventrally flattened and short shaft and weakly developed supinator process (SOM 1: figs. 2, 3).
A preliminary histological description of WMNM P64371 was published by Konietzko-Meier et al. (2019). Here we summarize the previous study and update the histology with the new details about the growth pattern. The shape of the cross section is triangular, with the apex of the triangle representing the radial crest (Fig. 5A1). The entire bone section is highly porous, mostly as a result of advanced remodelling (Fig. 5A1). The inner part of the bone is crushed, and thus it is difficult to determine whether an empty medullary cavity was present. Only a very small area of the trabeculae in the center of section seems to be exclusively of secondary origin (Fig. 5A2). Most trabeculae in the deep parts of the section still consist of some primary bone in the interstitial areas between the secondary bone, and the amount of primary bone increases towards the periphery. There are distinctive growth marks in the cortex, consisting of zones, annuli, and lines of arrested growth (LAGs). Zones are made up by a poorly organized parallel-fibered matrix complemented by a large amount of longitudinal vascular canals. The organization of the matrix increases toward the bone surface. In ontogenetically older zones, vascular canals are usually enlarged by resorption, transitioning to erosion cavities. Next to the outer bone surface, the vascular canals are represented by simple canals or primary osteons. In the annuli, the tissue is always highly organized parallel-fibred matrix and less vascularized compared to the zones. The first annulus followed by the first LAG is in the inner part of the section, marking the first period of slow growth. After the first LAG, there are nine regular cycles visible, each consisting of a highly vascularized, thick zone followed by a thin, avascular annulus terminating in a LAG (Fig. 5A2). The regular pattern breaks down after the tenth LAG, where a set of closely spaced growth marks, consisting of dark lines, is visible (Fig. 5A3). The exact number is difficult to determine, but at least nine lines are visible, with the last line being more pronounced (LAG XI, Fig. 5A2, A3). Beyond this set, there are three highly vascularized zones. However, these are thinner than those in the deeper cortex (Fig. 5A3). This pattern indicates further bone growth. The thickness of zones decreases in each cycle. Each zone is followed by an annulus terminating in a double LAG (LAGs d. XII to d. XIV, Fig. 5A4). Next to the bone surface, there is one last LAG (Fig. 5A3, LAG XV).
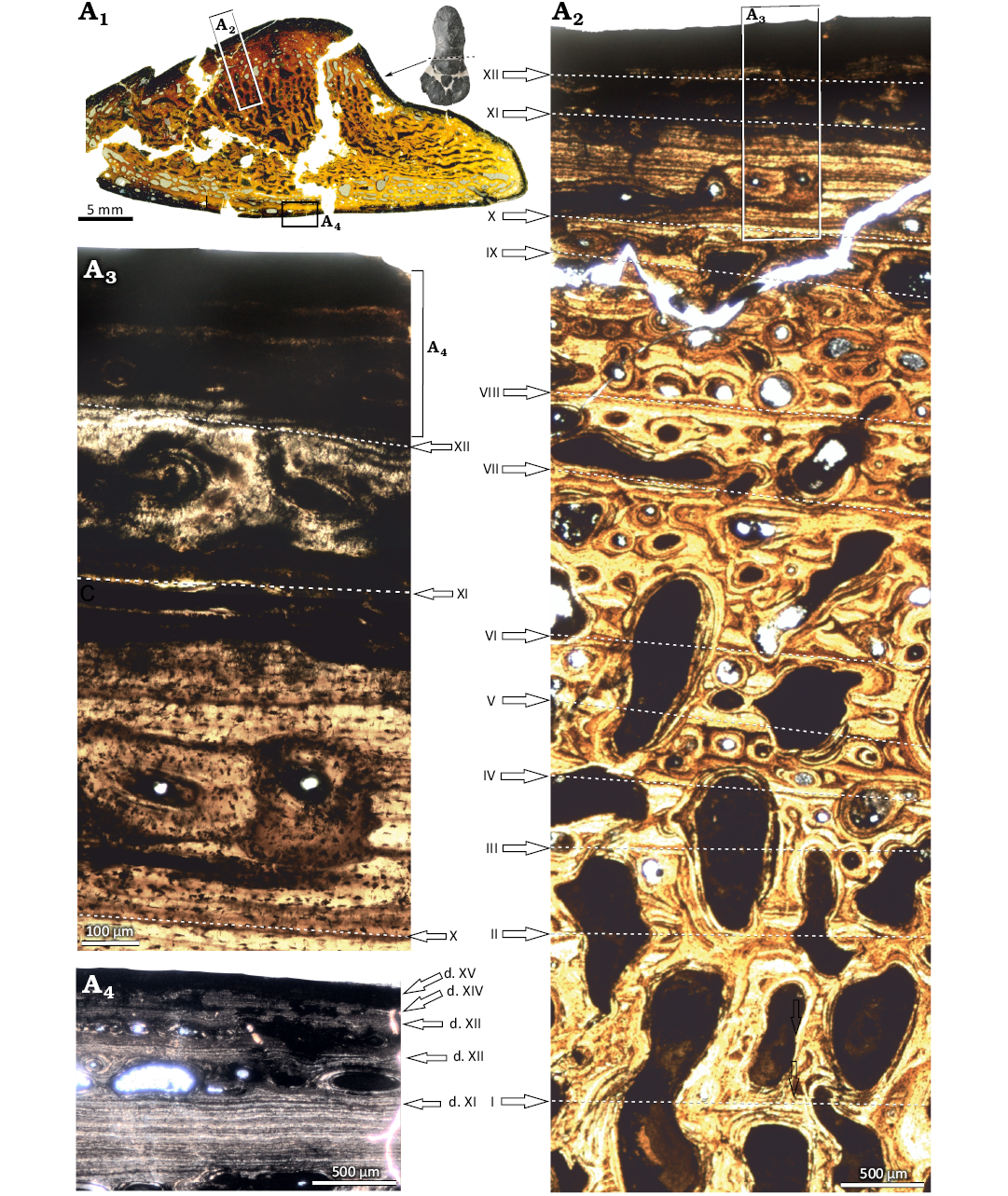
Fig. 5. Microanatomy and histology of stereospondyl amphibian cf. Cyclotosaurus sp. from the upper middle Rhaetian Bone Bed 2, Contorta Beds, Exter Formation, Bonenburg, North Rhine-Westphalia, Germany. WMNM P64371, the midshaft of humerus. Image in correct anatomical position with dorsal to the top, and thumbnail reduced image of the humerus, with a dashed line indicating the sectioning plane (A1). Close-up of cortex on ventral side; image in normal light (A2), close-up of external cortex in normal light (A3), close-up of the external-most part of cortex in polarized light (A4). Roman numbers indicate the sequence of lines of arrested growth (LAGs). Roman numbers with a “d” indicate a double LAG.
Remarks.—The morphology of the palatine ramus of the pterygoid indicates that it belongs to Capitosauria (Schoch 2008; Arbez et al. 2017; Marzola et al. 2017). However, the presence of ornamentation on the ventral side allows to exclude several capitosaurids listed in Schoch (2008: character #49). The subsequently described Stanocephalosaurus amenasensis Dahoumane et al., 2016 (Nedjari et al. 2010) shows an ornamented pterygoid but can be excluded from consideration since its palatine ramus is not concave, the ornamentation is “honeycomb-like’’ instead of straight ridges, and its geological age is too late (early Middle Triassic). Cyclotosaurus naraserluki (Marzola et al., 2017) shows ventral ornamentation on the palatine ramus of pterygoid, as in the Bonenburg fossil, and is almost coeval (see above). Thus, the shape and ornamentation of WMNM P97556 allow its identification as the palatine ramus of the left pterygoid and assignment to Cyclotosaurus sp. We assume this to be the same biological species as that represented by humerus WMNM P64371 (see below).
Based on the morphological comparison with other temnospondyl humeri, Konietzko-Meier et al. (2019) assigned humerus WMNM P64371 to Cyclotosaurus sp. As noted, the humerus represents the typical Cyclotosaurus morphology with a dorsoventrally flattened, short shaft and weakly developed supinator process (SOM 1: fig. 1). Furthermore, the bone histology of WMNM P64371 excludes amniotes from consideration (for discussion and details see Konietzko-Meier et al. 2019).
Capitosauroidea indet.
Figs. 6–16.
Material.—WMNM P97559, a partial right dentary with remains of the precoronoid and the presplenial (Bone Bed 2b); WMNM P97557, a right angular in matrix (Bone Bed 2a); WMNM P97791, a partial left clavicle (Bone Bed 2a); WMNM P98583, a portion of the left lateral wing (sensu Schoch 1999) of an interclavicle (Bone Bed 2a); WMNM P97555, a dermal bone indet. (Bone Bed 3); WMNM P97553, the proximal portion of a right femur (Bone Bed 2a); WMNM P97790, an almost complete left femur (Bone Bed 2a); all from clay pit #III of the August Lücking GmbH & Co. KG, northwest of the village of Bonenburg, North Rhine-Westphalia, Germany; Contorta Beds of the Exter Formation, Rhaetian, Triassic.
Description.—WMNM P97559 (Fig. 6; SOM 1: fig. 4): The dentary is dorsoventrally higher in the posterior portion, and it becomes lower in the rostralmost portion, just before the symphysial region. The specimen is weakly mesially recurved and is not complete, since it lacks the symphysial region and the posteriormost part of the dentary, and only the precoronoid and presplenial remain of all the bones normally attached on the lingual side. The lingual side holds a deep groove in the posterior half of the bone (Fig. 6A1). This groove is the Meckelian canal, which in a complete jaw would have been covered by the coronoids and splenials lacking here, but the sutural surface of attachment of these bones is visible in the area dorsal to the groove and just below the dental margin. The precoronoid and the presplenial are fragmentary and lack both the anterior and posterior to portions. The anteriormost part of the dentary, just before the symphysial region, is much narrower than the posterior one, and so is the groove. The labial side presents a deep groove, termed “horizontal groove” (Welles 1947) that crosses the surface diagonally, from the lowermost distal extremity up to the uppermost mesial one, close to the marginal tooth row (Fig. 6A2). Dorsally to this groove, the surface of the dentary is rather smooth, with the exception of damage produced by diagenetic alteration and weathering. Ventral to the groove, the surface is weakly ornamented. The ornamentation consists of a few ridges parallel to the long axis of the bone (Fig. 6A2). The dorsal region of the dentary holds 25 tooth sockets, all of which are elliptical with the main axis labiolingually oriented and all of the same size (Fig. 6A3).

Fig. 6. Capitosauroidea indet. from the upper middle Rhaetian Bone Bed 2b, Contorta Beds, Exter Formation, Bonenburg, North Rhine-Westphalia, Germany. WMNM P97559, dentary in lingual (A1), labial (A2), and occlusal (A3) views.
WMNM P97557 (Fig. 7; SOM 1: fig. 5): This sub-triangular bone is an angular which has the anterior part, the one contacting the dentary, highly elongated. The dorsoventrally tallest part of the bone is close to the posterior end, in correspondence with the suture between the dentary and the surangular (Fig. 7; SOM 1: fig. 5). This specimen has been prepared only labially, with the lingual face still being covered by matrix. The labial side is entirely ornamented, and the ornamentation appears as an arrangement of pits and ridges, with the ridges forming a reticular anastomosing pattern surrounding polygonal pits. The ridges are arranged radially and converge towards the region positioned ventrally to the tallest part of the bone. Proceeding distally from this region, the ridges progressively stop anastomozing and start being parallel to each other, especially in the anteriormost area.
WMNM P97791 (Figs. 8, 9; SOM 1: fig. 6): The ventral surface of the clavicular blade shows an ornamentation with an anastomozing pattern of ridges enclosing pits (Fig. 8A1). The dorsal surface of the blade is smooth in temnospondyls, but in WMNM P97791 this surface remains covered by sediment (Fig. 8A2). In lateral view, the anteriormost portion of the base of the ascending process (Rinehart and Lucas 2016) is visible (Fig. 8A2). The medial edge of the specimen shows a distinctive stepped morphology (Fig. 8A3). However, thin sectioning of the specimen revealed diagenetic fracturing as the explanation of this morphology.

Fig. 7. Capitosauroidea indet. from the upper middle Rhaetian Bone Bed 2, Contorta Beds, Exter Formation, Bonenburg, North Rhine-Westphalia, Germany. WMNM P97557, angular in matrix exposed in labial view.

Fig. 8. Capitosauroidea indet. from the upper middle Rhaetian Bone Bed 2a, Contorta Beds, Exter Formation, Bonenburg, North Rhine-Westphalia, Germany. WMNM P97791, clavicle in ventral (A1), dorsal (A2), and medial (A3) views. The dashed line indicates the histological sectioning plane.
The clavicle shows a diploë structure, typical for the dermal bone of temnospondyls with the ornamented external surface (Fig. 9A1–A3). In the external cortex, the primary tissue comprises highly organized parallel-fibered bone. The region of the cortex representing the structures of the ornamentation is almost avascular or with only irregularly arranged small vascular canals (Fig. 9A2–A5). Most of the canals show lamellar infilling, primary or secondary, while only very few canals are still simple. In the ornamented portion, the apices of the ridges all shifted medially with the growth of the bone, but still maintain the same distance between each other (Fig. 9A2, A3). The external cortex gradually transits to the medial region of the diploë. The size of the internal spaces increases with the decreasing amount of primary tissue present between cavities (Fig. 9A6, A7). However, the remains of a primary tissue are visible deep in the bone, and the trabeculae show the regular arrangement in rows. Only the most central part is composed of irregularly arragend secondary trabeculae. The medial region is bounded by the inner cortex, but the border here is also not sharp. The inner cortex is highly vascularized, and annuli and zones are easily distinguishable (Fig. 9A8, A9). The tissue in the annuli is avascular with a highly organized parallel-fibered matrix, whereas the matrix in the zones is less organized, and the longitudinal vascular canals are well arranged in rows (Fig. 9A8, A9). In the central-most region of the inner cortex, large cavities are present as a result of resorption and remodeling.
WMNM P98583 (Fig. 10; SOM 1: fig. 7): The ventral side of the interclavicle fragment is almost entirely ornamented with the exception of a portion of the lateral rim, where the clavicle once overlapped the interclavicle. The ornamentation is formed by thick, branched ridges radiating from the center of the interclavicle. In the center itself, the ridges form polygonal pits (Fig. 10). The dorsal surface is mostly smooth with the exception of weak radial striations.
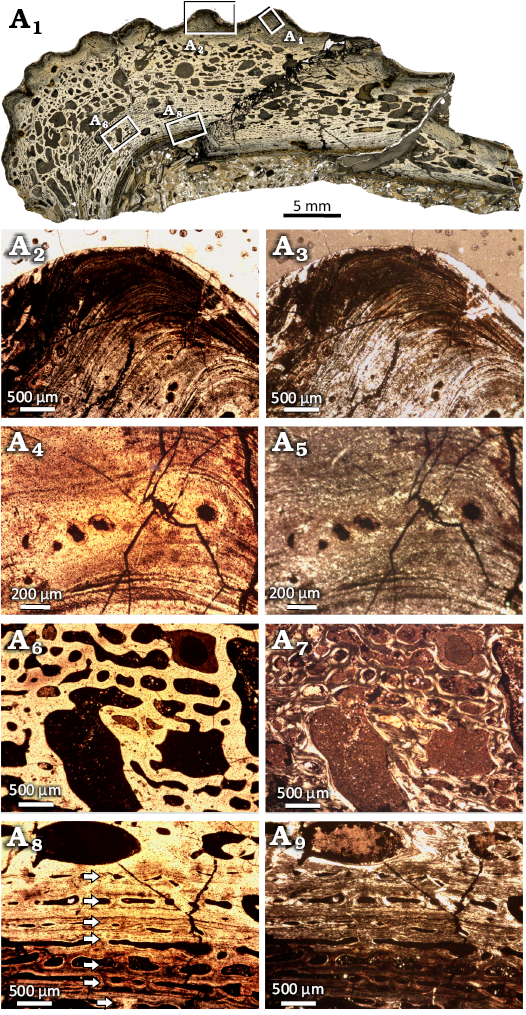
Fig. 9. Microanatomy and histology of Capitosauroidea indet. from the upper middle Rhaetian of the Contorta Beds of the Exter Formation of Bonenburg, North Rhine-Westphalia, Germany. WMNM P97791, clavicle overview image showing microanatomy (A1); close-up of external cortex, focused on ridge of ornamentation showing parallel-fibered bone with sparse, irregularly arranged vascular canals (arrows)(A2, A3); close-up of vascular canals in the external cortex (A4, A5); close-up image of the center region, with large cavities and mostly secondary tissue (A6, A7); close-up image of the visceral cortex, with annuli and zones indicated by arrows (A8, A9). Images in normal (A1, A2, A4, A6, A8) and polarized (A3, A5, A7, A9) light.

Fig. 10. Capitosauroidea indet. from the upper middle Rhaetian Bone Bed 2a, Contorta Beds, Exter Formation, Bonenburg, North Rhine-Westphalia, Germany. WMNM P98583, interclavicle fragment in ventral view.
WMNM P97555 (Figs. 11, 12): The bone is strongly weathered and is still partially embedded in sediment (Fig. 11A1, A2). Since the specimen shows ornamentation with thick ridges, it is identifiable as a dermal bone. However, due to poor preservation, it is impossible to determine what dermal bone it is.
WMNM P97555 was sectioned to observe its bone histology (Fig. 12). The internal structure of the specimen is highly crushed but the histological characters are partially preserved. The external cortex is very thin and preserved only in the region of ornamentation (Fig. 12A2, A3). The primary matrix is parallel-fibered with numerous interwoven structural fibers. Numerous longitudinal vascular canals, consisting of primary and secondary osteons, are arranged in rows (Fig. 12A2, A3). In the external-most portion, there are no vascular canals, but growth marks are visible, showing the shift of the ridges during growth. Detailed analysis of the growth marks is not possible due to the poor state of preservation. The medial region is built by secondary trabeculae separated by large cavities. In cores of some of these trabeculae, the remains of primary tissue with vascular canals can still be observed (Fig. 12A4, A5). It is important to note that the border between the external and medial regions is relatively sharp. The medial cortex is very thin and highly fragmentarily preserved (Fig. 12A6, A7). Similar to the external cortex, there is also a sharp border with the medial region. The matrix of the internal cortex is more highly organized than that of the external cortex, and the tissue is almost avascular, showing only a few very small vascular canals.

Fig. 11. Capitosauroidea indet. from the upper middle Rhaetian Bone Bed 3, Contorta Beds, Exter Formation, Bonenburg, North Rhine-Westphalia, Germany. WMNM P97555, dermal bone fragment in external (A1) and internal (A2) views. The dashed line indicates the sectioning plane.
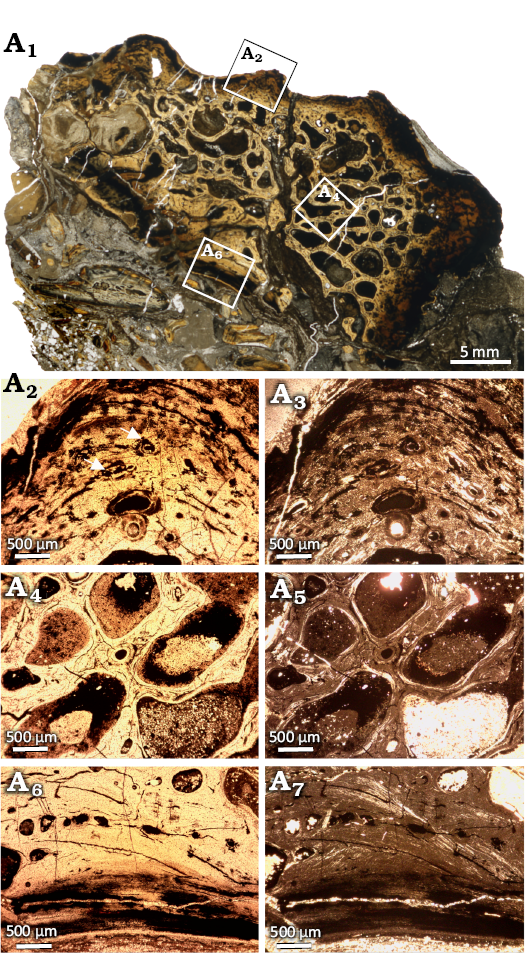
Fig. 12. Microanatomy and histology of Capitosauroidea indet. from the upper middle Rhaetian Bone Bed 3, Contorta Beds, Exter Formation, Bonenburg, North Rhine-Westphalia, Germany. WMNM P97555, dermal bone fragment. Scan of the entire section illustrating its microanatomy (A1); close-up of the external cortex, in particular of a ridge of the ornamentation where the vascular canals are visible (A2, A3); close-up of the center region, with large cavities separated by secondary trabeculae (A4, A5); close-up image of the scarcely vascularized visceral cortex (A6, A7). Images in normal (A1, A2, A4, A6) and polarized (A3, A5, A7) light.
WMNM P97553 (Figs. 13, 14; SOM 1: fig. 8): Femur heavily corroded by exposure on the sea floor and diagenetic processes, possibly compromising the morphology. The anteroventral surface of the shaft of the bone is weakly concave and smooth, except for the presence of the musculus pubisischiofemoralis internus insertion (Fig. 13A2) (Eltkin and Langer 2014). In the proximal-most portion of this surface, there is a ridge which is the result of cracking and does not correspond to any anatomical structure. The posterodorsal region shows the highly weathered remains of the internal trochanter, located on the ventral-most area of this surface (Fig. 13A1). In the proximal portion of the posterodorsal region, there is the intertrochanteric fossa (Fig. 13A1). The distal half of the bone is broken off just proximal to the proximal end of the adductor crest.
WMNM P97553 was sectioned close to the natural fracture. The histological cross section would anatomically have an oval shape, but because of crushing, the long sides of the oval are concave, resulting in a figure-eight shape (Fig. 14A1). The inner region of the bone is crushed, so it is hard to estimate the original shape of the medullary region (Fig. 14A2, A3). The transition between medullary region and cortex seems to be gradual. The size of erosion cavities decreases gradually from very large in the central-most part of the section to smaller in the primary cortex. In the middle portion of the cortex, cavities become larger again, then return to smaller sizes in the external-most cortex (Fig. 14A4, A5).
The cortex is characterized by lamellar-zonal bone tissue, with eleven annuli and zones easily distinguishable (Fig. 14A1, A4, A5). In the avascular annuli, the primary matrix is made up by highly organized parallel-fibered bone, with thick collagen fibers. The annuli are separated by zones hosting only one row of vascular canals. In the deeper part of the cortex, the vascular canals are usually altered by remodeling, represented by the secondary osteons or erosion cavities. Towards the bone surface, the vascular canals become smaller and less remodelled and mostly are primary osteons or simple vascular canals. The matrix in the zones is a poorly organized parallel-fibered bone matrix. In the cortex the distinct decrease of the growth rate expressed by thinning of the growth rings is not observed, as well as the relatively low remodeling when compared to the second femur indicate that the bone belonged to a still actively growing individual.

Fig. 13. Capitosauroidea indet. from the late middle Rhaetian Bone Bed 2a, Contorta Beds, Exter Formation, Bonenburg, North Rhine-Westphalia, Germany. WMNM P97553, proximal portion of right femur in posterodorsal (A1), anteroventral (A2), dorsal (A3), ventral (A4), and proximal (A5) views. Proximal is on the top. The dashed line indicates the histological sectioning plane.
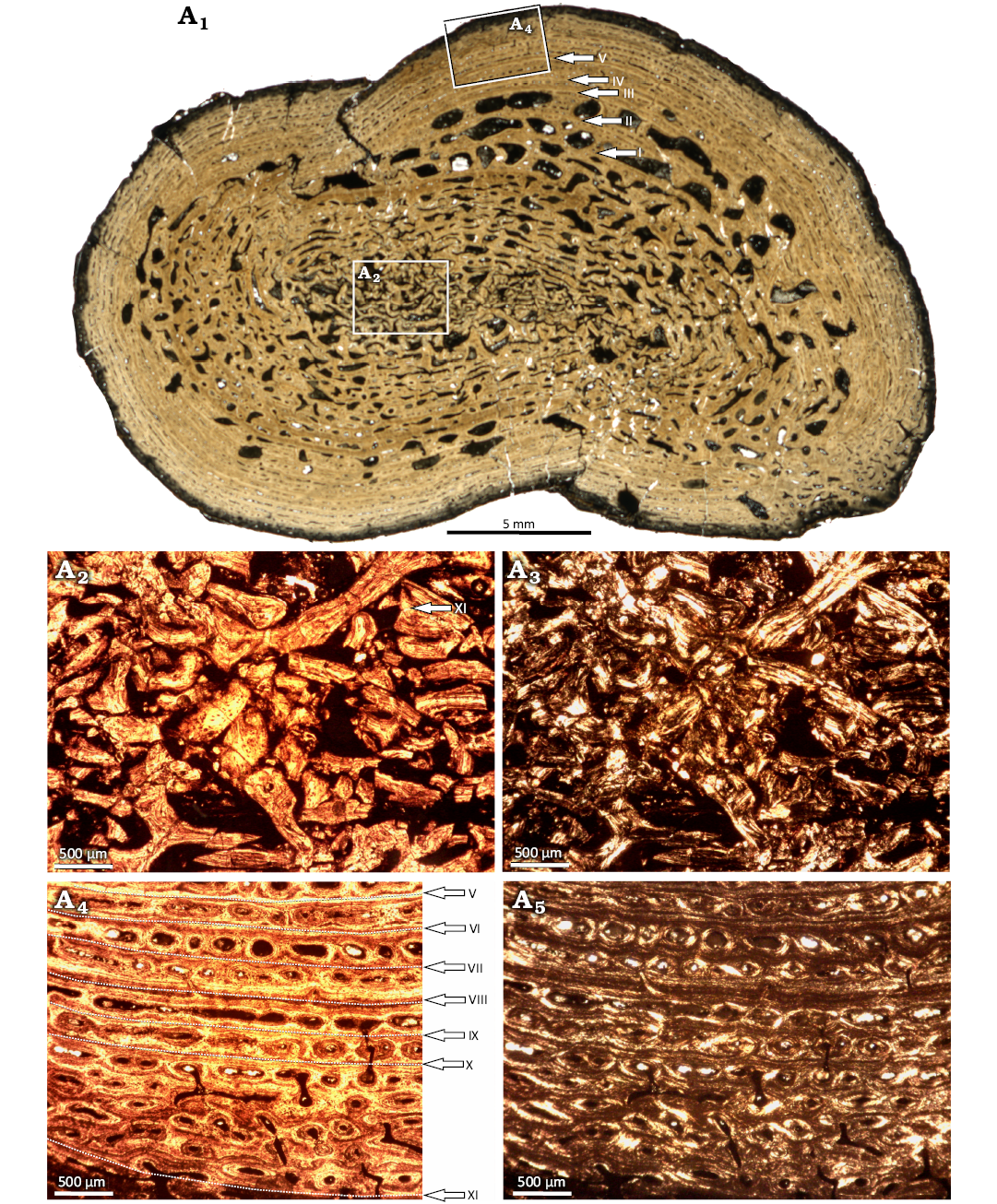
Fig. 14. Microanatomy and histology of Capitosauroidea indet. from the late middle Rhaetian Bone Bed 2a, Contorta Beds, Exter Formation, Bonenburg, North Rhine-Westphalia, Germany. WMNM P97553, proximal right femur. Scan of entire section showing the general microanatomy (A1); close-up image of the medullary region with crushed trabeculae (A2, A3); close-up image of the cortex, with annuli and zones clearly visible and indicated by arrows (A2, A3). LAGs are labelled with Roman numerals. Images in normal (A1, A2, A4) and polarized (A3, A5) light.
WMNM P97790 (Fig. 15, 16; SOM 1: fig. 9): A nearly complete femur, the proximal and distal articulation facets are damaged by corrosion, however. The incomplete preservation of the proximal and distal ends necessitates caution in estimating the femur’s size in the living animal. The shaft is long and almost straight in ventral and dorsal view. The torsion angle between the proximal and distal end is approximately 10°. The bone is dorsoventrally flattened (with the exception of the midshaft), which results in a ratio between the long and short midshaft diameter of approximately 1:1.27 (Fig. 15). At midshaft, the bone is almost round in cross-section and shows a pronounced curvature so that the dorsal edge is convex and the ventral edge is concave (Fig. 15). On the anteroventral side, the shallow indentation of the fossa poplitea is visible (Bystrow and Efremov 1940) (Fig. 15A1). In posterodorsal view, the distal end of the femur possesses a triangular indentation, the fossa tendinalis (Bystrow and Efremov 1940), which has its maximum width at its most distal point while narrowing more proximally (Fig. 15A2). These indentations separate the posterior articulation area for the fibula and the anterior articulation area for the tibia. Ventrally, the adductor crest emanates from the anterior articulation area for the tibia (Fig. 15A5). It extends proximally into the internal trochanter, aligning with the anterior margin of the shaft.
The femur (WMNM P97790) was sectioned at mid-shaft (Fig. 15A1). The shape of the section is circular, with an elevation representing the adductor crest (Fig. 16A1). Apart from a thin outermost layer, the entire section is highly remodeled and spongious (Fig. 16A1). The remains of the primary cortex consist of lamellar-zonal bone with ten visible growth cycles (Fig. 16A2). The annuli are made of well organized parallel-fibered bone, whereas the zones show a less organized matrix (Fig. 16A2). The zones mostly consist of longitudinal primary osteons, but in the deeper cortex, secondary osteons and erosion cavities are more common (Fig. 16A2).
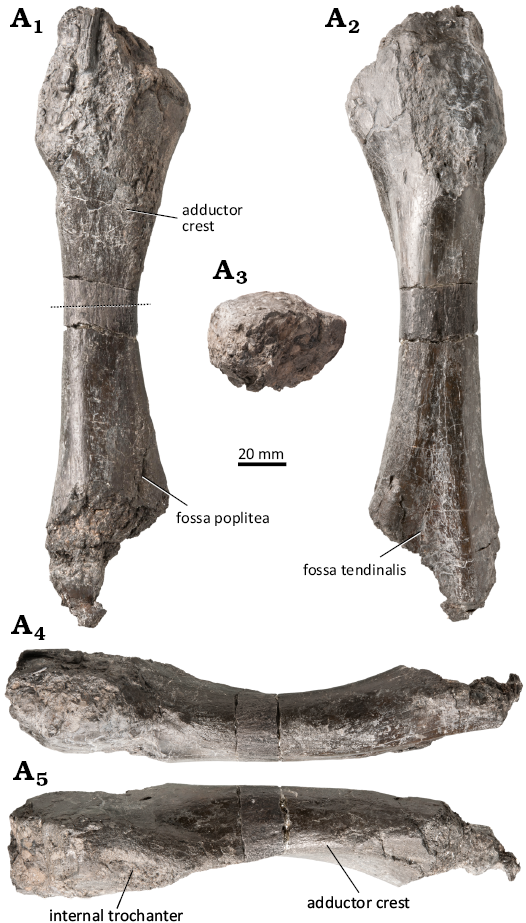
Fig. 15. Capitosauroidea indet. from the late middle Rhaetian Bone Bed 2a, Contorta Beds, Exter Formation, Bonenburg, North Rhine-Westphalia, Germany. WMNM P97790, left femur in anteroventral (A1), posterodorsal (A2), proximal (A3), dorsal (A4), and ventral (A5) views. Proximal is to the top (A1, A2) and to the left (A4, A5). The dashed line indicates the histological sectioning plane.
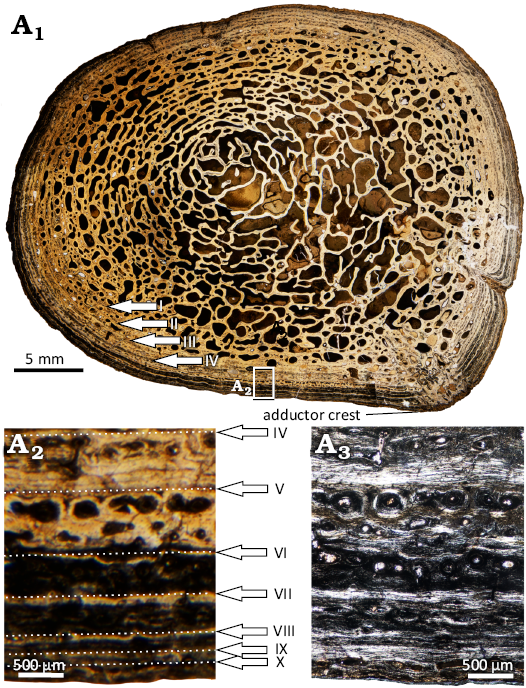
Fig. 16. Microanatomy and histology of Capitosauroidea indet. from the late middle Rhaetian Bone Bed 2a, Contorta Beds, Exter Formation, Bonenburg, North Rhine-Westphalia, Germany. WMNM P97790, left femur. Scan of the entire section showing the general microanatomy (A1), close-up image of the cortex showing ten annuli in normal light (A2) and in polarized light (A3). The Roman numbers indicate the sequence of LAGs.
Remarks.—Based on the labiolingually elliptical shape of the tooth sockets, it is possible to narrow down the identification of WMNM P97559 to either Capitosauroidea or Chigutisauridae (Warren and Davey 1992). However, the morphology points more towards Capitosauroidea, since the posterior region is enlarged compared to the anterior one, whereas in Chigutisauridae, the area just posterior to the symphysial region is enlarged (Schoch 1999; Warren 1981). Among Capitosauroidea, it is difficult to assign this specimen to a genus. This is because the most similar taxon is the Middle Triassic Mastodonsaurus giganteus Jaeger, 1828, but the precoronoid and the presplenial seem to terminate more posteriorly in this taxon, leaving a larger portion of the dentary exposed on the lingual side (Schoch 1999) than in WMNM P97559.
Since Cyclotosaurus was certainly present in Bonenburg, this would be the most logical attribution for WMNM P97559, but there are some anatomical inconsistencies. First, the groove on the labial side is parallel to the marginal tooth row in Cyclotosaurus (Sulej and Majer, 2005) but diagonal in WMNM P97559. Second, the lingually exposed portion of the dentary is smaller in Cyclotosaurus (Sulej and Majer 2005) than in the Bonenburg specimen. However, only one other dentary of Cyclotosaurus has been described (Sulej and Majer 2005), therefore intraspecific variability cannot be excluded. It is not possible to confirm or exclude the Cyclotosaurus attribution of the specimen, nor to link it to any of the other fossils described herein. For these reasons, the specimen is assigned in open nomenclature to Capitosauroidea indet.
The ornamentation of the angular WMNM P97557, with its reticular pattern of ridges and polygonal pits, is a character only shared by Capitosauroidea and Metoposauroidea (Shishkin 1987; Sulej and Majer 2005; Sulej 2007; de Buffrénil et al. 2016). Since the ornamentation is more robust than in Metoposauroidea, the specimen is assigned to Capitosauroidea. The type of ornamentation resembles that of clavicle WMNM P97791, while it shows differences to the fragment of interclavicle WMNM P98583 and the dermal bone indet. WMNM P97555, that show deeper and larger valleys between sharper ridges (see Discussion). Thus, the angular may belong to a different species than WMNM P98583 and WMNM P97555, but at the same time it is impossible to confirm or exclude its attribution to Cyclotosaurus or any other of capitosauroid. For these reasons, the specimen is assigned in open nomenclature to Capitosauroidea indet.
With its anastomozing pattern, the ornamentation of the clavicle fragment WMNM P97791 is typical of capitosauroids and metoposaurids. Nonetheless, the ridges of this specimen are relatively robust, more similar to those of capitosauroids (Schoch 1999; Sulej and Majer 2005). Furthermore, the microstructure with its high porosity seems to be more typical of capitosauroids when compared to Metoposaurus with its relatively compact bones (Kalita et al. 2022). Based on morphological and histological data, we assign clavicle WMNM P97791 to Capitosauroidea indet., with a high probability of it belonging to the same taxon as the angular described above, given the similarity of their ornamentation. On the other hand, the differences in the dermal sculpturing from the fragment of interclavicle WMNM P98583 and the dermal bone indet. WMNM P97555 are evident and could reflect a taxonomic distinction (see Discussion). We cannot exclude the possibility that the clavicle WMNM P97791 and the angular WMNM P97557 pertain to the same Cyclotosaurus species represented by the fragmentary pterygoid WMNM P97556 and the humerus WMNM P64371.
The ornamentation of the interclavicle fragment WMNM P98583 formed by ridges, grooves, and polygonal pits is typical of Capitosauroidea (Shishkin 1987; Sulej and Majer 2005; de Buffrénil et al. 2016). Nevertheless, the thickness of the ridges, the depth of the grooves, and the wide distance between ridges is quite significant, especially in comparison with other specimens described herein. The exception is the Capitosauroidea indet. dermal bone WMNM P97555, which shows a robust ornamentation as well (see Discussion). The interclavicle fragment WMNM P98583 is thus assigned to Capitosauroidea indet., and it might pertain to a different taxon than the clavicle WMNM P97791 and the angular WMNM P97557.
The ornamentation type of WMNM P97555 is characteristic of both Capitosauroidea and Metoposauroidea, but the histology excludes metoposauroid affinities since their bone is more compact (Kalita et al. 2022). The coarse nature of the bone ornamentation of WMNM P97555 suggest that it represent the same species of indeterminate capitosauroid as the fragmentary interclavicle WMNM P98583 (see above and Discussion).
The femoral fragment (WMNM P97553) does not show the clear torsion between the proximal and distal end that is typical of Trematosauria, but it shows the less pronounced torsion characteristic of Capitosauroidea (Schoch 1999; Sulej and Majer 2005). Also, Plagiosauridae can be excluded since they show an adductor crest arising from the posterior articulation area of the fibula, while in this specimen the adductor crest arises from the internal trochanter and terminates in the distal posterior articulation area for the tibia. This character and the less dorsoventrally flattened ends indicate a similarity of the specimen to Paracyclotosaurus davidii Watson, 1958, rather than Mastodonsaurus giganteus (Jaeger, 1828) (Watson 1958; Schoch 1999). The specimen herein described can thus be assigned to Capitosauroidea, with more similarities to Paracyclotosaurus than to Mastodonsaurus. Histological data suggest similarity with humerus WMNM P64371 and femur WMNM P97790 (see below), however again due to our scanty knowledge of the variation of histological characters among capitosaurs, it is difficult to state if these bones represent the same capitosauroid taxon at different ontogenetic stages or two separate taxa (see Discussion for more details).
Identification of WMNM P97790 as a capitosauroid femur emerges as the sole viable option from comparative analysis, given that the humerus in Stereospondyli features a short, flattened shaft with widened ends distally and posteriorly. The specimen shows resemblance with femur WMNM P97553 (see above). Regardless of its exact affinity, this specimen represents the largest Stereospondyli femur preserved in the fossil record. The limited availability of stereospondyl limbs, exemplified by the scarcity of described Brachyopoidea femora (only three worldwide), poses challenges to comparing specimen WMNM P97790 to other Stereospondyli. Nevertheless, the simplified limb bone morphology (compared to terrestrial archosaurs and synapsids) observed in Stereospondyli results from their adaptation to an aquatic lifestyle and aligns well with the characteristics exhibited by WMNM P97790. The femur of Mastodonsaurus giganteus reveals an elongated straight shaft with broadened proximal and distal ends, the latter being more expanded. In the dorsal view, the femur exhibits axial symmetry, convexity, and relative smoothness, except for the distal triangular fossa tendinalis. Conversely, the ventral aspect is concave, hosting a fossa poplitea and an asymmetrical adductor crest. These features demarcate articulation regions for the fibula and tibia in Mastodonsaurus, akin to those observed in WMNM P97790. Despite similarities, distinctions emerge in the dorsoventrally flattened articulation areas of M. giganteus compared to WMNM P97790. Notably, the adductor crest in M. giganteus originates from the distal posterior fibular articulation extending to the proximal central internal trochanter, while in WMNM P97790, it arises from the anterior tibial articulation extending to the anterior internal trochanter. The overall morphological resemblance between M. giganteus and WMNM P97790 suggests a probable affinity with Capitosauroidea.
Additionally, the preliminary study of the histology reveals similarities with the small femur WMNM P97553 and the cf. Cyclotosaurus sp. humerus WMNM P64371 (see above and Discussion). However, due to the rudimentary knowledge about histological variability among capitosaur (Konietzko-Meier et al. 2019; Teschner et al. 2023) and very scanty morphological data for cyclotosaur postcrania (Sulej and Majer 2005), it is not possible to confirm the Cyclotosaurus affinity of the femur, and it is thus assigned to Capitosauroidea indet. A dedicated study of the morphology and histology of WMNM P97790 is the subject of a forthcoming paper by one of the authors (LG).
Plagiosauridae Abel, 1919
Plagiosaurinae Shishkin, 1986
Plagiosaurinae indet.
Figs. 17, 18.
Material.—WMNM P97470, an almost complete right parietal; WMNM P97554, a partial right lateral process of an interclavicles; both from clay pit #III of the August Lücking GmbH & Co. KG, northwest of the village of Bonenburg, North Rhine-Westphalia, Germany; Bone Bed 2a, Contorta Beds of the Exter Formation, Rhaetian, Triassic.
Description.—WMNM P97470 (Fig. 17; SOM 1: fig. 10): an almost complete right parietal. The specimen is dorsoventrally flattened and is relatively narrow in the anterior region, while it is mediolaterally enlarged in the central and posterior regions. The bone is slightly thicker on the medial edge, corresponding to the margin of the pineal foramen (Fig. 17A1). The anterolateral margin of the parietal is concave and does not show any signs of damage, indicating that it is a natural anatomical edge, probably the border of the orbit. The other edges of the bone are all fracture surfaces. While the visceral surface of the bone is smooth, the external one is ornamented. The ornamentation is characterized by pustules arranged in radial lines from the center of the bone. In some areas, pustules are fused together, forming thin radial ridges.
WMNM P97554 (Fig. 18; SOM 1: fig. 11): a portion of the right lateral process of the interclavicle, as it is partially ornamented and partially covered with faint striations on the ventral side (Fig. 18A1; SOM 1: fig. 11). On the dorsal side it also shows the thickened ridge (Gee and Sidor 2022) typical of the lateral processes (Fig. 18A2).
Remarks.—The type of ornamentation seen in WMNM P97470, with pustules sometimes arranged in thin radial ridges, is characteristic of most plagiosaurids. More specifically, the ornamentation of WMNM P97470 is very similar to that of Plagiosaurinae, since it does not show polygonal pits and anastomosing ridges, which is the case for Plagiosterninae (Jaekel 1914; Hellrung 2003; Jenkins et al. 2008). Given this similarity of the ornamentation with Plagiosaurinae (Shishkin 1986), the specimen is assigned to this group. Since the specimen shows a natural edge in the anterolateral region (Fig. 17), the parietal contributed to the border of the orbit. In their redescription of Plagiosaurus depressus Jaekel, 1914, Witzmann and Schoch (2024) recognized that the postfrontal was independently lost in this plagiosaurine, enabling the parietal to contribute to the orbital margin. On the contrary, in the other known plagiosaurine, Gerrothorax pulcherrimus (Fraas, 1913), the parietal is excluded from the orbital margin due to the presence of the postfrontal (Hellrung 2003; Schoch and Witzmann 2012). Therefore, WMNM P97470 might be a parietal of Plagiosaurus depressus or a form more closely related to it than to Gerrothorax, but we prefer to be cautious and assign it only to Plagiosaurinae indet. due to the absence of other diagnostic fossils.
The ornamentation seen in WMNM P97554, with pustules arranged in radial ridges or scattered randomly, corresponds to Plagiosaurinae and differs from the one of Plagiosterninae, in which pustules are only present at the junctions of ridges bordering polygonal pits (Shishkin 1973, 1987; see also Witzmann et al. 2010; Schoch et al. 2014). Given the fact that the interclavicle fragment lacks further diagnostic characters, it is assigned to Plagiosaurinae indet.

Fig. 17. Plagiosaurinae indet. from the late middle Rhaetian Bone Bed 2a, Contorta Beds, Exter Formation, Bonenburg, North Rhine-Westphalia, Germany. WMNM P97470, right parietal in dorsal (A1) and ventral (A2) views.

Fig. 18. Plagiosaurinae indet. from the late middle Rhaetian Bone Bed 2a, Contorta Beds, Exter Formation, Bonenburg, North Rhine-Westphalia, Germany. WMNM P97554, interclavicle in ventral (A1) and dorsal (A2) views.
Discussion
The variability of capitosauroid dermal ornamentation in Bonenburg.—From a morphological perspective, bone ornamentation represented the most important character for identifying the dermal bones herein described. First of all, it allows easy distinction of the fossils of Stereospondyli Zittel, 1888, from those of all the other tetrapod taxa that can be found in Bonenburg, like ichthyosaurs, plesiosaurs, and Pachystropheus, as they do not show any dermal ornamentation (Sander et al. 2016; Wintrich et al. 2017). Secondly, bone ornamentation permits recognition of some of the fossils as Plagiosaurinae Shishkin, 1986 (WMNM P97470 and WMNM P97554, a parietal and a fragment of an interclavicle, respectively) among the other remains of Stereospondyli, thanks to their unique “pustular” ornamentation composed of isolated bony pustules which may be arranged in radiating lines. All the other ornamented bones were attributed to Capitosauroidea Säve-Söderbergh, 1935. Two of these fossils, the pterygoid (WMNM P97556) and the dentary (WMNM P97559), present a weakly developed ornamentation, hence they were assigned to Capitosauroidea based on other morphological characters (for the pterygoid, the simple presence of the ornamentation, rather than its type, was considered diagnostic; Schoch 2008). The remaining dermal bones of Capitosauroidea all showed the so-called “honeycomb” ornamentation, characterized by polygonal cells enclosed by prominent ridges that form a reticular pattern (Rinehart and Lucas 2013). Even though the ornamentation of all these specimens can be defined as “honeycomb-type”, two distinct morphological subgroups can be identified: one with smoother and more closely spaced ridges and shallower pits, and the other with more widely spaced, thicker and sharper ridges, with deeper pits and valleys in between. The angular (WMNM P97557, Fig. 7) and the clavicle (WMNM P97791, Fig. 8) belong to the first morphotype, while the fragment of interclavicle (WMNM P98583, Fig. 10) and the unidentified dermal bone (WMNM P97555, Fig. 11) belong to the second morphotype. These two morphotypes show evident differences, which suggest the presence of two separate taxa of Capitosauroidea in Bonenburg, each of them with its specific type of “honeycomb” ornamentation. However, despite always being mentioned as an important trait of dermal bones, the ornamentation is generally not considered a diagnostic character within Capitosauroidea (Schoch 2008). Witzmann et al. (2010) tested the phylogenetic potential of the ornamentation or dermal sculpture, stating that it is helpful in distinguishing the main temnospondyl lineages, but it appears to be insufficient for a more precise diagnosis. Antczak and Bodzioch (2018) used a quantitative approach as well to the study of ornamentation, but only applied it to observe intraspecific variation in Metoposaurus krasiejowensis. However, Antczak and Bodzioch (2018) noticed that the Cyclotosaurus honey-comb cells have a statistically larger diameter when compared to metoposaurs. Consequently, the subdivision of the capitosauroid fossils of Bonenburg into two subgroups is highly probable, but not confirmed. Furthermore, this dichotomous separation based on ornamentation cannot be extended to the other capitosauroid fossils described herein, either because they are not ornamented (since they are not dermal bones, like the two femora WMNM P97790 and WMNM P97553 and the humerus WMNM P64371), or because their ornamentation is limited (like the pterygoid WMNM P97556 and the dentary WMNM P97559). As an attempt to provide more evidence that involved all the capitosauroid specimens, we decided to use bone histology with a diagnostic purpose.
Histology to support the morphological diagnosis.—Bone histology is a useful tool for identifying a fossil lacking ornamentation as a stereospondyl. This is especially true for the long bones, which may show morphological resemblance to some terrestrial reptiles. The histology of Stereospondyli bones, however, is clearly different from the rest of the fauna of Bonenburg (Konietzko-Meier et al. 2019), allowing assignment to this clade. The long bone microstructure of ichthyosaurs varies from very loose spongy organization with a thin cortex to a more compact spongiosa with very small vascular cavities, which are located circumferentially in the cortex and are distributed randomly in the center (Houssaye et al. 2014). Generally, ichthyosaurs lack an open medullary cavity, and the center of the bone is completely filled with cancellous bone (Lopuchowycz and Massare 2002). The long bones of temnospondyls have larger vascular cavities than ichthyosaurs, and these cavities are arranged in a layered, circumferential arrangement, unlike ichthyosaurs, in which the trabeculae are all of similar size (Konietzko-Meier et al. 2019). Another difference is that in ichthyosaurs, the primary trabeculae are in part constituted by woven-fibered bone matrix, whereas this is absent in the Bonenburg stereospondyls (Konietzko-Meier et al. 2019).
Plesiosaur histology has been extensively studied in the past ( Kiprijanoff 1881; Wiffen et al., 1995; Fostowicz-Frelik and Gaździcki 2001; Salgado et al. 2007; Liebe and Hurum 2012; Wintrich et al. 2017; see Sander and Wintrich 2021 for a review). The histology of plesiosaur primary bone tissue cannot be mixed up with any taxon of temnospondyls because of the dominance of radial fibrolamellar bone (Sander and Wintrich 2021). Moreover, in plesiosaurs, growth marks manifest a sudden directional change of the radial canals (Sander and Wintrich 2021), while in the temnospondyls of Bonenburg there are annuli and lines of arrested growth (LAGs) (Konietzko-Meier et al. 2019).
Concerning the differences within Temospondyli, the microanatomy of the clavicle fragment WMNM P97791 is inconsistent with that of metoposaurids (Fig. 9) because of the greater compactness of their dermal bone, which is always above 50% (Kalita et al. 2022). The bone histology of clavicle WMNM P97791 shows similarities with that of Cyclotosaurus intermedius (Kalita et al. 2022). The inner cortex is very similar in both samples, being highly vascularized and showing distinctive zones and annuli (Fig. 9A8; Kalita et al. 2022: fig. 4b). However, the transition between the inner cortex and the center region is gradual in specimen WMNM P97791 and abrupt in C. intermedius. In the Bonenburg specimen, the center region still preserves some patches of primary bone among the secondary bone, and the resorption cavities, even though they are larger than in the inner cortex, are still relatively small (Fig. 9A6, A7). On the other hand, the center region of C. intermedius only shows secondary bone, and the resorption cavities are distinctly larger. Finally, whereas the external cortex of WMNM P97791 is almost avascular, the one of C. intermedius is highly vascularized (Fig. 9A2, A3; Kalita et al. 2022: fig. 4b). However, the potential of histological features of dermal bones for taxonomic studies is poorly explored. Only Witzmann (2009) raised this question and noted variations in the compactness of dermal bones in different non-amniote taxa. Gruntmejer et al. (2016) and Kalita et al. (2022) suggested that temnospondyl dermal bone structure may be sensitive to mechanical stress and ontogenetic stage.
The microanatomy of dermal bone indet. WMNM P97555 (Fig. 12) differs from clavicle WMNM P97791. In WMNM P97555, the ridges forming the ornamentation are more vascularized, and avascular tissue only forms the external-most portion of each ridge (Fig. 12A2, A3). Also, the primary tissue of the ridges shows interwoven structural fibers. Furthermore, the erosion cavities in the center region are not arranged in any order, and their size is highly variable (Fig. 12A4, A5). These observations suggest that dermal bone indet. WMNM P97555 belongs to a different taxon than the clavicle WMNM P97791 and the angular WMNM P97557.
The bone histology of WMNM P97553, the proximal portion of the right femur, was compared with the humerus WMNM P64371 (Konietzko-Meier et al. 2019) and the larger femur WMNM P97790. These bones all share some common histological features, such as a highly vascularized cortex with parallel-fibred matrix (Figs. 5, 14, and 16). The cortex in all specimens is formed by lamellar-zonal bone with well expresed zones and annuli terminated by LAGs (Figs. 14A2, A3 and 16A4, A5). Also, the size of erosion cavities decreases from the medullary region towards the cortex. All these similarities suggest that all three bones may belong to the same taxon. However, the thickness and the composition of the cortex in the two femora is different: while in the femur WMNM P97790, the cortex is relatively thin and already shows signs of secondary remodeling (Fig. 16A1), the cortex in WMNM P97553 is thicker in relation to the diameter of the bone and almost exclusively composed of primary tissue (Fig. 14A4, A5). The explanation of this important difference might be that the two femora represent different histological ontogenetic stages (HOS) of the same taxon, with the smaller femur being a younger individual, still actively growing, and the larger femur being an older one, approaching maximum size. This approach is similar to the HOS concept described by Klein and Sander (2008) for sauropods as well as Sander and Wintrich (2021) for plesiosaurs. The gradual change of the histological characters with the preservation of the main histological framework also applies to Metoposaurus krasiejowensis intercentra (Konietzko-Meier et al. 2012).
It is questionable whether the humerus belongs to the same taxon as the two femora. Histologically, some of the similarities of the femora are shared with the humerus WMNM P64371 (Fig. 5) (see also Konietzko-Meier et al. 2019): large erosion cavities in the medullary region, progressively decreasing in size towards the external region and a cortex formed by lamellar-zonal bone with highly organized annuli and zones. Well expressed growth marks with thin annuli and comparable thicker zones seem to be a typical feature of Cyclotosaurus in contrast to Metoposaurus (Teschner et al. 2023), which additionally confirms the taxonomic assignment of the Bonenburg humerus. However, this humerus is only half of the length of the large femur but shows the same or an only slightly younger histological ontogenetic stage. Both bones are highly remodelled, and a distinct decrease of growth rate is present. However, among stereospondyls, the length difference between fore- and hind limbs is never as great as between the humerus and the large femur; usually humerus and femur are of similar length (Dutuit 1976). This makes it rather improbable that the humerus and the large femur belong to the same individual. On the other hand, the small femur, which seems to fit better with the size of the humerus, is distinctly ontogenetically younger.
In general, humerus and femur histology are not entirely comparable in Temnospondyli, however, because they differ in the expression of growth marks (Konietzko-Meier and Sander 2013; Konietzko-Meier and Klein 2013; Teschner et al. 2018). This is because in the region of the adductor crest, the erosion cavities and the vascular canals are differently oriented from the rest of the medullary region. Also, the cortex in the adductor crest has a different thickness from the rest of the bone (Teschner et al. 2018, 2023). Another reason why it is difficult to draw conclusions based on the histology, but also on the morphology, is that no femur of Cyclotosaurus has been described yet, neither morphologically, nor histologically. This impedes any sort of comparison with the only stereospondyl taxon that was definitely present in Bonenburg (Konietzko-Meier et al. 2019).
The presence of Plagiosaurinae in Bonenburg.—The attribution of the parietal (WMNM P97470) and the interclavicle (WMNM P97554) to Plagiosaurinae Shishkin, 1986, is strongly supported by the unique type of ornamentation they display (Shishkin 1973, 1987; Witzmann et al. 2010; Schoch et al. 2014). However, the parietal shows some peculiar morphological traits that make it an important specimen in this collection. The clade Plagiosaurinae consists of only two genera, Gerrothorax and Plagiosaurus (Witzmann and Schoch 2024). Although similar in morphology to that of Gerrothorax, the parietal WMNM P97470 has a natural anterolateral margin, representing part of the margin of the orbit (Fig. 17A2). This differentiates it from Gerrothorax, where the parietal does not contribute to the orbit due to the presence of the postfrontal (Hellrung 2003; Schoch et al. 2014). The loss of the postfrontal has been considered a diagnostic character that evolved independently in both Plagiosuchinae and Plagiosterninae, but not in Plagiosaurinae (Schoch et al. 2014). The other Late Triassic plagiosaurine taxon is Plagiosaurus depressus Jaekel, 1914, of which only the posterior part of the skull, fragments of the lower jaw and parts of the pectoral girdle, humerus and axial skeleton are known (Jaekel 1914; Nilsson 1937; Hellrung 2003; Witzmann and Schoch 2024). Jaekel (1914) and Nilsson (1937) reconstructed a large postfrontal and a small parietal. Hellrung (2003: 107) disagreed with this view and interpreted the postfrontal as a parietal and a small bone fragment anterior to it as a remnant of a very short postfrontal. Witzmann and Schoch (2024) agreed with the interpretation of Hellrung (2003) but suggested that the small fragment is actually part of the parietal. The boundary between the supposed postfrontal and the parietal appears to be a crack rather than a suture. Nilsson’s photo (Nilsson 1937: pl. 5) lacks this crack, suggesting that it occurred later. These considerations, plus the pronounced pustular and radial sculpture, suggest that the Bonenburg parietal might belong to a new taxon closely related to Plagiosaurus depressus or to P. depressus itself. However, because of the fragmentary nature of this fossil, we opt for a more cautious assignment attribution to Plagiosaurinae indet.
Extinction of non-brachyopoid stereospondyls and the end-Triassic extinction event.—The original hypothesis about the extinction of stereospondyls was that they disappeared at the very end of the Triassic, killed off by the end-Triassic mass extinction event (Colbert 1958). This hypothesis was later challenged by Milner (1993), Tanner et al. (2004), and Lucas (2017). These authors noted that stereospondyls gradually disappeared before the mass extinction, which Lucas and Tanner (2004: 31) considered “a myth” in its suddenness and severity. In his latest review, Lucas (2017) affirmed that “capitosaurid records are no younger than Revueltian, and metoposaurids and plagiosaurids have records that end in the Apachean”. If the correlation between the “Land Vertebrate Faunachrons” and the International Chronostratigraphic Chart is correct, equating the Revueltian with the Norian and the Apachean with the end of the Norian and the Rhaetian (but see Rayfield et al. 2009 for a criticism on the matter), capitosaurs thus went extinct before the Rhaetian and the only Rhaetian non-brachyopoid stereospondyls were metoposaurs and plagiosaurs (Lucas 2017). However, the description of the cf. Cyclotosaurus humerus WMNM P64371, Konietzko-Meier et al. (2019) demonstrated that at least capitosaurs survived to the Rhaetian. The presence of capitosaurs in Bonenburg is further confirmed by the fossils described in this work, which also makes it appear possible that more than one capitosaur taxon is preserved in this locality. Furthermore, the occurrence of plagiosaurids, only hinted at in Konietzko-Meier et al. (2019), is confirmed by the discovery of parietal WMNM P97470 and interclavicle WMNM P97554. This plagiosaurid material belongs to the subfamily Plagiosaurinae, as do Gerrothorax and Plagiosaurus, the geologically youngest known plagiosaurids (Jaekel 1914; Nilsson 1946; Buffetaut and Wouters 1986). However, none of these occurrences can be dated as precisely as Bonenburg, and two of the three occurrences (those of Plagiosaurus) cannot even be shown to be Rhaetian with certainty (see also Witzmann and Schoch 2024, Sander and Wellnitz 2024). On a last note, if the parietal WMNM P97470 actually pertains to Plagiosaurus, it would represent the first find of this genus outside of Halbertstadt.
The environment and the provenance of the Stereospondyli of Bonenburg.—While the presence of Stereospondyli in the Rhaetian of Europe is now confirmed, the environments in which they lived remains an open question. The bone beds of Bonenburg were deposited in an open marine environment in the CEB (Sander et al. 2016; Schobben et al. 2019; Gravendyck et al. 2020), which is recorded by abundant Chondrichthyes, Osteichthyes, plesiosaurs, and ichthyosaurs. Terrestrial or freshwater taxa, including stereospondyls, are allochthonous. Salinity in the CEB has been demonstrated to be lower than the one of the open sea, with a shift towards extensively brackish conditions in the eastern area of the basin due to the influx of fluvial water from the Fennoscandian and Bohemian highs (Fischer et al. 2012). However, although some tolerance to salinity has been hypothesised for stereospondyls (Marzola et al. 2017 for capitosauroids; Witzmann and Soler-Gijon 2010; Hellrung 2003; Sanchez and Schoch 2013; Schoch et al. 2022 for plagiosaurids; Pawlak et al. 2022; Czepiński et al. 2023 for both, whereas some trematosaurs where clearly marine, e.g., Lindemann 1991), it is unlikely that they were permanently inhabiting saltwater and brackish environments. Their remains must have been transported from a freshwater habitat located on the land masses surrounding the CEB, as is the case for several other faunal elements from the Contorta Beds of Bonenburg (Sander et al. 2016; Sander and Wellnitz 2024). These landmasses probably are the Fennoscandian and Bohemian highs, which provided the freshwater and the debris material that formed the vast deltaic plains in the CEB (Bachmann et al. 2008; Fischer et al. 2012; Gravendyck et al. 2020). In particular, a deltaic system was present in the area around Bebenhausen, near Tübingen (Aepler 1974). These deltas were the result of a stillstand, a break in the rise of the sea level (transgression) that characterized the Late Triassic in the CEB. From these deposits has been recovered a dermal bone fragment of a capitosauroid housed at the Staatliches Museum für Naturkunde, Stuttgart, labelled as “Mastodonsaurus sp.” (SMNS 91479, AP personal observation; SOM 1: fig. 12). While the attribution to Mastodonsaurus sp. is far from being confirmed, the fossil shows the same type of ornamentation as the interclavicular fragment (WMNM P98583) and the unidentified dermal bone (WMNM P97555) found in Bonenburg, suggesting that the same taxon was present also here. Beside this, fossils of cynodonts (Clemens and Martin 2014) have also been found in these deposits, which also represent part of the continental faunal input at Bonenburg (Schwermann 2016). However, the dating of these bone beds is imprecise due to time averaging of the deposition, but nevertheless the majority of vertebrates found are of Rhaetian age (Aepler 1974; Clemens and Martin 2014). The same type of fauna has been found in other localities from the former CEB or from Greenland that have a similar age as Bonenburg (the Höganäs Formation, Saint-Nicolas-de-Port and the Fleming Fjord Formation, see Introduction). The issue with these findings is their problematic dating, contrarily to Bonenburg (Schobben et al. 2019; Gravendyck et al. 2020), or their fragmentary status, but their existence supports the concept that the extinction of non-brachyopoid stereospondyls, at least in Europe, did not occur until the end of the Triassic. In other localities, especially in North America, it seems like the stereospondyl fauna disappeared before the Rhaetian (Lucas 2017). This might be due to a regional biological and environmental cause or due to a dating bias in the fossil record. Moreno et al. (2024) studied the changes in biogeographic distribution of different stereospondyl clades from the Anisian to the Carnian, showing how habitat suitability varied during this time across Pangea. Extending this type of study to the Rhaetian might shed light on the habitat suitability of North America and the CEB at the time, possibly explaining the differences between the two faunas. This, and further research about the CEB sites mentioned above, will enlighten the true story behind the extinction of the non-brachyopoid Stereospondyli.
Another important point regarding temnospondyl extinction during the ETEE is the absence of brachyopoids from the entire European fossil record. Brachyopoids are the only group of stereospondyls to survive the ETEE and lived on until the Late Jurassic in Asia (Shishkin 1991; Averianov et al. 2008), Africa (Steyer and Damiani 2005) and in Australia until the Early Cretaceous (Warren et al. 1997; Vickers-Rich 1996; Steyer and Damiani 2005). Late Triassic records of the group exist also from Argentina and India (Marsicano 1993, 1999; Sengupta 1995), but there is no record of them in Europe. This absence is peculiar since Europe at that time was located between Eurasia and Gondwana, both representing territories populated by brachyopoids, and brachyopoids should have crossed Europe during their dispersal. One possibility is that brachyopoids did occur in the European Rhaetian but were not recognized as such. Some of the specimens herein described and assigned to Capitosauroidea sp. (i.e., the interclavicle WMNM P98583, the dermal bone indet. WMNM P97555, and the two femora (WMNM P97790 and WMNM P97553)) could actually pertain to Brachyopoidea, but their incomplete state of preservation and the scarcity of comparative material currently makes testing of this hypothesis difficult.
Conclusions
The stereospondyl remains described here are from the Rhaetian Contorta Beds of Bonenburg, North Rhine-Westphalia, Germany. The importance of these finds lies in their stratigraphic age, since no other stereospondyls have ever been found in sediments unambiguously dated as Rhaetian, in this case based on palynostratigraphy, biostratigraphy, and carbon isotopes (Sander et al. 2016; Schobben et al. 2019; Gravendyck et al. 2020). This contradicts the notion that all Stereospondyli, with the exception of Brachyopoidea, gradually became extinct during the Triassic, before the Rhaetian. The fossils pertain to two stereospondyl groups, the Capitosauroidea and the Plagiosaurinae. While one capitosauroid from Bonenburg had already been described previously as cf. Cyclotosaurus sp. (Konietzko-Meier et al. 2019), the fossils of Plagiosaurinae are the first reports from this locality. For both taxa, these fossils represent the stratigraphically youngest occurrences in the fossil record. The identification of the specimens is mainly based on morphological features, such as the type of ornamentation for dermal bones. Nevertheless, when morphology was not sufficient, specimens were sectioned to observe their histology and microanatomy to corroborate the assignment. Variation in bone surface ornamentation and microanatomy of the capitosauroid specimens from Bonenburg suggests the presence of two species of this group, one with smooth ridges and shallow pits, and the other with thick and pointy ridges and deep pits. However, dermal bone sculpture and microanatomy are not considered diagnostic, therefore the presence of two species of capitosauroid in Bonenburg is probable, but not confirmed. The new fossils help to shed light on the extinction of non-brachyopoid stereospondyls, which probably perished in the end-Triassic mass extinction event rather than gradually dying out through the Late Triassic. Other European localities, slightly older than Bonenburg, bear fossils of a fauna similar to the one described here, which means that at least in the Central European Basin, stereospondyls were fairly abundant and diverse in the Late Triassic.
Acknowledgements
We would like to thank the State of North Rhine-Westphalia for the financial support of the excavations in Bonenburg and the accessioning of the fossils as part of the paleontological heritage management. We would also like to thank the company August Lücking GmbH & Co. KG in Bonenburg, in particular its director Joachim Thater, for the excellent cooperation and technical support for the annual excavation activities. Special thanks go to the Wiemers family as landowners, who also support the excavations. Citizen scientist Michael Mertens (Altenbeken, Germany) is thanked for the many years of trustful cooperation. We are indebted to Olaf Dülfer of the Section Paleontology of the Bonn Institute of Organismic Biology, University of Bonn, as well as Michael Ludorf and Jerome Gores (both WMNM) and the numerous students and excavation volunteers for their work in the field and the preparation of the fossils. The photos of the specimens were taken by Georg Oleschinski (Section Paleontology of the Bonn Institute of Organismic Biology, University of Bonn). The petrographic thin sections were done in the paleohistological laboratory (Section Paleontology of the Bonn Institute of Organismic Biology, University of Bonn, Germany) by Olaf Dülfer and his co-workers. We thank Tanja Schaaf (Section Paleontology of the Bonn Institute of Organismic Biology, University of Bonn, Germany) for the supervision on the Master’s thesis of Laurent Garbay about the femur WMNM P97790. Two reviewers (Eudald Mujal and Sanjukta Chakravorti [both State Museum of Natural History, Stuttgart, Germany]) are appreciated for their very useful comments which greatly improved the manuscript. This is contribution number 4 of the DFG Research Unit 5581 “Evolution of life histories in early tetrapods”.
References
Abel, O. 1919. Die Stämme der Wirbeltiere. 914 pp. De Gruyter, Berlin, Crossref
Aepler, R. 1974. Der Rhätsandstein von Tübingen – ein kondensiertes Delta. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 147: 113–162.
Andrews, S.D., Kelly, S.R., Braham, W., and Kaye, M. 2014. Climatic and eustatic controls on the development of a Late Triassic source rock in the Jameson Land Basin, East Greenland. Journal of the Geological Society 171: 609–619. Crossref
Antczak, M. and Bodzioch, A. 2018. Ornamentation of dermal bones of Metoposaurus krasiejowensis and its ecological implications. PeerJ 6: e5267. Crossref
Arbez, T., Dahoumane, A., and Steyer, J.S. 2017. Exceptional endocranium and middle ear of Stanocephalosaurus (Temnospondyli: Capitosauria) from the Triassic of Algeria revealed by micro-CT scan, with new functional interpretations of the hearing system. Zoological Journal of the Linnean Society 180: 910–929. Crossref
Averianov, A.O., Martin, T., Skutschas, P.P., Rezvyi, A.S., and Bakirov, A.A. 2008. Amphibians from the Middle Jurassic Balabansai Svita in the Fergana Depression, Kyrgyzstan (Central Asia). Palaeontology 51: 471–485. Crossref
Bachmann, G.H., Voigt, T., Bayer, U., von Eynatten, H., Legler, B., and Littke, R. 2008. Depositional history and sedimentary cycles in the Central European Basin System. In: R. Littke, U. Bayer, D. Chajewski, and S. Nelskamp (eds.), Dynamics of Complex Intracontinental Basins. The Example of the Central European Basin System, 157–172. Springer, Heidelberg.
Barth, G., Franz, M., Heunisch, C., Ernst, W., Zimmermann, J., and Wolfgramm, M. 2018. Marine and terrestrial sedimentation across the T-J transition in the North German Basin. Palaeogeography Palaeoclimatology Palaeoecology 489: 74–94. Crossref
Benton, M.J. 2015. When Life Nearly Died: The Greatest Mass Extinction of All Time. Second Edition. 352 pp. Thames & Hudson, New York.
Boy, J.A., 1972. Die Branchiosaurier (Amphibia) des saarpfälzischen Rotliegenden (Perm, SW-Deutschland). Abhandlungen des hessischen Landesamtes für Bodenforschung 65: 1–137.
Brusatte, S.L., Butler, R.J., Mateus, O., and Steyer, J.S. 2015. A new species of Metoposaurus from the Late Triassic of Portugal and comments on the systematics and biogeography of metoposaurid temnospondyls. Journal of Vertebrate Paleontology 35: e912988. Crossref
Buffetaut, E. and Wouters, G. 1986. Amphibian and reptile remains from the Upper Triassic of Saint-Nicolas-de-Port (eastern France) and their biostratigraphic significance. Modern Geology 10: 133–145.
Bystrow, A.P. and Efremov, I.A. 1940. Benthosuchus sushkini Efr.—a labyrinthodont from the Eotriassic of Sharjenga river. Trudy Paleontologičeskogo Instituta Akademiâ Nauk SSSR 10: 1–152.
Chakravorti, S. and Sengupta, D.P. 2023. The first record of chigutisaurid amphibian from the Late Triassic Tiki Formation and the probable Carnian pluvial episode in central India. PeerJ 11: e14865. Crossref
Clemens, W.A. and Martin, T. 2014. Review of the non-tritylodontid synapsids from bone beds in the Rhaetic Sandstone, southern Germany. Paläontologische Zeitschrift 88: 461–479. Crossref
Clemmensen, L.B., Kent, D.V., and Jenkins, F.A. 1998. A Late Triassic lake system in East Greenland: facies, depositional cycles and palaeoclimate. Palaeogeography, Palaeoclimatology, Palaeoecology 140: 135–159. Crossref
Colbert, E.H. 1958. Tetrapod extinctions at the end of the Triassic Period. Proceedings of the National Academy of Sciences USA 44: 973–977. Crossref
Czepiński, Ł., Pawlak, W., Rytel, A., Tałanda, M., Szczygielski, T., and Sulej, T. 2023. A new Middle Triassic vertebrate assemblage from Miedary (southern Poland). Journal of Vertebrate Paleontology. 43:2, e2265445 Crossref
Damiani, R.J. 2001. A systematic revision and phylogenetic analysis of Triassic mastodonsauroids (Temnospondyli: Stereospondyli). Zoological Journal of the Linnean Society 133: 379–482. Crossref
de Buffrénil, V., Clarac, F., Canoville, A., and Laurin, M. 2016. Comparative data on the differentiation and growth of bone ornamentation in gnathostomes (Chordata: Vertebrata). Journal of Morphology 277: 634–670. Crossref
de Buffrénil, V., de Ricqlès, A.J., Zylberg, L., and Padian, K. (eds.) 2021. Vertebrate Skeletal Histology and Paleohistology. 837 pp. CRC Press, Paris. Crossref
Dutuit, J.-M. 1976. Introduction à l’étude paléontologique du Trias continental marocain. Description des premiers stégocéphales recueillis dans le couloir d’Argana (Atlas occidental). Mémoires du Muséum National d’Histoire Naturelle, Paris, Serie C 36: 1–253.
Dzik, J. and Sulej, T. 2007. A review of the early Late Triassic Krasiejów Biota from Silesia, Poland. Palaeontologia Polonica 64: 3–27.
Dzik, J., Sulej, T., Kaim, A., and Niedźwiedzki, R. 2000. Późnotriasowe cmentarzysko kręgowców lądowych w Krasiejowie na Śląsku Opolskim. Przegląd Geologiczny 48: 226–235.
Dzik, J., Sulej, T., and Niedzwiedzki, G. 2008 A dicynodont-theropod association in the latest Triassic of Poland. Acta Palaeontologica Polonica 53: 733–738. Crossref
Eltink, E. and Langer, M.C. 2014. A new specimen of the temnospondyl Australerpeton cosgriffi from the late Permian of Brazil (Rio do Rasto Formation, Paraná Basin): comparative anatomy and phylogenetic relationships. Journal of Vertebrate Paleontology 34: 524–538. Crossref
Erwin, D.H., Bowring, S.A., and Yugan, J. 2002. End-Permian mass extinctions: A review. In: C. Koeberl and K.G. MacLeod (eds.), Catastrophic Events and Mass Extinctions: Impacts and Beyond. Geological Society of America Special Paper 356: 363–383. Crossref
Fischer, J., Voigt, S., Franz, M., Schneider, J.W., Joachimski, M., Tichomirowa, M., Götze, J., and Furrer, H. 2012. Palaeoenvironments of the Late Triassic Rhaetian Sea: implications from oxygen and strontium isotopes of hybodont shark teeth. Palaeogeography, Palaeoclimatology, Palaeoecology 353: 60–72. Crossref
Fostowicz-Frelik, Ł. and Gaździcki, A. 2001. Anatomy and histology of plesiosaur bones from the Late Cretaceous of Seymour Island, Antarctic Peninsula. In: A. Gaździcki (ed.), 52 Palaeontological Results of the Polish Antarctic Expeditions. Part III. Palaeontologia Polonica 60: 7–32.
Fraas, E. 1889. Die Labyrinthodonten der schwäbischen Trias. Palaeontographica 36: 1–158.
Gee, B.M. and Kufner, A.M. 2022. Revision of the Late Triassic metoposaurid “Metoposaurus” bakeri (Amphibia: Temnospondyli) from Texas, USA and a phylogenetic analysis of the Metoposauridae. PeerJ 10: e14065. Crossref
Gee, B.M. and Sidor, C.A. 2022. Cold capitosaurs and polar plagiosaurs: new temnospondyl records from the upper Fremouw Formation (Middle Triassic) of Antarctica. Journal of Vertebrate Paleontology 41(4): e1998086-2. Crossref
Grasmück, K. and Trümpy, R. 1969. Triassic stratigraphy and general geology of the country around Fleming Fjord (East Greenland). Meddelelser om Grønland 2: 5–71.
Gravendyck, J., Schobben, M., Bachelier, J., and Kürschner, W.M. 2020. Macroecological patterns of the terrestrial vegetation history during the end-Triassic biotic crisis in the Central European Basin: A palynological study of the Bonenburg section (NW-Germany) and its supra-regional implications. Global and Planetary Change 194: 103286. Crossref
Gruntmejer, K., Konietzko-Meier, D., and Bodzioch, A. 2016. Cranial bone histology of Metoposaurus krasiejowensis (Amphibia, Temnospondyli) from the Late Triassic of Poland. PeerJ 4: e2685. Crossref
Hellrung, H. 2003. Gerrothorax pustuloglomeratus, ein Temnospondyle (Amphibia) mit knöcherner Branchialkammer aus dem Unteren Keuper von Kupferzell (Süddeutschland). Stuttgarter Beiträge zur Naturkunde, Serie B 330: 1–132.
Houssaye, A., Scheyer, T.M., Kolb, C., Fischer, V., and Sander, M.P. 2014. A new look at ichthyosaur long bone microanatomy and histology: Implications for their adaptation to an aquatic life. PLoSONE, 9(4): e95637. Crossref
Hunt, A.P. 1993. Revision of the Metoposauridae (Amphibia, Temnospondyli) and description of a new genus from Western North America. In: M. Morales (ed.), Aspects of Mesozoic Geology and Paleontology of the Colorado Plateau. Museum of Northern Arizona Bulletin 59: 67–97.
Jäger, G.F.V. 1824. De Ichthyosauri sive Proteosauri, Fossilis, Speciminibus, in Agro Bollensi in Wurtembergia Repertis. 14 pp. Cotta, Stuttgart.
Jäger, G.F.V. 1828. Über die fossilen Reptilien, welche in Württemberg aufgefunden worden sind. 72 pp. J.B. Metzler, Stuttgart. Crossref
Jaekel, O. 1914. Über die Wirbeltierfunde in der oberen Trias von Halberstadt. Paläontologische Zeitschrift 1: 155–215. Crossref
Jenkins, F.A., Shubin, N.H., Gatesy, S.M., and Warren, A. 2008. Gerrothorax pulcherrimus from the Upper Triassic Fleming Fjord Formation of East Greenland and a reassessment of head lifting in temnospondyl feeding. Journal of Vertebrate Paleontology 28: 935–950. Crossref
Jewuła, K., Matysik, M., Paszkowski, M., and Szulc, J. 2019. The late Triassic development of playa, gilgai floodplain, and fluvial environments from Upper Silesia, southern Poland. Sedimentary Geology 379: 25–45.
Kalita, S., Teschner, E.M., Sander, P.M., and Konietzko-Meier, D. 2022. To be or not to be heavier: the role of dermal bones in the buoyancy of the Late Triassic temnospondyl amphibian Metoposaurus krasiejowensis. Journal of Anatomy 241: 1459–1476. Crossref
Kear, B.P., Poropat, S.F., and Bazzi, M. 2016. Late Triassic capitosaurian remains from Svalbard and the palaeobiogeographical context of Scandinavian Arctic temnospondyls. Geological Society, London, Special Publications 434: 113–126. Crossref
Kiprijanoff, V. 1881. Studien über die fossilen Reptilien Russlands. Mémoire de Académie Impériale des Sciences de St. Petersbourg. 7: 28–31.
Klein, N. and Sander, P.M. 2008. Ontogenetic stages in the long bone histology of sauropod dinosaurs. Paleobiology 34: 247–263. Crossref
Konietzko-Meier, D. and Klein, N. 2013. Unique growth pattern of Metoposaurus diagnosticus krasiejowensis (Amphibia, Temnospondyli) from the Upper Triassic of Krasiejów, Poland. Palaeogeography, Palaeoclimatology, Palaeoecology 370: 145–157. Crossref
Konietzko-Meier, D. and Sander, P.M. 2013. Long bone histology of Metoposaurus diagnosticus (Temnospondyli) from the Late Triassic of Krasiejów (Poland) and its paleobiological implications. Journal of Vertebrate Paleontology 35: 1–16. Crossref
Konietzko-Meier, D., Bodzioch, A., and Sander, P.M. 2012. Histological characteristics of the vertebral intercentra of Metoposaurus diagnosticus (Temnospondyli) from the Upper Triassic of Krasiejów (Upper Silesia, Poland). Earth and Environmental Science Transactions of the Royal Society of Edinburgh 103: 237–250. Crossref
Konietzko-Meier, D., Shelton, C.D., and Sander, P.M. 2016. The discrepancy between morphological and microanatomical patterns of anamniotic stegocephalian postcrania from the Early Permian Briar Creek Bonebed (Texas). Comptes Rendus Palevol 15: 103–114. Crossref
Konietzko-Meier, D., Werner, J., Wintrich, T., and Sander, P.M. 2019. A large temnospondyl humerus from the Rhaetian (Late Triassic) of Bonenburg (Westphalia, Germany) and its implications for temnospondyl extinction. Journal of Iberian Geology 45: 287–300. Crossref
Kowalski, J., Bodzioch, A., Janecki, P.A., Ruciński, M.R., and Antczak, M. 2019. Preliminary report on the microvertebrate faunal remains from the Late Triassic locality at Krasiejów, SW Poland. Annales Societatis Geologorum Poloniae 89: 291–305. Crossref
Kufner, A.M. and Gee, B.M. 2021. Reevaluation of the holotypes of Koskinonodon princeps Branson and Mehl, 1929, and Borborophagus wyomingensis Branson and Mehl, 1929 (Temnospondyli, Metoposauridae). Journal of Vertebrate Paleontology 41: e1922067. Crossref
Lamm, E.T. 2013. Preparation and sectioning of specimens. In: K. Padian, and E.T. Lamm (eds.), Bone Histology of Fossil Tetrapods: Advancing Methods, Analysis, and Interpretation, 55–160. University of California Press, Los Angeles. Crossref
Liebe, L. and Hurum, J.H. 2012. Gross internal structure and microstructure of plesiosaur limb bones from the Late Jurassic, central Spitsbergen. Norwegian Journal of Geology 92: 285–309.
Lindemann, F.J. 1991. Temnospondyls and the Lower Triassic paleogeography of Spitsbergen. Contributions from the Paleontological Museum, University of Oslo 364: 39–40.
Lopuchowycz, V.B. and Massare, J.A. 2002. Bone microstructure of a Cretaceous ichthyosaur. Paludicola 3: 139–147.
Lucas, S.G. 2010. The Triassic timescale based on nonmarine tetrapod biostratigraphy and biochronology. In: S.G. Lucas (ed.), The Triassic Timescale. Geological Society London, Special Publication 334: 447–500. Crossref
Lucas, S.G. 2017. The Late Triassic timescale. In: L.H. Tanner (ed.), The Late Triassic World: Earth in a Time of Transition. Topics in Geobiology 46: 1–25. Crossref
Lucas, S.G. and Spielmann, J.A. 2013. Magnetostratigraphy of the Upper Triassic Chinle Group in New Mexico: An appraisal of 40 years of analysis. In: L.H. Tanner, J.A. Spielmann, and S.G. Lucas (eds.), The Triassic System. New Mexico Museum of Natural History and Science, Bulletin 61: 375–381.
Lucas, S.G. and Tanner, L.H. 2004. Late Triassic extinction events. Albertiana 31: 31–40.
Lucas, S.G. and Tanner, L.H. 2015. End-Triassic nonmarine biotic events. Journal of Palaeogeography 4: 331–348. Crossref
Lydekker, R. 1885. The Reptilia and Amphibia of the Maleri and Denwa Groups. Palaeontologia Indica Series IV 1 (3): 30–38.
Maisch, M.W. and Matzke, A.T. 2005. Temnospondyl amphibians from the Jurassic of the Southern Junggar Basin (NW China). Paläontologische Zeitschrift 79: 285–301. Crossref
Marsicano, C.A. 1993. Postcranial skeleton of a brachyopoid (Amphibia, Temnospondyli) from the Triassic of Mendoza (Argentina). Alcheringa 17: 185–189. Crossref
Marsicano, C.A. 1999. Chigutisaurid amphibians from the Upper Triassic of Argentina and their phylogenetic relationships. Palaeontology 42: 545–565. Crossref
Marsicano, C., Angielczyk, K.D., Cisneros, J.C., Richter, M., Kammerer, C.F., Fröbisch, J., and Smith, R.M. 2021. Brazilian Permian dvinosaurs (Amphibia, Temnospondyli): revised description and phylogeny. Journal of Vertebrate Paleontology 41: e1893181. Crossref
Marzola, M., Mateus, O., Milàn, J., and Clemmensen, L.B. 2018. A review of Palaeozoic and Mesozoic tetrapods from Greenland. Bulletin of the Geological Society of Denmark 66: 21–46. Crossref
Marzola, M., Mateus, O., Shubin, N.H., and Clemmensen, L.B. 2017. Cyclotosaurus naraserluki, sp. nov., a new Late Triassic cyclotosaurid (Amphibia, Temnospondyli) from the Fleming Fjord Formation of the Jameson Land Basin (East Greenland). Journal of Vertebrate Paleontology 37: e1303501. Crossref
Mateus, O., Butler, R.J., Brusatte, S.L., Whiteside, J.H., and Steyer, J.S. 2014. The first phytosaur (Diapsida, Archosauriformes) from the Late Triassic of the Iberian Peninsula. Journal of Vertebrate Paleontology 34: 970–975. Crossref
Milàn, J., Clemmensen, L.B., Adolfssen, J.S., Estrup, E.J., Frobøse, N., Klein, N., Mateus, O., and Wings, O. 2012. A preliminary report on coprolites from the Late Triassic part of the Kap Stewart Formation, Jameson Land, East Greenland. New Mexico Museum of Natural History and Science Bulletin 57: 203–206.
Milner, A.R. 1990. The radiations of temnospondyl amphibians. In: P.D. Taylor and G.P. Larwood (eds.), Major Evolutionary Raditaions, 321–349. Clarendon Press, Oxford. Crossref
Milner, A.R. 1993. Amphibian-grade Tetrapoda. In: M.J. Benton (ed.), The Fossil Record 2. 2: 665–679. Chapman & Hall, London.
Milner, A.R. and Sequeira, S.E.K. 1994. The temnospondyl amphibians from the Viséan of East Kirkton, West Lothian, Scotland. Transactions of the Royal Society of Edinburgh: Earth Sciences 84: 331–361. Crossref
Moreno, R., Dunne, E.M., Mujal, E., Farnsworth, A., Valdes, P.J., and Schoch, R.R. 2024. Impact of environmental barriers on temnospondyl biogeography and dispersal during the Middle–Late Triassic. Palaeontology 67: e12724. Crossref
Nedjari, A., Aït Ouali, R., Bitam, L., Steyer, J.S., Taquet, P., Vacant, R., Bouzidi, W., and Kedadra, B. 2010. Découverte d’un nouveau gisement de stégocéphales d’une conservation exceptionnelle dans le Trias d’in Amenas (basin d’Illizi, Algérie). Bulletin du Service Géologique National 21: 1–18.
Nilsson, T. 1934. Vorläufige Mitteilung über einen Stegocephalenfund aus dem Rhät Schonens. Geologiska Föreningen I Stockholm Förhandlingar 56: 428–442. Crossref
Nilsson, T. 1937. Ein Plagiosauride aus dem Rhät Schonens. Beiträge zur Kenntnis der Organisation der Stegocephalengruppe Brachyopoidei. Lunds Universitets Årsskrift N.F. 34: 1–74.
Nilsson, T. 1946. A new find of Gerrothorax rhaeticus Nilsson a plagiosaurid from the Rhaetic of Scania. Lunds Universitets Årsskrift N. F. 42: 1–42.
Ogg, J.G. and Chen, Z.-Q. 2020. The Triassic Period. In: F.M. Gradstein, J.G. Ogg, M.D. Schmitz, and G.M. Ogg (eds.), The Geologic Time Scale 2020, Vol. 2, 903–953. Elsevier, Boston. Crossref
Pawlak, W., Rozwalak, P. and Sulej, T. 2022. Triassic fish faunas from Miedary (Upper Silesia, Poland) and their implications for understanding paleosalinity. Palaeogeography, Palaeoclimatology, Palaeoecology 590: 110860. Crossref
Pawley, K. and Warren, A. 2005. A terrestrial stereospondyl from the Lower Triassic of South Africa: the postcranial skeleton of Lydekkerina huxleyi (Amphibia: Temnospondyli). Palaeontology 48: 281–298. Crossref
Pawley, K. and Warren, A. 2006. The appendicular skeleton of Eryops megacephalus Cope, 1877 (Temnospondyli: Eryopoidea) from the Lower Permian of North America. Journal of Paleontology 80: 561–580. Crossref
Rayfield, E.J., Barrett, P.M., and Milner, A.R. 2009. Utility and validity of Middle and Late Triassic “land vertebrate faunachrons”. Journal of Vertebrate Paleontology 29: 80–87. Crossref
Rinehart, L.F. and Lucas, S.G. 2013. The functional morphology of dermal bone ornamentation in temnospondyl amphibians. In: L.H. Tanner, J.A. Spielmann, and S.G. Lucas (eds.), The Triassic System. New Mexico Museum of Natural History and Science, Bulletin 61: 524–532.
Rinehart, L.F. and Lucas, S.G. 2016. Eocyclotosaurus appetolatus, a Middle Triassic amphibian: osteology, life history and paleobiology. New Mexico Museum of Natural History and Science 70: 1–104.
Salgado, L., Fernandez, M.S., and Talevi, M. 2007. Observaciones histologicas en reptiles marinos (Elasmosauridae y Mosasauridae) del Cretacico Tardio de Patagonia y Antartida. Ameghiniana 44: 513–523.
Sanchez, S. and Schoch, R.R. 2013. Bone histology reveals a high environmental and metabolic plasticity as a successful evolutionary strategy in a long-lived homeostatic temnospondyl. Evolutionary Biology 40: 627–647. Crossref
Sander, P.M. and Wellnitz, P.W. 2024. A phytosaur osteoderm from a late middle Rhaetian bone bed of Bonenburg (North Rhine-Westphalia, Germany): Implications for phytosaur extinction. Fossil Record 27 (1): 147–158. Crossref
Sander P.M. and Wintrich T. 2021. Sauropterygia: histology of Plesiosauria. In: V. de Buffrénil, A.J. de Ricqlès, L. Zylberberg, and K. Padian (eds.), Vertebrate Skeletal Histology and Paleohistology, 444–455. CRC Press, Paris. Crossref
Sander, P.M., Wintrich, T., Schwermann, A.H., and Kindlimann, R. 2016. Die paläontologische Grabung in der Rhät-Lias-Tongrube der Fa. Lücking bei Warburg-Bonenburg (Kr. Höxter) im Frühjahr 2015. Geologie und Paläontologie in Westfalen 88: 11–37.
Säve-Söderbergh, G. 1935. On the dermal bones of the head in labyrinthodont stegocephalians and primitive Reptilia, with special reference to Eotriassic stegocephalians from East Greenland. Meddelelser om Grønland 98 (3): 1–211.
Schobben, M.A.N., Gravendyck, J., Mangels, F., Struck, U., Bussert, R., Kürschner, W.M., Korn, D., Sander, P.M., and Aberhan, M. 2019. A comparative study of total organic carbon-δ13C signatures in the Triassic–Jurassic transitional beds of the Central European Basin and western Tethys shelf seas. Newsletter on Stratigraphy 52: 461–486. Crossref
Schoch, R.R. 1999. Comparative osteology of Mastodonsaurus giganteus (Jaeger, 1828) from the Middle Triassic (Lettenkeuper: Longobardian) of Germany (Baden-Württemberg, Bayern, Thüringen). Stuttgarter Beiträge zur Naturkunde, Serie B 278: 1–175.
Schoch, R.R. 2008. The Capitosauria (Amphibia): characters, phylogeny, and stratigraphy. Palaeodiversity 1: 189–226.
Schoch, R.R. 2013. The evolution of major temnospondyl clades: an inclusive phylogenetic analysis. Journal of Systematic Palaeontology 11: 673–705. Crossref
Schoch, R.R. 2014. Life cycles, plasticity and palaeoecology in temnospondyl amphibians. Palaeontology 57: 517–529. Crossref
Schoch, R.R. and Milner, A.R. 2000. Stereospondyli. In: P. Wellnhofer (ed.), Encyclopedia of Paleoherpetology, Part 3B, 1–203. Verlag Dr. Friedrich Pfeil, Munich.
Schoch, R.R. and Milner, A.R. 2014. Temnospondyli I. In: H.D. Sues (ed.), Encyclopedia of Paleoherpetology, Part 3A2, 1–150. Verlag Dr. Friedrich Pfeil, Munich.
Schoch, R.R. and Rubidge, B.S. 2005. The amphibamid Micropholis stowi from the Lystrosaurus Assemblage Zone of South Africa: Journal of Vertebrate Paleontology 25: 502–522. Crossref
Schoch, R.R. and Witzmann, F. 2012. Cranial morphology of the plagiosaurid Gerrothorax pulcherrimus as an extreme example of evolutionary stasis. Lethaia 45: 371–385. Crossref
Schoch, R.R., Milner, A.R., and Witzmann, F. 2014. Skull morphology and phylogenetic relationships of a new Middle Triassic plagiosaurid temnospondyl from Germany, and the evolution of plagiosaurid eyes. Palaeontology 57: 1045–1058. Crossref
Schoch, R.R., Seegis, D., and Mujal, E. 2022. The Middle Triassic vertebrate deposits of Kupferzell (Germany): Palaeoenvironmental evolution of complex ecosystems. Palaeogeography, Palaeoclimatology, Palaeoecology 603: 111181. Crossref
Schwermann, A.H. 2016. Über den Nachweis von Lepagia gaumensis (Eucynodonta incertae sedis) aus dem Rhät (Trias) von Warburg-Bonenburg (Kr. Höxter). Geologie und Paläontologie in Westfalen 88: 35–44.
Sengupta, D.P. 1995. Chigutisaurid temnospondyls from the Late Triassic of India and review of the family Chigutisauridae. Palaeontology 38: 313–339.
Shishkin, M.A. 1964. Suborder Stereospondyli [in Russian]. In: Y.A. Orlov (ed.), Osnovy paleontologii. Zemnovodnye, presmykaûŝiesâ i ptitsy, 83–122. Izdatel’stvo Nauka, Moskva.
Shishkin, M.A. 1973. The morphology of the early Amphibia and some problems of lower tetrapod evolution [in Russian]. Trudy Paleontologičeskogo Instituta Akademiâ Nauk SSSR 137: 1–257.
Shishkin, M.A. 1986. New data on plagiosaurs from the Triassic of the USSR [in Russian]. Bûlleten’ Moskovskogo Obŝestva Ispytatelej Prirody, Otdel Geologičeskij 61: 97–102.
Shishkin. M.A. 1987. The evolution of early amphibians [in Russian]. Trudy Paleontologičeskogo Instituta, Akademiâ Nauk SSSR 225: 3–142.
Shishkin, M.A. 1991. A Late Jurassic labyrinthodont from Mongolia. Paleontological Journal 1991: 78–91.
Spielmann, J.A. and Lucas, S.G. 2012. Tetrapod fauna of the Upper Triassic Redonda Formation of east-central New Mexico: the characteristic assemblage of the Apachean land-vertebrate faunachron. New Mexico Museum of Natural History and Science Bulletin 55: 1–119.
Steyer, J.S. and Damiani, R. 2005. A giant brachyopoid temnospondyl from the Upper Triassic or Lower Jurassic of Lesotho. Bulletin de la Societe géologique de France 176: 243–248. Crossref
Steyer, J.S., Damiani, R., Sidor, C.A., O’Keefe, F.R., Larsson, H.C.E., Maga, A., and Ide, O. 2006. The vertebrate fauna of the Upper Permian of Niger. IV. Nigerpeton ricqlesi (Temnospondyli: Cochleosauridae), and the edopoid colonization of Gondwana. Journal of Vertebrate Paleontology 26: 18–28. Crossref
Storrs, G.W. 1994. Fossil vertebrate faunas of the British Rhaetian (latest Triassic). Zoological Journal of the Linnean Society 112: 217–259. Crossref
Sulej, T. 2007. Osteology, variability, and evolution of Metoposaurus, a temnospondyl from the Late Triassic of Poland. Palaeontologia Polonica 64: 29–139.
Sulej, T. and Majer, D. 2005. The temnospondyl amphibian Cyclotosaurus from the Upper Triassic of Poland. Palaeontology 48: 157–170. Crossref
Szulc, J., Racki, G., Jewuła, K., and Środoń, J. 2015. How many Upper Triassic bone-bearing levels are there in Upper Silesia (southern Poland)? A critical overview of stratigraphy and facies. Annales Societatis Geologorum Poloniae 85: 587–626. Crossref
Tanner, L.H., Lucas, S.G., and Chapman, M.G. 2004. Assessing the record and causes of Late Triassic extinctions. Earth-Science Reviews 65: 103–139. Crossref
Teschner, E.M., Sander, P.M., and Konietzko-Meier, D. 2018. Variability of growth pattern observed in Metoposaurus krasiejowensis humeri and its biological meaning. Journal of Iberian Geology 44: 99–111. Crossref
Teschner, E.M., Garbay, L., Janecki, P., and Konietzko-Meier, D. 2023. Palaeohistology helps reveal taxonomic variability in exceptionally large temnospondyl humeri from the Upper Triassic of Krasiejów, SW Poland. Acta Palaeontologica Polonica 68: 63–74. Crossref
Vajda, V. and Wigforss-Lange, J. 2009. Onshore Jurassic of Scandinavia and related areas. GFF 131: 5–23. Crossref
Vickers-Rich, P. 1996. Early Cretaceous polar tetrapods from the great southern rift valley, southeastern Australia. Memoirs-Queensland Museum 39: 719–724.
Warren, A.A. 1981. A horned member of the labyrinthodont superfamily Brachyopoidea from the early Triassic of Queensland. Alcheringa 5: 273–288. Crossref
Warren, A.A. and Davey, L. 1992. Folded teeth in temnospondyls—a preliminary study. Alcheringa 16: 107–132. Crossref
Warren, A.A. and Hutchinson, M.N. 1983. The last labyrinthodont? A new brachyopoid (Amphibia, Temnospondyli) from the early Jurassic Evergreen Formation of Queensland, Australia. Philosophical Transactions of the Royal Society of London. B, Biological Sciences 303: 1–62. Crossref
Warren, A.A., Rich, T.H., and Vickers-Rich, P. 1997. The last last labyrinthodonts? Palaeontographica 247: 1–24. Crossref
Watson, D.M.S. 1919. The structure, evolution and origin of the Amphibia. The “orders” Rachitomi and Stereospondyli. Philosophical Transactions of the Royal Society of London B 209: 1–73. Crossref
Watson, D.M.S. 1958. A new labyrinthodont (Paracyclotosaurus) from the Upper Triassic of New South Wales. Bulletin of the British Museum of Natural History, London (Geology) 3: 233–263.
Welles, S.P. 1947. Vertebrates from the upper Moenkopi Formation of northern Arizona. University of California Publications in Geological Sciences 27: 241–289.
Werneburg, R., Witzmann, F., and Schneider, J.W. 2019. The oldest known tetrapod (Temnospondyli) from Germany (Early Carboniferous, Viséan). Paläontologische Zeitschrift 93: 679–690. Crossref
Wiffen, J., de Buffrénil, V., de Ricqlès, A., and Mazin, J.M. 1995. Ontogenetic evolution of bone structure in Late Cretaceous Plesiosauria from New Zealand. Geobios 28: 625–640. Crossref
Wintrich, T., Hayashi, S., Houssaye, A., Nakajima, Y., and Sander, P.M. 2017. A Triassic plesiosaurian skeleton and bone histology inform on evolution of a unique body plan. Science Advances 3: e1701144. Crossref
Witzmann, F. 2009. Comparative histology of sculptured dermal bones in basal tetrapods, and the implications for the soft tissue dermis. Palaeodiversity 2: 233–270.
Witzmann, F. and Gassner, T. 2008. Metoposaurid and mastodonsaurid stereospondyls from the Triassic–Jurassic boundary of Portugal. Alcheringa 32: 37–51. Crossref
Witzmann, F. and Soler-Gijon, R. 2010. The bone histology of osteoderms in temnospondyl amphibians and in the chroniosuchian Bystrowiella. Acta Zoologica (Stockholm) 91: 96–114. Crossref
Witzmann, F. and Schoch, R.R. 2024. Osteology and phylogenetic position of Plagiosaurus depressus (Temnospondyli: Plagiosauridae) from the Late Triassic of Germany and the repeated loss of dermal bones in plagiosaurids. Zoological Journal of Linnean Society 202: 1–29. Crossref
Witzmann, F., Sachs, S., and Nyhuis, C.J. 2016. A new species of Cyclotosaurus (Stereospondyli, Capitosauria) from the Late Triassic of Bielefeld, NW Germany, and the intrarelationships of the genus. Fossil Record 19: 83–100. Crossref
Witzmann, F., Scholz, H., Müller, J., and Kardjilov, N. 2010. Sculpture and vascularization of dermal bones, and the implications for the physiology of basal tetrapods. Zoological Journal of Linnean Society 160: 302–340. Crossref
Yates, A.M. and Warren, A.A. 2000. The phylogeny of the “higher” temnospondyls (Vertebrata: Choanata) and its implications for the monophyly and origins of the Stereospondyli. Zoological Journal of Linnean Society 128: 77–121. Crossref
Zittel, K.A. 1888. Handbuch der Paläontologie. Abteilung 1. Paläozoologie Band III. Vertebrata (Pisces, Amphibia, Reptilia, Aves). 900 pp. R. Oldenbourg, Munich.
Acta Palaeontol. Pol. 69 (4): 687–712, 2024
https://doi.org/10.4202/app.01147.2024