
Boreal waterways: An Early Cretaceous plesiosaur from Ellesmere Island, Nunavut, Canadian Arctic and its palaeobiogeography
LENE L. DELSETT, ADAM S. SMITH, STEPHEN INGRAMS, and SIMON SCHNEIDER
Delsett, L.L., Smith, A.S., Ingrams, S., and Schneider, S. 2024. Boreal waterways: An Early Cretaceous plesiosaur from Ellesmere Island, Nunavut, Canadian Arctic and its palaeobiogeography. Acta Palaeontologica Polonica 69 (4): 565–585.
A plesiosaur specimen collected from Ellesmere Island (Nunavut, Arctic Canada) by Danish geologist Johannes Troelsen in 1952 is described for the first time. The plesiosaur is late Berriasian to early Valanginian in age based on palynostratigraphy. The specimen is the only plesiosaur known from the Lower Cretaceous of the Sverdrup Basin in the Canadian Arctic, and is assigned to the cryptoclidid genus Colymbosaurus. From a taxonomic point of view, the presence of vertebrae from several regions and four propodials improve our understanding of the morphology of the genus. Furthermore, Colymbosaurus is shown to have survived through the Jurassic–Cretaceous transition. Its presence in the Sverdrup Basin is additional evidence for the connectivity of Arctic Canada and the Svalbard region during the Jurassic–Cretaceous transition, at a time when sea levels were low and microplankton, like dinoflagellates, experienced enhanced provincialism. Last but not least, the new plesiosaur adds to our knowledge of the palaeoenvironment of the Sverdrup Basin, ranking at the top of a food chain that is largely unrecorded from the area, due to adverse taphonomy and diagenesis.
Key words: Plesiosauria, Colymbosaurus, palaeobiogeography, Early Cretaceous, Sverdrup Basin, Ellesmere Island, Nunavut, Canada.
Lene L. Delsett [lenelie@uio.no; ORCID: https://orcid.org/0000-0002-6806-1411 ], Natural History Museum, University of Oslo, Oslo, Norway.
Adam S. Smith [adam.smith@nottinghamcity.gov.uk; ORCID: https://orcid.org/0000-0002-8166-1139 ], Nottingham Natural History Museum, Wollaton Hall, Nottingham, NG8 2AE, UK.
Stephen Ingrams[Stephen.Ingrams@cgg.com; ORCID: https://orcid.org/0000-0003-2610-8135 ], CGG, Tyn-y-Coed, Pentywyn Road, Llandudno LL30 1SA, UK.
Simon Schneider [simon.schneider@casp.org.uk; ORCID: https://orcid.org/0000-0001-6493-357X ], CASP, West Building, Madingley Rise, Madingley Road, Cambridge CB3 0UD, UK.
Received 27 February 2024, accepted 19 July 2024, published online 2 December 2024.
Copyright © 2024 L.L. Delsett et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
In spring 1952, Danish geologist Johannes Christian Troelsen set out for fieldwork on Fosheim Peninsula, Ellesmere Island, Arctic Canada (Fig. 1). Close to Eureka Weather Station, he collected “parts of the skeleton of a large plesiosaur” from a succession of “soft shales” provisionally assigned to Upper Jurassic to Lower Cretaceous based on the occurrence of fossil bivalves of the genus Buchia, and briefly reported on his find (Troelsen 1952: 208). Further reports regarding the condition of the skeleton and its age are unpublished (Troelsen 1953, 1954), and the plesiosaur remained unstudied. Here we describe the specimen in detail, assess its taxonomy and determine its age.
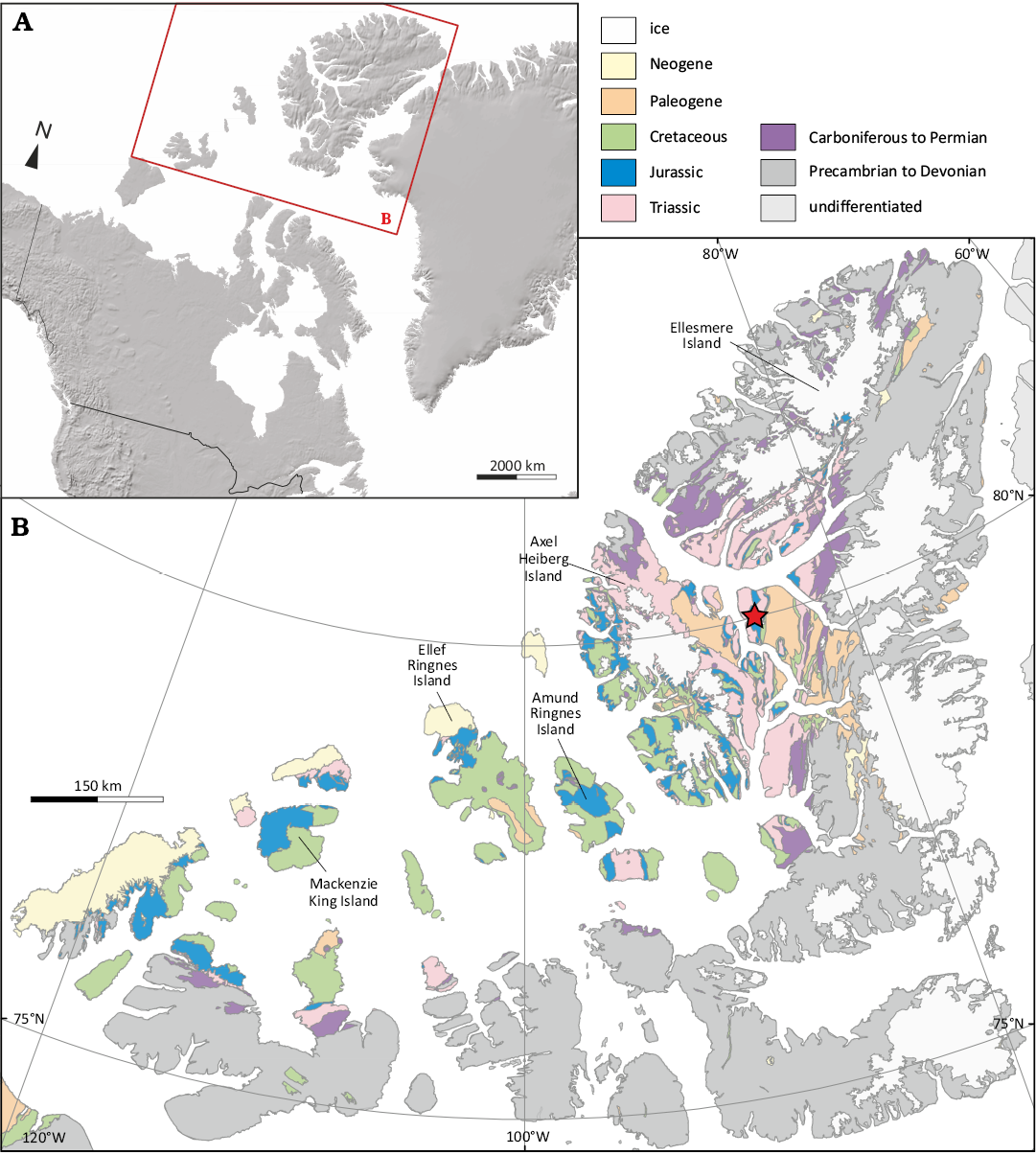
Fig. 1. Geographical and geological overview. A. Location map of the Sverdrup Basin. B. Geology of the Sverdrup Basin (modified from Harrison et al. 2011) and location of the plesiosaur locality on Ellesmere Island (red star; modified from Schneider et al. 2020).
Plesiosaurs are secondarily aquatic sauropterygians with a fossil record ranging from the Triassic to the Cretaceous/Paleogene boundary (Benson et al. 2012; Wintrich et al. 2017; Madzia and Cau 2020). By the Early Jurassic the group had reached a global distribution (Bardet et al. 2014), including high latitudes. Palaeogeographically, this was facilitated by the break-up of Pangea, leading to the opening of the Viking and Hispanic corridors, and by generally high sea levels.
However, sea levels did not remain universally high subsequently, and connectivity between basins varied over time. The Jurassic/Cretaceous transition interval in particular, which is critical for the present study, experienced significant sea level drops (e.g., Van Der Meer et al. 2022), resulting in extinction events in many tetrapod groups (Tennant et al. 2016), and provincialism in marine communities (Mutterlose et al. 2003). Sauropterygians were probably less hard hit by the extinction event at the Jurassic/Cretaceous boundary transition than many other groups, but their particularly poor earliest Cretaceous fossil record prevents an understanding of which plesiosaur lineages actually survived into the Cretaceous (Benson and Druckenmiller 2014; Tennant et al. 2016; Zverkov et al. 2018; Roberts et al. 2020). It seems most plesiosaur genera are endemic at basin scale, despite their swimming abilities (Bardet et al. 2014). However, their migration patterns and distribution potential are also incompletely known, and some genera, such as Libonectes, instead show a wide dispersal (Bardet et al. 2014; Sachs and Kear 2017). The new plesiosaur from Ellesmere Island adds to a sparse fossil record of Early Cretaceous plesiosaurs and comes from a geographic location to help shed light on these topics.
Institutional abbreviations.—NHMD, Natural History Museum of Denmark, University of Copenhagen, Denmark; NMNH/USNM, National Museum of Natural History, Smithsonian Institution, Washington DC, USA; PMO, Natural History Museum palaeontological collection, University of Oslo, Norway; MJML, The Etches Collection, Museum of Jurassic Marine Life, Kimmeridge, UK.
Geological setting
The Sverdrup Basin is located in the Queen Elizabeth Islands, in today’s provinces Nunavut and the Northwest Territories, covering an area of approximately 300 000 km2 (Embry and Beauchamp 2019; Fig. 1). Basin development commenced in the Mississippian on Neoproterozoic to Devonian strata and terminated with the onset of the Eurekan Orogeny in the latest Cretaceous (e.g., Balkwill 1978; Embry and Beauchamp 2019). The Sverdrup Basin is filled with sedimentary strata up to 15 km thick, ranging from the Carboniferous to the Eocene, which conform to eight first-order, unconformity-bounded depositional sequences, as defined by Embry and Beauchamp (2019).
Only Sequence 5, which comprises four second-order transgressive-regressive sequences (Rhaetian–Sinemurian, Pliensbachian–Aalenian, Bajocian–Callovian and Oxfordian–Valanginian), is relevant to the present study. The Deer Bay Formation, which yielded the plesiosaur specimen discussed herein, was deposited during the Tithonian to Valanginian, conforming to the upper part of second-order sequence 5.4 (Embry and Beauchamp 2019; Galloway et al. 2020; Schneider et al. 2020), and conforms to two of 57 tectonically-generated large-magnitude sequences (numbers 41 and 42) sensu Embry et al. (2018) (Fig. 2). Based on a multivariate statistical analysis of dinoflagellate cyst assemblages, the Deer Bay Formation extends over large parts of two third-order sequences, and its upper, Valanginian part can be further subdivided into eight fourth-order sequences (Ingrams et al. 2022). In the study area on Fosheim Peninsula, the Deer Bay Formation conformably overlies sandstones of the Slidre Member of the Awingak Formation (Embry 1986), and is in turn conformably overlain by sandstones of the Isachsen Formation (Thorsteinsson 1971).
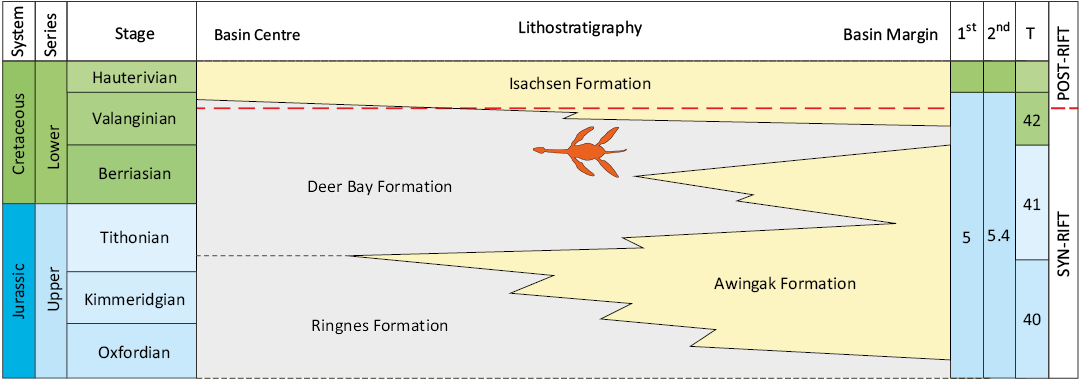
Fig. 2. Oxfordian to Hauterivian lithostratigraphy, sequence stratigraphy and tectonostratigraphy of the Sverdrup Basin. Light yellow and light grey colours indicate sandstone- and mudstone-dominated units, respectively. The approximate stratigraphic position of the plesiosaur is indicated. First and second order transgressive-regressive sequences are taken from Embry and Beauchamp (2019), tectonically-generated sequences (T) from Embry et al. (2018) and tectonostratigraphic phases from Hadlari et al. (2016).
The Deer Bay Formation was redefined by Balkwill (1983), and classified as a succession of dark mudstone and siltstone with rare intercalations of very-fine- to fine-grained sandstone. For Fosheim Peninsula, the total thickness of the Deer Bay Formation is given as 275 m (900 ft) by Thorsteinsson (1971). No detailed log is available for the area, but according to Kemper (1975: fig. 2), the upper, Valanginian part of the succession is approximately 150 m thick.
Material and methods
Specimen history.—The plesiosaur NHMD 189834 was found approximately two miles (= 3.2 km) away from Eureka Weather Station (see dotted-line circle segment on Fig. 3; Troelsen 1952). In this area, a succession of mudstones of the Deer Bay Formation is poorly exposed at the slopes of an initially unnamed creek (Troelsen 1952; Thorsteinsson 1971), christened Reptile Creek by Troelsen (1953, 1954). Later, this name was adopted in the literature (Jeletzky 1966; Kemper 1975). Jeletzky (1979, 1984), who never visited the area, seems to have confused Reptile Creek and Station Creek (named Weather Station Creek in Jeletzky 1979: 28, and Camp Creek in Jeletzky 1984: 221). Based on the coordinates provided by Kemper (1975), Reptile Creek is clearly the eastern tributary of Station Creek, as marked in Fig. 3. Troelsen briefly mentioned the plesiosaur specimen in his paper (Troelsen 1952) and in two unpublished reports (Troelsen 1953, 1954). These texts include no description of the plesiosaur fossils, and no log of the respective strata.
The plesiosaur specimen was sent to the University of Copenhagen, where a student of curator Eigil Nielsen sorted (Troelsen 1953) and probably also cleaned and glued several of the fossil skeletal elements. It was probably the subsequent curator, Niels Bonde, who later marked the vertebrae, ribs, and transverse processes with letters and numbers. Subsequently, the bones were packed into several boxes and wooden crates. The contents of one of these boxes, comprising several vertebrae, distal parts of a forelimb and other isolated limb elements, one complete pelvic rib, and more than a hundred fragments of ribs and girdle elements, were studied by co-author ASS in November 2005. Possibly during the flooding of the museum in 2011, this box got misplaced, but was located again by curators Bent Lindow and Arden Bashforth; its contents were subsequently stored in drawers in the main fossil vertebrate collection. Two additional wooden crates marked “Reptile Creek, Troelsen’s office” were rediscovered in 2019, and contained all four propodials and several girdle elements of the plesiosaur specimen. A third assemblage of skeletal elements from the same plesiosaur specimen was discovered by LLD in the collections of the Zoological Museum in 2020. These were mislabelled as part of a “Scoresbysund plesiosaur”, likely when specimens had to be moved during the flooding of the collections in 2011, but clearly belong to NHMD 189834. Another box labelled “NHMD 189689”, containing fragmentary ribs that closely resemble those of plesiosaur NHMD 189834, was also found in the collections of the Zoological Museum, but has no documentation, so it is not included in this study.
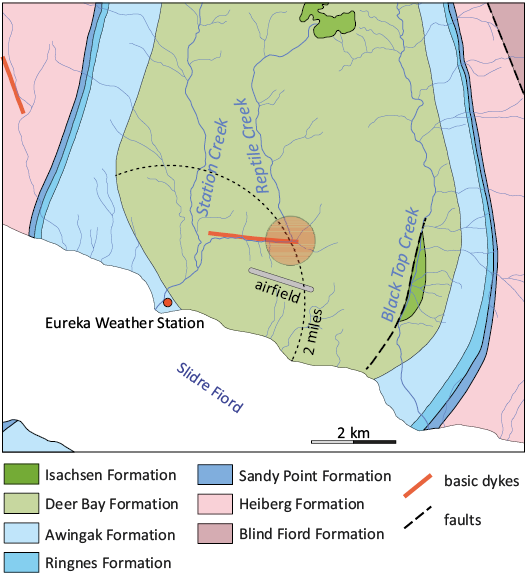
Fig. 3. Geological map of the area around Eureka Weather Station north of Slidre Fiord (modified from Thorsteinsson 1971). The approximate area where the plesiosaur was found is indicated by an orange circle. The dotted-line circle segment indicates Deer Bay Formation exposure at two miles radius from Eureka Weather station.
Methods.—NHMD 189834 was studied at the Natural History Museum in Copenhagen by ASS in 2005 (parts only) and by LLD in 2020 and 2022; SS assisted with photography. Measurements were taken with manual callipers and a tape measure.
Letters and numbers written on the vertebrae, ribs, and transverse processes by the previous curator are used in this paper. None of the limb elements have letters or numbers.
Plesiosaur specimens Tatanectes laramiensis (PAL 536970, PAL 536971), Pantosaurus striatus (PAL 536973, USNM 5782, 536965, 418494, 536963), Cryptoclidus sp. (USNM 419630), plesiosaur indet. (PAL 536974, USNM 536977a) were studied in the collections of NMNH, autumn 2021 by LLD for comparison.
Two samples of matrix sediment adhering to the plesiosaur bones were removed using a pneumatic chisel, and processed for palynological analysis, following the standard procedures outlined in Ingrams et al. (2021). The recovered palynomorph assemblages were screened for biostratigraphically relevant dinoflagellate cyst, pollen and spore taxa.
Phylogenetic analysis.—In order to determine the phylogenetic position of NHMD 189834 and evaluate the taxonomic placement as suggested by morphological traits, a phylogenetic analysis was run in TNT 1.6 (Goloboff and Morales 2023), using the matrix from Sachs et al. (2024) with an additional operational taxonomic unit (OTU) and scores added for NHMD 189834. As in Sachs et al. (2024), Neusticosaurus pusilllus was used as the outgroup, and analyses were performed using the same searches and parameters. This included “New technology” searches with different values of K (6, 9, and 12), and analyses equally weighted and implied weighting of characters, as well as “traditional search” with TBR (Sachs et al. 2024). For NHMD 189834, 63 out of 270 characters (23%) could be scored with confidence (SOM, Supplementary Online Material available at http://app.pan.pl/SOM/app69-Delsett_etal_SOM.pdf).
Biostratigraphy
Macrofauna.—Troelsen (1952) collected Buchia from the lower part of the Deer Bay Formation, and ammonites and belemnites from its upper part. These fossils are now curated at the Geological Survey of Canada, and their precise stratigraphic relationships to the plesiosaur specimen are unclear. Jeletzky identified Buchia cf. terebratulides (Lahusen, 1888), B. volgensis (Lahusen, 1888) and B. keyserlingi (Trautschold, 1868) from Troelsen’s material (Troelsen 1953), which are indicative of late Tithonian to early Berriasian, early to late Berriasian and early Valanginian ages, respectively. Ammonites collected by Troelsen were collectively determined as Tollia cf. tolli (Pavlow, 1914) by C.W. Wright (Troelsen 1953), but later reassigned to Temnoptychites (Costamenjaites) troelseni (Jeletzky, 1979), Temnoptychites (C.) grandiosus (Voronets, 1962), Temnoptychites (C.) aff. grandiosus (Voronets, 1962), Tollia aff. tolli Pavlow, 1914, and Tollia aff. klimovskiensis Krimgolts, 1953 [sic!] (in Krymgolts et al. 1953) by Jeletzky (1979). This assemblage is latest Berriasian in age and correlates to the Tollia tolli Zone of northern Siberia (cf. Wierzbowski et al. 2011). Collectively, the macrofauna indicates a late Tithonian to early Valanginian age for the Deer Bay Formation north of Slidre Fiord.
Palynoflora.—Part of the dinoflagellate cyst Oppel zonation established for the Sverdrup Basin by Davies (1983) was revised by Ingrams et al. (2021). Based on the taxon ranges observed in the latter study, the three species Prolixosphaeridiopsis spissa, Trichodinium erinaceoides, and Oligosphaeridium complex define a latest Berriasian to early Valanginian age for the sediment adhering to the plesiosaur bones. Terrestrial palynomorphs (Microreticulatisporites diatretus Norris, 1969, Deltoidospora fsp., Sigmopollis callosus Norris, 1969, Undulatasporites araneus Norris, 1969, Gleicheniidites minor Döring, 1965, Gleicheniidites delcourti Döring, 1965) (Döring 1965; Norris 1969) indicate an even narrower age bracket around the Berriasian–Valanginian transition. Given that the zonation schemes in use are still based on very few data points, these results should be interpreted with a certain degree of caution. Nonetheless, biostratigraphy supports a latest Berriasian to earliest Valanginian age for the plesiosaur specimen (Fig. 4).
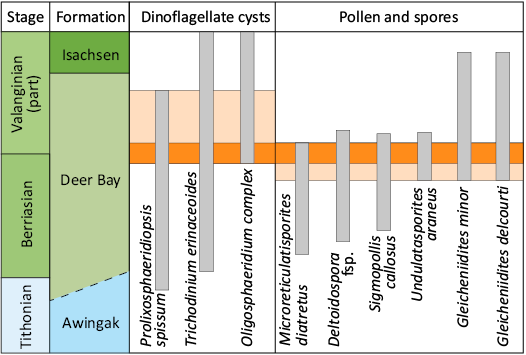
Fig. 4. Chronostratigraphic ranges of biostratigraphically significant dinoflagellate cyst, pollen and spore taxa, recovered from the studied samples.
Systematic palaeontology
Sauropterygia Owen, 1860
Plesiosauria de Blainville, 1835
Plesiosauroidea Welles, 1943
Cryptoclididae Williston, 1925
Genus Colymbosaurus Seeley, 1874
Type species: Colymbosaurus megadeirus Seeley, 1869; Kimmeridgian–lower Tithonian, Kimmeridgian Clay Formation in Haddenham and Ely, Cambridgeshire, UK.
Colymbosaurus sp.
Figs. 5–7, 9.
Material.—NHMD 189834, a partial skeleton consisting of 22 non-consecutive vertebrae from the cervical, dorsal, and caudal regions, hundreds of rib fragments, partial girdle elements, all four propodials, and several distal limb elements from Reptile Creek, approximately 3 km away from Eureka Weather station, Ellesmere Island, Nunavut, Canada, uppermost Berriasian to lowermost Valanginian.
Description.—Preservation: NHMD 189834 preserves all four propodials along with several limb elements, partial girdle elements, 22 vertebrae and numerous partial ribs. The elements are preserved as three-dimensional, mostly undistorted bones, and many of them show well-preserved surface details and facets. However, all of the bones are weathered, and many are incomplete or preserved in many fragments. The propodials have been partly reconstructed using plaster, but not to an extent that makes descriptions misleading.
Ontogeny: Where preserved, the neural arches are fused to the vertebral centra, indicating that the individual was osteologically mature (Araújo and Smith 2023), and therefore an “adult” as per the criteria of (Brown 1981). In addition, the relatively coarse, finished bone outer layer, as well as the large size of the skeletal elements and the well-defined distal facets on all propodials are indicative of adulthood.
Body size: The humeri of NHMD 189834 are 390 mm (left) and 400 mm (right) long, and the left and right femora are 350 mm and 360 mm long, respectively. All of these bones are slightly shorter than the respective elements in the three specimens of Colymbosaurus svalbardensis from the Slottsmøya Member (Knutsen et al. 2012c; Roberts et al. 2017), and the femora are slightly shorter than in Colymbosaurus megadeirus (CAMSMJ.29596, etc.) from the UK (Benson and Bowdler 2014), but fit within the large and small size classes of Russian colymbosaurines (Arkhangelsky et al. 2020). In the specimen of C. megadeirus, the neck length was estimated to be 2 m and the trunk 1.7 m (Benson and Bowdler 2014), so, based on femoral length, the Ellesmere Island specimen would have been somewhat shorter than that.
Axial skeleton: In total, 22 non-consecutive vertebrae are preserved (Fig. 5). 21 vertebrae are complete or almost complete centra, and some have partial neural arches preserved intact. One vertebra is represented by only a small fragment. Sachs et al. (2013) is used for referring to the regions of the axial skeleton. As the specimen is neither complete nor articulated, the total number of vertebrae is unknown for the different body regions. Three cervical centra are preserved (Fig. 5A), and since this was a long-necked species, this means the vast majority are missing. Two pectoral vertebrae are preserved (Fig. 5B, C). Two dorsal vertebrae are preserved with an intact transverse process on one side, but these processes have broken off the other vertebrae from the same region. Five centra were too weathered or incomplete to be assigned to any specific vertebral region. In addition to the centra, several isolated transverse process portions are preserved. There are also five large disarticulated neural arches, but which region of the vertebral column they belong to is unknown. Centrum measurements of the preserved vertebrae are provided in Table 1.
Table 1. Vertebrae measurements (in mm). Names of vertebrae refer to letters written on the fossils. NA, not available.
|
Vertebra |
Region |
Dorsoventral |
Anteroposterior length |
Maximum mediolateral width |
|
Centrum A |
cervical |
NA |
55 |
NA |
|
Centrum B |
cervical |
NA |
59 |
NA |
|
Centrum C |
cervical |
61 |
53 |
83 |
|
Centrum D |
pectoral |
65 |
48 |
74 |
|
Centrum E |
pectoral |
NA |
52 |
73 |
|
ARROW |
dorsal |
60 |
59 |
NA |
|
Centrum G |
dorsal |
68 |
57 |
72 |
|
Centrum R |
dorsal |
NA |
NA |
NA |
|
no marking |
dorsal |
73 |
60 |
72 |
|
Centrum H |
dorsal |
66 |
57 |
75 |
|
Centrum I |
dorsal |
72 |
67 |
67 |
|
Centrum K |
dorsal |
69 |
65 |
73 |
|
Centrum T |
dorsal |
63 |
64 |
79 |
|
Centrum M |
dorsal |
69 |
63 |
71 |
|
Centrum L |
dorsal |
69 |
68 |
78 |
|
Centrum O |
caudal |
61 |
46 |
71 |
|
Centrum P |
caudal |
55 |
40 |
67 |
|
Centrum F |
NA |
NA |
NA |
72 |
|
Centrum N |
NA |
NA |
63 |
NA |
|
Centrum S |
NA |
NA |
54 |
NA |
|
small piece |
NA |
NA |
NA |
NA |
|
tiny, eroded centrum |
NA |
NA |
NA |
NA |
Cervical vertebrae: Three cervical vertebrae are preserved in NHMD 189834, all missing the neural arch and neural spine (Fig. 5A). As they were disarticulated, it is not known for certain which ones are from the more anterior position in the neck. One cervical centrum is complete (centrum C). It is shorter anteroposteriorly than tall dorsoventrally, as in Colymbosaurus megadeirus, which is slightly shorter than tall (Benson and Bowdler 2014), Muraenosaurus sp. specimen MUHNCAL.20176 (Otero et al. 2020), and Tatanectes laramiensis (O’Keefe and Street 2009; USNM 536970, LLD personal observations). However, the latter is significantly shorter anteroposteriorly than NHMD 189834. This contrasts with Spitrasaurus wensaasi and Pantosaurus striatus (USNM 5782, LLD personal observations), where the cervical vertebrae are longer than tall, and S. larseni and Abyssosaurus nataliae, which have cervical centra that are equally long and tall (Berezin 2011; Knutsen et al. 2012d). However, in Ophthalmothule cryostea, which preserves a complete neck, the relative measurements of the vertebrae vary between different parts of the neck (Roberts et al. 2020), which might also be the case for NHMD 189834. The cervical centrum is also significantly wider mediolaterally than tall dorsoventrally, as in Colymbosaurus megadeirus (Benson and Bowdler 2014), Muraenosaurus leedsi (Andrews 1910), Abyssosaurus nataliae (Berezin 2011), Spitrasaurus larseni (Knutsen et al. 2012d) and Pantosaurus striatus (USNM 5782 and 536963, LLD personal observations; Wilhelm and O’Keefe 2010). The vertebral length index is 0.74, similar to the posterior vertebrae of C. megadeirus (Benson and Bowdler 2014). For C. svalbardensis, cervical vertebrae are not known (Roberts et al. 2017).
On the dorsal surface, the neural canal is equally wide mediolaterally for its entire length, as also figured for C. megadeirus (Benson and Bowdler 2014), but in contrast to Spitrasaurus wensaasi, where it is hour-glass shaped (Knutsen et al. 2012d). Two anteroposteriorly elongated foramina are situated on the floor of the neural canal, as in Spitrasaurus wensaasi (Knutsen et al. 2012d) and Ophthalmothule cryostea (Roberts et al. 2020).
Two of the cervical centra are platycoelous (centrum A and B), with one side flat and the other only very slightly concave, whereas the other centrum (centrum C) is amphicoelous with two slightly concave surfaces. The latter morphology is the same as in the cervical centra of C. megadeirus, Spitrasaurus wensaasi, S. larseni, and Abyssosaurus nataliae (Berezin 2011; Knutsen et al. 2012d; Benson and Bowdler 2014), in contrast to Cryptoclidus eurymerus, where all centra are amphicoelous (Brown 1981). A groove around the perimeter of the articular faces, as described in Colymbosaurus megadeirus (Benson and Bowdler 2014), is absent. The cervical centrum is spool shaped, with expanded articular ends compared to the main body of the centrum.

Fig. 5. Selected vertebrae of plesiosaur Colymbosaurus sp. NHMD 189834 from Ellesmere Island, Nunavut, Canada, upper Berriasian–lower Valanginian. A. Cervical vertebra (centrum C), in articular (A1), dorsal (A2), ventral (A3), and lateral (A4) views. B, C. Pectoral vertebrae. B. Centrum E in articular view. C. Centrum D, in articular (C1) and lateral (C2) views. D. Sacral? rib. E–G. Dorsal vertebrae. E. Centrum H, in articular (E1) and lateral (E2) views. F. Centrum M (dorsal?) in articular view. G. Centrum I in articular view. H, I. Caudal vertebrae. H. Centrum O in articular view. I. Centrum P in articular (I1) and lateral (I2) views. The letters used in the element names are written on the individual elements.
The lateral surface of the centrum lacks a longitudinal ridge, as it does in Colymbosaurus megadeirus (Benson and Bowdler 2014), Spitrasaurus wensaasi and S. larseni (Knutsen et al. 2012d). However, as only three cervical centra are preserved it is possible that a lateral ridge was present in other centra. The cervical rib facet is single headed and situated on the ventralmost portion of the lateral surface of the centrum (Fig. 5A4). This is similar to the situation in Muraenosaurus leedsi, Colymbosaurus megadeirus, Pantosaurus striatus (USNM 5782, LLD personal observations) and Spitrasaurus wensaasi (Andrews 1910; Knutsen et al. 2012d; Benson and Bowdler 2014). The facet is anteroposteriorly longer than dorsoventrally tall, as in Spitrasaurus wensaasi (Knutsen et al. 2012d). The facet does not contact the anterior or posterior margin of the centrum. The facet in centrum C is fused to the cervical rib.
On the ventral surface, two nutritive foramina are positioned posteriorly. These are anteroposteriorly oriented ovals in centrum A, but circular in centrum C (Fig. 5A3). They are widely spaced, several centimetres apart, in contrast to Colymbosaurus megadeirus (Benson and Bowdler 2014) and Spitrasaurus wensaasi, where the foramina are positioned close to each other (Knutsen et al. 2012d), even more so than in Tatanectes laramiensis (O’Keefe and Street 2009). The ventral surface is almost flat, with a slight bulge between the foramina. Spitrasaurus wensaasi has a keel running along the entire ventral surface (Knutsen et al. 2012d), whereas Tatanectes laramiensis has no ventral keel (O’Keefe and Street 2009).
Pectoral vertebrae: Two vertebrae are interpreted as pectoral vertebrae (Fig. 5B, C), because their rib facets are dorsoventrally tall and contact the base of the neural arch. Alternatively, they might represent sacral vertebrae, which have a rather similar morphology. Their centra are mediolaterally wider than dorsoventrally tall as in Colymbosaurus megadeirus and Ophthalmothule cryostea (Benson and Bowdler 2014; Roberts et al. 2020), but not to such an extent as in the cervicals. The articular surface is only slightly concave. As in the cervical vertebrae, the neural canal is evenly mediolaterally wide. On the left lateral side of one vertebra (centrum D) is an anterodorsally oriented ridge that connects the rib facet and the neural arch base. This morphology is also found in the pectoral and sacral vertebrae of Colymbosaurus megadeirus and Ophthalmothule cryostea (Benson and Bowdler 2014; Roberts et al. 2020). The paired foramina on the ventral surface are more widely spaced than in the cervical centra, so they are also visible in lateral view. The distribution of foramina is also asymmetrical, with an additional foramen on the right side of centrum D. The rib facets are incomplete but confirm that the centrum had single-headed ribs.
Dorsal vertebrae: The ten dorsal vertebrae are determined based on the more or less circular outline of the articular surfaces of the centra, the lack of facets for ribs on the centrum body and, where preserved, a neural arch that bears dorsolaterally directed transverse processes for articulation with the dorsal ribs (Sachs et al. 2013). Except for one centrum (centrum L, anteroposterior length : dorsoventral height = 1.01), all are slightly dorsoventrally taller than anteroposteriorly long, as in Colymbosaurus megadeirus (Benson and Bowdler 2014). Most of the dorsal centra are also slightly dorsoventrally taller than mediolaterally wide, whereas some are slightly wider than tall, as in C. svalbardensis (Roberts et al. 2017). The dorsal centra are platycoelous with a completely flat anterior articular surface, and an almost flat but slightly concave posterior articular surface. They are strongly spool shaped, with articular ends mediolaterally wider than the main body of the centrum. In addition, some centra have a small ridge or lip around the articular margins where the centrum becomes mediolaterally wider still. A few possess a small knob in the centre of the articular face. Vertebrae ARROW, G, and R probably belong to the anteriormost portion of the dorsal series. In these, the floor of the neural canal widens at both ends, in contrast to most others, where it is evenly wide throughout. In one centrum (centrum M), the floor of the neural canal narrows at one end.
Three centra preserve one or both of their transverse processes (centra H, I, and L); one of these processes is almost complete (centrum I). The neural canal in the dorsal vertebrae is significantly taller than wide and is dorsoventrally ovate in cross-section. In centrum I, the most complete dorsal vertebra, it is 30 mm tall, twice the maximum mediolateral width of the canal (Fig 5G). This character has been suggested as an autapomorphy of C. svalbardensis (Roberts et al. 2017), differentiating it from Kimmeridge Clay Formation cryptoclidids. However, although Benson and Bowdler (2014) figure a plesiosauroid vertebra from the Kimmeridge Clay (OXFUM J.55491) with a circular dorsal neural canal to show the possible anatomy in C. megadeirus, the dorsal canal is not known for certain in C. megadeirus. In C. megadeirus, the neural canal in the posterior cervical vertebrae is dorsoventrally tall, and one dorsal vertebra also appears to show a dorsoventrally tall neural canal (Benson and Bowdler 2014; Fig 6F); however, this vertebra is distorted, and the tall canal could be an artefact of crushing. The ventral portion of the neural canal is wider than the top, exactly as seen in C. svalbardensis (Knutsen et al. 2012c). In Ophthalmothule cryostea, the neural canal is oval in anterior view (Roberts et al. 2020).
A cross-section through the neural arch pedicles is visible in centrum R. These are mediolaterally widest about 1/3 from the anterior margin where they form a triangular buttress below the transverse process. They taper anteriorly to a sharp anterior edge, and posteriorly to a flat posterior edge.
The transverse processes are long, gracile, and inclined dorsolaterally and slightly posteriorly, as in C. svalbardensis (Knutsen et al. 2012c; Benson and Bowdler 2014; Roberts et al. 2017). The distal extremity is expanded dorsoventrally to form an anterodorsally oriented facet that faces slightly posterolaterally. The rib facet is oval, as in C. svalbardensis and Tatanectes laramiensis (USNM 536974, LLD personal observations; Knutsen et al. 2012c). The bases of the transverse processes are oriented anteroposteriorly, and cover the entire length of the centrum except for a short anterior portion. Where it can be evaluated, a transverse subdiapophyseal fossa, such as the one shown in Hampe (2013), is absent. The arrangement of foramina is asymmetrical in some of the dorsal centra and at least two centra (centra G and M) have up to six small foramina on the base of the neural canal. On the ventralmost portion of the lateral side the dorsal centra have up to five anteroposteriorly elongated foramina, a condition also observed in anterior dorsal vertebrae in Tatanectes laramiensis (USNM 536974, LLD personal observations). For example, in centrum K, there are five clear foramina on the left side of the centrum, but only one foramen on the right side.
In lateral view, several of the dorsal vertebrae possess one articular margin that extends more ventrally compared to the other articular margin. In the holotype of C. svalbardensis (PMO A27745), a similar condition, termed “anterior lip on the ventral margin”, was observed in one dorsal vertebra and interpreted as a taphonomical artefact (Knutsen et al. 2012c). However, as it is observed in several dorsal centra of the studied specimen we here interpret it as a genuine morphological feature in some of the dorsal vertebrae in this species.
Caudal vertebrae: Two caudal vertebrae (O and P) are preserved, but are missing the neural arch and spine. The caudal rib facets and chevron facets contribute to an almost pentagonal outline in articular view. In Abyssosaurus nataliae the shape is hexagonal (Berezin 2011). The caudal vertebrae in NHMD 189834 are mediolaterally wider than dorsoventrally tall, as in C. svalbardensis and C. megadeirus (Benson and Bowdler 2014; Roberts et al. 2017), and anteroposteriorly shorter than dorsoventrally tall, but not as tall as in C. megadeirus (Benson and Bowdler 2014).
The caudal vertebrae bear processes for the caudal ribs positioned approximately at mid height on the lateral surface of the centrum, ventral to and disconnected from the neural arch, as in most cryptoclidids (Benson and Bowdler 2014). The rib facets are also positioned midway anteroposteriorly. The rib facets are subtriangular ovals with a flat to concave anterior surface. This feature is known to vary along the caudal series in C. svalbardensis (Knutsen et al. 2012c).
On the ventrolateral corners are small chevron facets that are triangular in ventral and lateral views. The posterior facets are significantly larger than the anterior facets. Triangular chevron facets are also found in C. svalbardensis but differ from C. megadeirus, which has oval chevron facets (Knutsen et al. 2012c; Benson and Bowdler 2014; Roberts et al. 2017). The chevron facets are significantly smaller than the caudal rib facets. The ventral surface of the centrum is almost flat and bears four elongated foramina arranged in two pairs, one in front of the other.
The neural arch pedicels appear to have been fused to the centrum but have broken off at their base. They are mediolaterally narrow for their entire length, and the floor of the neural canal is equally mediolaterally wide for its entire length. The pedicels extend anteriorly to the anterior edge of the centrum but terminate posteriorly well before the posterior edge of the centrum.
Ribs and gastralia: Approximately 100 rib fragments and gastralia are preserved, but none are complete. The longest pieces are 150 mm. It is possible that some pieces may match up. Some elements preserve the proximal ends of dorsal ribs including the articulation facets. The facets are oval with concave articular faces, and the main part of the ribs are flattened. Fragments of other ribs are larger and more robust, and most likely belong in the dorsal section. Their cross-sections vary greatly in outline, from oval to almost circular, and from nearly figure-of-eight-shaped to thickened T-shaped. As discussed for C. svalbardensis (Roberts et al. 2017), cross sections vary along the length of the ribs. One complete and one incomplete sacral rib are preserved (Fig. 5D). These elements are slenderer than the dorsal ribs. The proximal articular facet is trilobate in proximal view, anteroposteriorly widest ventrally. The single facet is divided into distinct upper and lower parts, with the slightly smaller upper part inclined about 40° relative to the larger ventral part. The distal end of the rib is dorsoventrally expanded into a rounded bulb, but it is not expanded anteroposteriorly. Although both ends of the rib are dorsoventrally expanded, the proximal end of the sacral rib is dorsoventrally much thicker than the distal end.
Girdle elements: One incomplete element is large, flat and 1–2 cm thick, but partly covered with matrix (Fig. 6A). It was found attached to one of the humeri, and although this was probably not its real position in life, this proximity suggests this is part of the pectoral girdle, probably the coracoid. The element has a partially preserved outline including a long gently concave margin, which might represent the lateral margin of the coracoid. A tapering, slightly squared-off end could represent the posterolateral cornua of the coracoid.

Fig. 6. Girdle elements of plesiosaur Colymbosaurus sp. NHMD 189834 from Ellesmere Island, Nunavut, Canada, upper Berriasian–lower Valanginian. A. Coracoid? B. Scapula? C. Partial coracoid or clavicle.
One incomplete fan shaped element is interpreted as a partial scapula (Fig 6B). The posterior process with the glenoid contribution is preserved, whereas the anterior outline is incomplete, so that the full size and outline of the bone is unknown. The process is thicker and more robust than the rest of the element, as in Spitrasaurus wensaasi (Knutsen et al. 2012d). The glenoid contribution is complete and has an almost circular cross-section. The process with the glenoid contribution is offset ventrally from the plate of the scapula, likely medially and ventrally.
A separate, small fragment has a tapering shape that resembles the posterolateral cornua of the other coracoid, or the lateral part of a clavicle (Fig. 6C). For example, the shape of this element matches the posterolateral cornua of C. svalbardensis figured by Roberts et al. (2017: fig 8A, B) and MM LL.5513-8 figured by Brown (1981: fig. 43e). It could also be a partial ilium, resembling the proximal end of this element in C. svalbardensis (Knutsen et al. 2012c; Roberts et al. 2017). However, it does not appear broken in the narrowest portion, and if so, it is far shorter and more massive than the ilia in C. svalbardensis and Tatanectes laramiensis (USNM 536974, LLD personal observations; O’Keefe et al. 2009).
Two unidentified, massive partial elements probably belong to the coracoid or pubis, but they are too incomplete to be certain. The specimen also preserves four relatively large fragmentary bones and many smaller fragments that likely are parts of the girdles but are too incomplete to be assigned with certainty. As material is preserved from both the fore and hindlimbs, the partial girdle elements might belong to either the pectoral or pelvic girdle.
Limbs: The overall shapes and proportions of fore- and hindlimbs are relatively similar to each other in plesiosaurs, which is one of the reasons for the continued debate on swimming style. Four propodials are preserved in this specimen. The humeri and femora exhibit a significant difference in size and shape from each other, so they may become helpful to differentiate isolated forelimbs and hindlimbs in other specimens where this is ambiguous. Two propodials are proximodistally longer, more robust, anteroposteriorly wider, have more clearly defined distal facets, and one more distal facet than the other two. These two limb propodials also have a more posteriorly displaced prominence above the proximal head (epipophysis). In plesiosaurs, this prominence in the humerus (tuberosity) is typically displaced posteriorly relative to the epipophysis, whereas the prominence in the femur (trochanter) is situated directly above the epipophysis/capitulum (Andrews 1910; Brown 1981; Knutsen et al. 2012d; Roberts et al. 2017). The humerus and femur in this specimen can therefore be confidently differentiated on this basis.
Many long-necked plesiosaurs have longer forelimbs than hindlimbs (Krahl 2021), and within cryptoclidids, the humeri are more robust than femora (Arkhangelsky et al. 2020).
Humerus: The larger and more robust pair of propodials is interpreted as the humeri (Fig. 7A, B) based on the posteriorly positioned tuberosity, as in e.g., C. svalbardensis and Spitrasaurus wensaasi (Knutsen et al. 2012d; Roberts et al. 2017). The humerus of Tatanectes laramiensis appears less robust than the one in NHMD 189834, and has a smaller proximal head (O’Keefe and Street 2009; USNM 536976, LLD personal observations). The left humerus is almost complete, with a small part of the preaxial margin of the shaft reconstructed in plaster, and some damage to the distal facet for the radius. The tuberosity is posteriorly positioned (Fig. 7A). In proximal view, the tuberosity (dorsal) has a slightly smaller surface area than the capitulum (ventral), and the tuberosity and capitulum are partly separated by a constriction, the isthmus, which is more pronounced anteriorly than posteriorly. The right humerus is significantly more reconstructed in plaster but still the majority is intact. It is broken in the distal portion, missing parts of the anteriormost distal facet.
The Ellesmere Island specimen (NHMD 189834) is similar to C. svalbardensis (PMO 222.663) in having a mediolaterally wider humeral proximal head compared to the femora (Roberts et al. 2017). Because the tuberosity and the capitulum are not situated directly above each other, the proximal heads of the humeri are wider than the shafts in dorsal and ventral view. The Ellesmere Island specimen is also like C. svalbardensis (PMO 222.663) because the shafts of the humeri are more robust than those of the femora (Fig. 7). The shaft is at its minimum anteroposterior width directly distal to the proximal head, then increases in anteroposterior width very gradually, until a large increase for the expanded distal portion in the last third of the anteroposterior length. This contrasts with Spitrasaurus larseni, which widens anteroposteriorly throughout the shaft (Knutsen et al. 2012d). The distal portion widens more posteriorly, and only slightly towards the anterior direction. This is similar to C. svalbardensis, Tricleidus sp., and Tatanectes laramiensis (Brown 1981; O’Keefe and Street 2009; Knutsen et al. 2012c; Roberts et al. 2017), but differs from Spitrasaurus wensaasi and S. larseni which have evenly expanded distal ends anteriorly and posteriorly (Knutsen et al. 2012d).
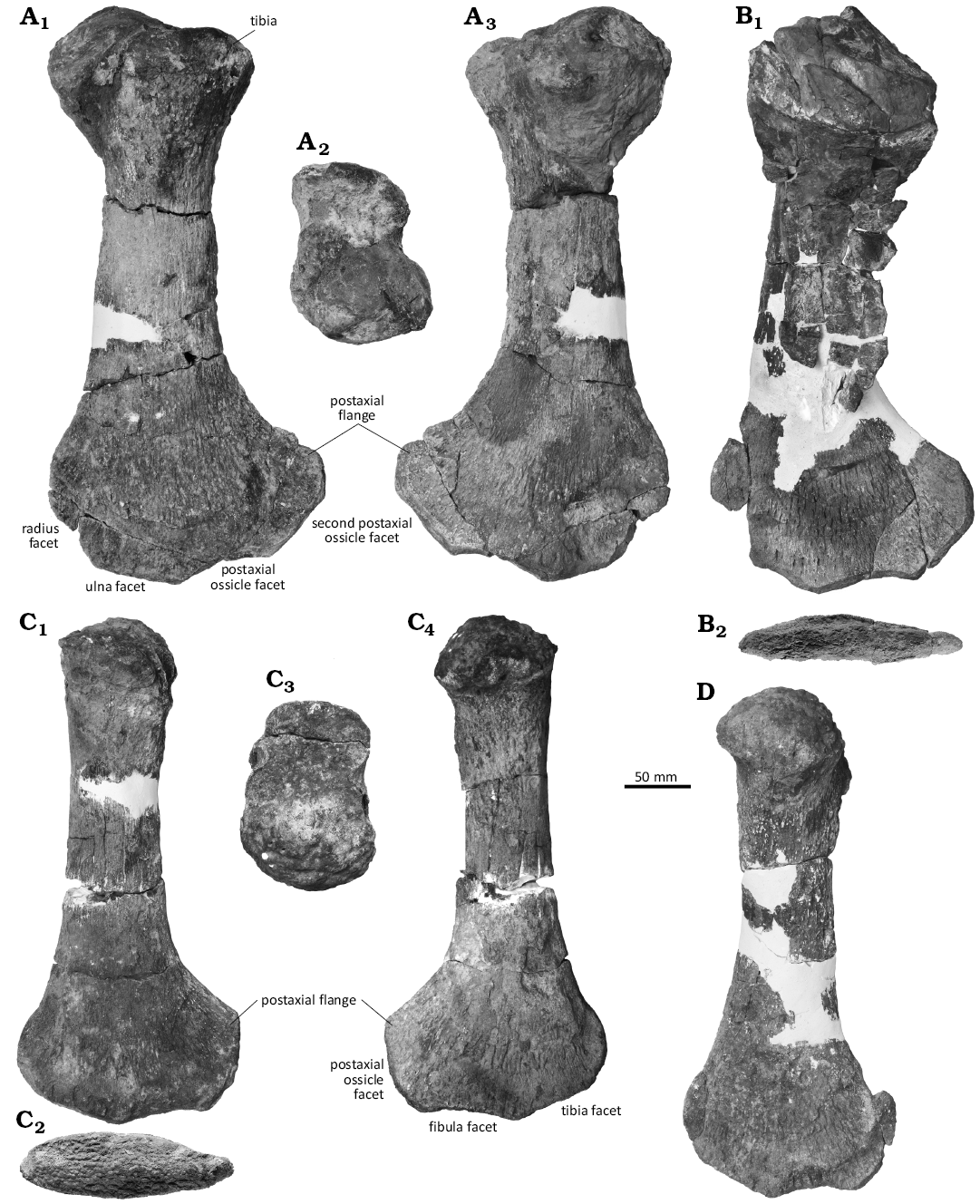
Fig. 7. Propodials of plesiosaur Colymbosaurus sp. NHMD 189834 from Ellesmere Island, Nunavut, Canada, upper Berriasian–lower Valanginian. A. Left humerus in dorsal (A1), proximal (A2), and ventral (A3) views. B. Right humerus in dorsal (B1) and distal (B2) views. C. Femur in dorsal or ventral (C1, C4), distal (C2), and proximal (C3) views. D. Femur in dorsal or ventral view.
The distal margin of the humerus has four distinct facets in dorsal view, as in Tatanectes laramiensis (O’Keefe and Street 2009) and “Plesiosaurus” manselii (Hulke, 1870). Colymbosaurus svalbardensis (Roberts et al. 2017) and Pantosaurus striatus (Wilhelm and O’Keefe 2010) have three facets, whereas in C. megadeirus the number of distal facets is unknown (Benson and Bowdler 2014; Fig. 8). The osteologically immature holotype of Spitrasaurus larseni has no facets, but it has three elements in the epipodial row (Knutsen et al. 2012d), which suggests that three humerus facets would have been present in osteologically mature individuals. Based on comparison to C. svalbardensis PMO 222.663 (Roberts et al. 2017), the anterior facet is interpreted as for the radius, the second facet is interpreted as the ulnar facet, and the third facet for the postaxial ossicle. The fourth facet probably articulated with a second postaxial ossicle, as seen in a complete articulated forelimb of a colymbosaurine (tentatively identified as Colymbosaurus sp.) from the Kimmeridge Clay of Wiltshire, UK (MJML K2334) (Fig. 8). The ulna facet is probably the widest one, but this is not fully certain as the anterior facet (radius) is not complete in either humerus. The ulna facet is also longest in Colymbosaurus svalbardensis (Roberts et al. 2017). In Pantosaurus striatus the radius facet is much longer and broader than the ulna facet (Wilhelm and O’Keefe 2010), in contrast to NHMD 189834, where the facets show the opposite relationship. The facet for the ulna is slightly concave, whereas the facets for the radius and the posterior accessory ossicle are flat.
On the distal surface of the right humerus (Fig. 7B2), a slight ridge bisects the epipodial facets. This ridge is less defined than in C. megadeirus in Benson and Bowdler (2014). In the left humerus, the presence of the ridge is uncertain.
Femora: Both femora are preserved (Fig. 7C, D). The femur is shorter and narrower than the humerus and has slenderer proximal ends in dorsal view due to the tuberosity being situated directly dorsal to the capitulum. In dorsal and ventral view, the shaft is narrower than the proximal head. In contrast, a proximal head and shaft of equal anteroposterior width is a unique trait of C. svalbardensis compared to all other taxa in the family Cryptoclididae (Roberts et al. 2017). The shaft widens anteroposteriorly, more prominently posteriorly, in the distalmost quarter of the element. This is similar to C. svalbardensis and C. megadeirus but differs from Spitrasaurus wensaasi (Knutsen et al. 2012d; Benson and Bowdler 2014; Roberts et al. 2017; Figs. 7, 8).
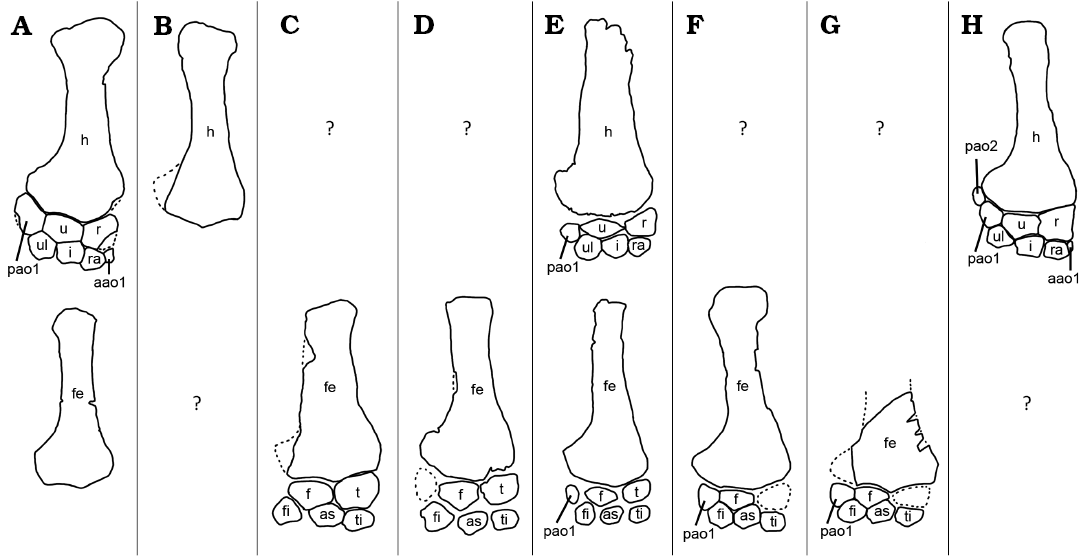
Fig. 8. Line drawings of reconstructed propodials, epipodials, and proximal mesopodial rows in Colymbosaurus specimens: Colymbosaurus sp. (A, H), Colymbosaurus megadeirus (B, C), Colymbosaurus svalbardensis (D–G), The top row shows the forelimbs, the bottom row shows the hindlimbs, question marks denote missing limbs. A. NHMD 189834, showing the partially reconstructed forelimb. Identification of the anterior accessory ossicle (aao1) in this specimen is tentative, so its position in the reconstruction is hypothetical. B. NHMUK PV OR 31787, the type specimen, which is just an isolated humerus C. CAMSM J.29596 etc. D. PMO 27745, the type specimen. E. PMO 222.663. F. PMO 216.838. G. PMO 218.377. H. MJML K2334, an undescribed specimen. Specimens are not to scale and some have been mirrored to ease comparison. Dotted lines show the possible outlines of missing elements or parts of elements. Abbreviations: aao1, anterior accessory ossicle; as, astragalus; f, fibula; fe, femur; fi, fibulare; h, humerus; i, intermedium; pao1, posterior accessory ossicle 1; pao2, posterior accessory ossicle 2; r, radius; ra, radiale; t, tibia; ti, tibiale; u, ulna; ul, ulnare.
The distal margin of the femur bears only three facets, which differentiates it from the four-faceted humerus. Colymbosaurus svalbardensis also has three facets on the femur (Knutsen et al. 2012c; Roberts et al. 2017; Fig. 7C1, C4, D). The femoral facets are also less pronounced than the facets on the humeri. The fibula facet, situated in the middle, is the anteroposteriorly longest of the three. The posteriormost facet for the postaxial element is the shortest. The fibula facet is concave, whereas the tibia facet is flat, and the posterior accessory ossicle facet is slightly convex.
On the distal surface of one femur (Fig. 7C2), a shallow ridge bisects the epipodial facets, whereas in the other femur, this feature is less clear.
Limb elements: In total 20 limb elements are preserved in addition to the propodials. Twelve of these are flattened elements that are interpreted as epipodials and mesopodials, and eight hourglass-shaped elements, interpreted as metapodials or phalanges.
Six elements can be confidently reconstructed along sharp and tightly fitting articulations (Fig 9A). These comprise an epipodial row and the proximal mesopodial row. The reconstructed epipodial row forms a combination of proximal facets that most closely matches the distal facets of the humeri in size and orientation (Fig. 9A). On this basis the reconstructed elements are interpreted as belonging to a forelimb. The epipodial row comprises at least three subequally sized elements: the radius, ulna, and a postaxial accessory ossicle that articulates with the ulna on the postaxial margin of the limb.
The radius is almost complete but missing part of the postaxial distal margin. It has a long straight facet for the humerus, a shorter facet for the ulna, and a postaxial facet that contacts two elements in the proximal mesopodial row (a short contact with the intermedium and a long contact with the radiale). The preserved part of the preaxial margin is convex. The overall shape and size of the radius are more similar to the proportions of the tibia of Colymbosaurus megadeirus (the ulna is not known in that species) than to those in the radius of C. svalbardensis. The facet on the radius for the ulna is relatively much shorter in C. svalbardensis (Roberts et al. 2017).
The ulna is of a pentagonal shape in dorsal view, with distinct facets for the humerus, radius, intermedium, ulnare, and postaxial accessory ossicle. This is unlike in C. svalbardensis (PMO 222.663), which has a diamond shape ulna with strongly reduced facets for the radius and postaxial accessory ossicle (Roberts et al. 2017). The preaxial facet on the ulna for the radius is slightly shorter than the postaxial facet for the postaxial accessory ossicle. The facets for the intermedium and ulnare are subequal. A similar condition in the hindlimb—“fibula … with facets for the fibulare and astragalus subequal in length”—was regarded as a synapomorphy of Colymbosaurus by Roberts et al (2017: 3). An additional isolated element can be identified as the second ulna because its shape and size match the equivalent element in the more complete forelimb (Fig. 9B). Both ulnae possess a distinct prominence in the middle of the distal margin between the facets for the intermedium and ulnare, similar to the fibula in the hindlimb of Colymbosaurus megadeirus (Benson and Bowdler 2014).

Fig. 9. Flipper elements of plesiosaur Colymbosaurus sp. NHMD 189834 from Ellesmere Island, Nunavut, Canada, upper Berriasian–lower Valanginian. A. Seven mesopodials that articulate together well, likely from a forelimb. B. The second ulna in dorsal or ventral view. C–F. The four additional mesopodials. G–J. Phalanges.
A further three limb-bones fit well along the distal margin of the epipodial row and articulate with each other, to form a proximal mesopodial row. These are therefore identified (preaxially to postaxially) as the radiale, intermedium, and ulnare. A smaller trapezoid-shaped element with a wedge-like transverse cross section tapering preaxially (or postaxially) also articulates well with this row (Fig. 9A), but fits equally well in the position of the second postaxial accessory ossicle, in articulation with the first postaxial accessory ossicle and the humerus (i.e., the fourth bone in the epipodial row). So, this bone cannot be placed with confidence. It could also potentially be from elsewhere in the forelimb, as many small subrectangular accessory ossicles are present along the preaxial margin of a complete articulated forelimb of a colymbosaurine (tentatively identified as Colymbosaurus sp.) from the Kimmeridge Clay of Wiltshire, UK (MJML K2334), one in the proximal mesopodial row, one in the distal mesopodial row, and one in the metapodial row (ASS personal observations). Alternatively, this ossicle could also be from the hindlimb.
Four additional mesopodials are preserved but are difficult to identify (Fig. 9C–F). None of these bones seems to articulate well with the reconstructed proximal mesopodial row of the forelimb, so they are probably not from the distal mesopodial row of the forelimb, and could be mesopodials from the hindlimb instead. The fore- and hindlimbs in plesiosaurs often show the same general features (Caldwell 1997), so comparison of these isolated bones with identified bones in the forelimb may help determine their position in the hindlimb. One of the mesopodials is a hexagonal wedge shape in dorsal view (Fig. 9E). It is similar, but not identical, in shape and size to the radiale. So, it could be from the equivalent position in the hindlimb, i.e., one of the tibiale. A second ossicle (Fig. 9D) is also hexagonal, but slightly elongate with two longer facets parallel to each other. It is similar, but not identical, in shape and size to the intermedium, so it could be the astragalus, the bone in the equivalent position in the hindlimb. The third element (Fig. 9F) is also hexagonal, with two longer facets subparallel to each other. It could also be an astragalus, but it is different to the one figured in Fig. 9D, so only one of them (or none of them) can be the astragalus. The fourth element (Fig. 9C) is the largest mesopodial in dorsal view. It is a disc-like hexagonal element with all facets subequal in length. It is also dorsoventrally shallower than the other mesopodials. It does not match any of the mesopodials in the preaxial row of the forelimb, but it is also larger in dorsal view, so it is unlikely to be a distal mesopodial. It is probably from the proximal mesopodial row of the hindlimb, may be the fibulare.
Eight phalanges are also preserved (four are shown in Fig. 9G–J), although the largest of these may be a metapodial. They are all expanded at their distal and proximal ends, and constricted in the middle, to form an hour-glass shape in dorsal or ventral view. Some are more proximodistally elongate than other, and one is almost as wide pre-postaxially as it is long proximodistally. Some of the elements also have an additional small facet on the proximal (?) surface, similar to the notches described in the tarsals of C. megadeirus (Benson and Bowdler 2014).
Phylogenetic analysis
Throughout all the phylogenetic analyses run with different settings, the placement of NHMD 189834 within the genus Colymbosaurus is consistent. The specimen either is recovered as a sister taxon of Colymbosaurus megadeirus or Colymbosaurus svalbardensis, or, as in the strict consensus tree, forms an unresolved polytomy with these two OTUs (Fig. 10).
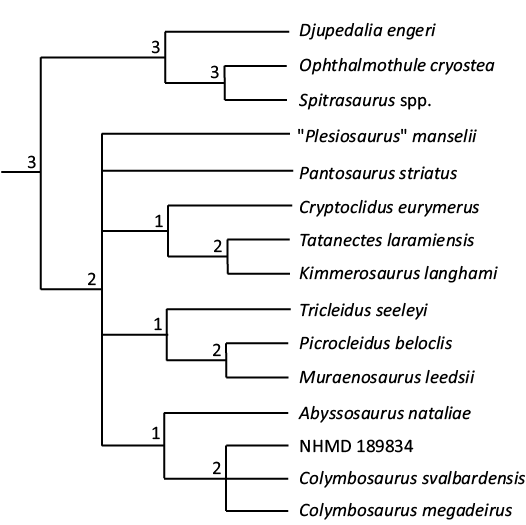
Fig. 10. Strict consensus tree with Bremer support values showing the phylogenetic placement of NHMD 189834 within Cryptoclididae.
Discussion
Taxonomy.—The morphology of the cervical vertebrae of NHMD 189834 is indicative of the plesiosauroid superfamily. The distal margin of the humeri, centrum proportions, the high neural arch and ovate canal, and zygopophyses borne high on the neural arch, are similar to cryptoclidids, and the specimen can be confidently placed within this well-supported family (Benson and Bowdler 2014; Roberts et al. 2020).
NHMD 189834 can be assigned to Colymbosaurus because it has (middle) caudal centra with a subrectangular articular face due to a flat ventral surface and widely spaced chevron facets, and propodials with a large posterodistal expansion (Roberts et al. 2017). It also shares with Colymbosaurus a postaxial ossicle facet of subequal size to the epipodial facets (Benson and Bowdler 2014); ulna conspicuously anteroposteriorly wider than the radius and proximodistally short; fibula symmetrically pentagonal in outline having equally long pre- and postaxial margins and with facets for the fibulare and astragalus subequal in length. The specimen also aligns with the emended diagnosis of Colymbosaurus (Roberts et al. 2017) with cervical vertebrae that are anteroposteriorly shorter than dorsoventrally tall. They also lack a longitudinal ridge on the lateral surface, a trait found in most elasmosaurids but only some cryptoclidids, e.g., Muraenosaurus.
The phylogenetic analysis (Fig. 10) also supports placement within Colymbosaurus, as specimen NHMD 189834 is consistently found to be most closely related to C. megadeirus and C. svalbardensis.
An anteroposteriorly directed ridge bisecting the epipodial facet has previously been used as a character for Colymbosaurus (Benson and Bowdler 2014). However, after re-examination of the type material, Roberts et al. (2017) called this character into question and removed it from the diagnosis of the genus, because its presence is inconsistent and might relate to taphonomy. The propodials of NHMD 189834 support the notion that this is a variable character, but the presence of a slight ridge on at least some of the propodials lends support to the assignment to Colymbosaurus.
The specific identification of the Ellesmere Island specimen is more problematic. There is little overlap of the material with specimens of C. megadeirus, so direct comparison with this species is limited. The anatomy of the humerus in NHMD 189834 and the holotype of C. megadeirus (Figs. 7, 8) is broadly similar, but the latter is incomplete, so the number of articular facets is unknown. A referred specimen of C. megadeirus (CAMSM J.29596 etc) is more completely known. The femur of NHMD 189834 is more gracile than the one in that specimen, but the epipodials and proximal mesopodials are generally similar, more closely similar than the equivalent bones in C. svalbardensis, even if only comparing the forelimb of NHMD 189834 with the hindlimb of CAMSM J.29596, etc. (Fig. 8).
NHMD 189834 shares one trait suggested as an autapomorphy with the species C. svalbardensis; a dorsoventrally taller neural canal on the mid-dorsal vertebrae, with a similar shape (Roberts et al. 2017). However, the state of this character is not known for certain in C. megadeirus. In addition, the proximal head of the femur in the present specimen is slightly wider than the femoral shaft, which might resemble more the situation in C. megadeirus than in C. svalbardensis (Roberts et al. 2017). The forelimb is only known in one referred specimen of C. svalbardensis (PMO 222.663). NHMD 189834 differs from that specimen in having a differently shaped ulna (Fig. 9A, E, see description), and a more gracile humerus with more defined distal facets. However, an exact comparison is confounded because the humerus in PMO 222.663 is slightly compressed and damaged (Roberts et al. 2017).
The number of sacral vertebrae and the shape of the posterior margin of the ischium, two characters used to differentiate C. megadeirus from C. svalbardensis, could not be evaluated as they are not preserved in the studied specimen. There are both similarities and differences between NHMD 189834 and both known species of Colymbosaurus, and it is possible that NHMD 189834 represents a distinct species of Colymbosaurus. However, the material has no clear autapomorphies and is too incomplete to justify the erection of a new taxon, so we refer it to Colymbosaurus in open nomenclature.
Palaeoecology and palaeoenvironment.—The Colymbosaurus sp. specimen NHMD 189834 from Ellesmere Island is the first plesiosaur described from the Lower Cretaceous of the Canadian Arctic and expands our knowledge of the biota occurring in the Sverdrup Basin during the Jurassic–Cretaceous transition interval. Its presence in this area is proof of a “normal” boreal marine environment hosting all the elements of a food chain supporting large predators. Plesiosaurs depended on fish and cephalopods as food (Foffa et al. 2018); however, while ammonites and belemnites both have a patchy fossil record, no fish fossils are known from the Deer Bay Formation. Curiously, the vertebrate-rich Slottsmøya Member in Spitsbergen also initially seemed to lack fish fossils; later, a number of isolated teleost remains were found (Koevoets et al. 2018), but still remain rare. For the Deer Bay Formation, the plesiosaur is the first, circumstantial evidence for the presence of fish. This observation may seem trivial. However, Jeletzky (1971) and Kemper (1975) originally regarded the Tithonian to Valanginian seas of the Sverdrup Basin as cold and hostile, home only to an impoverished, partly endemic biota, which had successfully adapted to these conditions. According to these authors, the fauna was further characterised by punctuated invasions during warmer episodes with higher sea levels. Recently, Schneider et al. (2020) argued that, although cold conditions were prevalent and sea levels were low during the latest Jurassic and earliest Cretaceous, the exceptionally low macro-biodiversity recorded from this interval was to a considerable extent a function of a poor fossil record. Macrofauna, predominantly ammonites, bivalves and belemnites, is restricted to a few horizons in the Deer Bay Formation, which are characterised by the occurrence of siderite concretions (Schneider et al. 2020). This suggests that macrofossil-bearing horizons represent favourable taphonomical snapshots rather than punctuated invasions.
Another striking feature of this interval is the near absence of shallow shelf biota. Coastal and marginal marine communities, which are typically characterised by higher abundances and diversities than those from deeper waters, are clearly underrepresented in the Tithonian to Valanginian of the Sverdrup Basin. We assume that this is predominantly a matter of the preservation and exposure of the respective strata and fossils. The Deer Bay Formation is dominated by mudstones, and thus predominantly represents more distal environments, where shallow marine taxa are absent. The corresponding Awingak Formation is characterised by sandstones generally relating to more proximal settings (Fig. 2), but its fossil record is sparse (Balkwill 1983; Poulton 1994). However, some of the fossils collected on the Queen Elizabeth Islands, including the plesiosaur described herein, simply remained unpublished. This includes dense bivalve pavements of an assemblage of around 15 taxa from the Awingak Formation of the study area (which is rather diverse by boreal measures), but also most of the benthic invertebrates collected by Troelsen (1952) (SS personal observations). With few exceptions (Frebold 1964; Jeletzky and Poulton 1987; Schneider et al. 2020), only those fossils relevant for biostratigraphy, i.e., ammonites and bivalves of the genus Buchia, were described and figured from the Sverdrup Basin (Frebold 1961; Jeletzky 1964, 1965, 1966, 1970, 1973, 1979, 1984, 1986; Kemper 1975; Kemper and Jeletzky 1979; Jeletzky and Kemper 1988; Galloway et al. 2020). Lists of additional Late Jurassic to Valanginian macrofossils are scattered over geological mapping reports or studies of regional geology (Fortier et al. 1963; Fricker 1963; Tozer and Thorsteinsson 1964; Stott 1969; Balkwill et al. 1977; Balkwill 1983; Poulton 1994). As a result, our knowledge of the Tithonian to Valanginian environments of the Sverdrup Basin remains remarkably incomplete.
Palaeobiogeography.—The new Colymbosaurus specimen from Ellesmere Island significantly expands the geographical range of the genus, which was formerly known from Spitsbergen, southern England and western Russia (Knutsen et al. 2012c, d; Benson and Bowdler 2014; Roberts et al. 2017; Arkhangelsky et al. 2020). This underpins that, unlike most other plesiosaur genera (Bardet et al. 2014), Colymbosaurus was widespread and not endemic to a single basin. Its rather distant occurrences are evidence of faunal exchange between the Boreal Arctic (Arctic Canada, Siberia, Svalbard) and Boreal Atlantic sub-realms (European parts of Russia, UK) during the Late Jurassic (Arkhangelsky et al. 2020). North-south connectivity was facilitated by the NE Atlantic seaway between Norway and Greenland in the west, and in the east by the broad, shallow Timan-Pechora-Moscow-Precaspian Seaway in present day western Russia (Mutterlose et al. 2003). For the Svalbard region, the large degree of endemicity of invertebrates and a partly closed seaway between the Boreal and Tethyan realms in the Late Jurassic raised the question whether marine reptiles from Spitsbergen were endemic (Tyborowski and Błażejowski 2019). However, this was subsequently shown not to be the case for ichthyosaurs (Delsett et al. 2019; Zverkov and Prilepskaya 2019). Moreover, close faunal relationships to Spitsbergen and Russia have been proven for an ichthyosaur from the Ringnes Formation of Melville Island (Maxwell 2010; Delsett et al. 2019; Zverkov and Prilepskaya 2019).
For the Deer Bay and Awingak formations, faunal relationships outside the Sverdrup Basin have previously been documented for ammonites, belemnites and bivalves (Jeletzky 1971, 1973, 1984; Mutterlose et al. 2020; Schneider et al. 2020) and are now also established for plesiosaurs. In turn, only very few endemic invertebrate taxa are known from these units (Jeletzky 1966; Jeletzky and Poulton 1987). However, diversities of these taxa are still significantly higher in coeval strata in other parts of the Arctic (Fürsich 1982, 1984; Zakharov et al. 2003; Rogov and Zakharov 2009; Mutterlose et al. 2020). This is also the case for vertebrates, as evidenced by the Slottsmøya Member of Spitsbergen, where at least five plesiosauroid and two pliosauroid species were present (Knutsen et al. 2012a–d; Delsett et al. 2016; Roberts et al. 2017, 2020). Whether this is a function of intermittently restricted connectivity or rather the fossil record and its documentation, as discussed above (see also Maxwell 2010), remains unclear. Palaeoclimate has also been discussed as a controlling factor in marine reptile distribution (Foffa et al. 2018), and the higher latitude of the Sverdrup Basin relative to comparable sites may have played a role.
It is obvious, though, that different taxa had different dispersal strategies and capabilities. Plesiosaurs were adapted to moderately fast but efficient long-distance swimming, which, together with endothermy, certainly helped with propagation (O’Keefe 2001; Bernard et al. 2010; Fleischle et al. 2018). Furthermore, osteohistological analysis of two C. svalbardensis specimens from Spitsbergen indicated that these plesiosaurs were active animals, characterized by rapid growth and high metabolic rates (Liebe and Hurum 2012). Similarly, ammonites and belemnites were more or less capable swimmers, and species recorded from the Deer Bay Formation provide evidence for connections to East Greenland, Svalbard and different parts of Siberia (Schneider et al. 2020). On the other hand, bivalves, including the widespread Buchia, were reliant on ocean currents for dispersal during their presumably planktonic larval stage. Nevertheless, most Buchia species, including those from the Deer Bay Formation, are widely distributed in the Boreal Realm (Jeletzky 1973, 1984; Zakharov 1981; Zakharov and Rogov 2020; Schneider et al. 2020). In contrast, Boreal dinoflagellate cyst assemblages from the Jurassic/Cretaceous boundary interval are markedly provincial in aspect (see discussion in Ingrams et al. 2021). As a result, first occurrence dates of shared dinocyst taxa are not the same in different basins, and the Sverdrup Basin zonal scheme of Davies (1983) does not correlate at the Pan-Boreal scale (e.g., Harding et al. 2011; Pestchevitskaya et al. 2011; Ingrams et al. 2021).
Crossing the Jurassic/Cretaceous boundary.—The Colymbosaurus sp. specimen from Ellesmere Island is significant, because it adds to a sparse record of Berriasian–Valanginian plesiosaur specimens and taxa known worldwide (Benson et al. 2013). These include Brancasaurus brancai and a Plesiosauria indet. specimen from the Berriasian of Germany (Sachs et al. 2016, 2018), Hastanectes valdensis and several incomplete specimens from the Valanginian of the UK (Kear and Barrett 2011; Benson et al. 2013), late Valanginian Leptocleidus capensis from South Africa (Cruickshank 1997), and indeterminate plesiosaur specimens from Peru (Meza-Vélez and O’Gorman 2020) and Argentina (Lazo et al. 2018). Ophthalmothule cryostea from the uppermost Tithonian or lowermost Berriasian of Spitsbergen and Brancasaurus brancai are unusual in having major parts of their skeletons preserved (Roberts et al. 2020). Consequently, NHMD 189834 from Ellesmere Island is the most complete plesiosaur from the Berriasian–Valanginian time interval in North America, and among the most complete specimens from this interval globally.
Cryptoclidids were long necked plesiosauroids with small skulls, known from the Middle Jurassic to the earliest Cretaceous (Benson and Druckenmiller 2014). Most taxa and specimens are known from the Northern Hemisphere (Fig. 11). These include Colymbosaurus megadeirus from the Kimmeridge Clay Formation and Muraenosaurus leedsii from the Oxford Clay Formation in the UK; Colymbosaurus svalbardensis, Spitrasaurus larseni, Spitrasaurus wensaasi, Djupedalia engeri, and Ophthalmothule cryostea from the Slottsmøya Member Lagerstätte of Spitsbergen, Norway; Pantosaurus striatus and Tatanectes laramiensis from the Oxfordian Sundance Formation in Wyoming, USA; Abyssosaurus nataliae, Colymbosaurus sp., and Colymbosaurinae indet. from the Tithonian (Volgian) of western Russia and a Cryptoclididae? indet. specimen from Poland (O’Keefe and Street 2009; O’Keefe et al. 2011; Benson and Bowdler 2014; Berezin 2018; Arkhangelsky et al. 2020; Roberts et al. 2020; Madzia et al. 2021). Furthermore, cryptoclidids have been reported from the Oxfordian Cerra Campamento Formation of Chile (Otero et al. 2020). Potential additional Southern Hemisphere records come from the Oxfordian Jagua Formation of Cuba, as well as from Argentina and India (Bardet et al. 1991; Gasparini and Spalletti 1993; Gasparini et al. 2002).
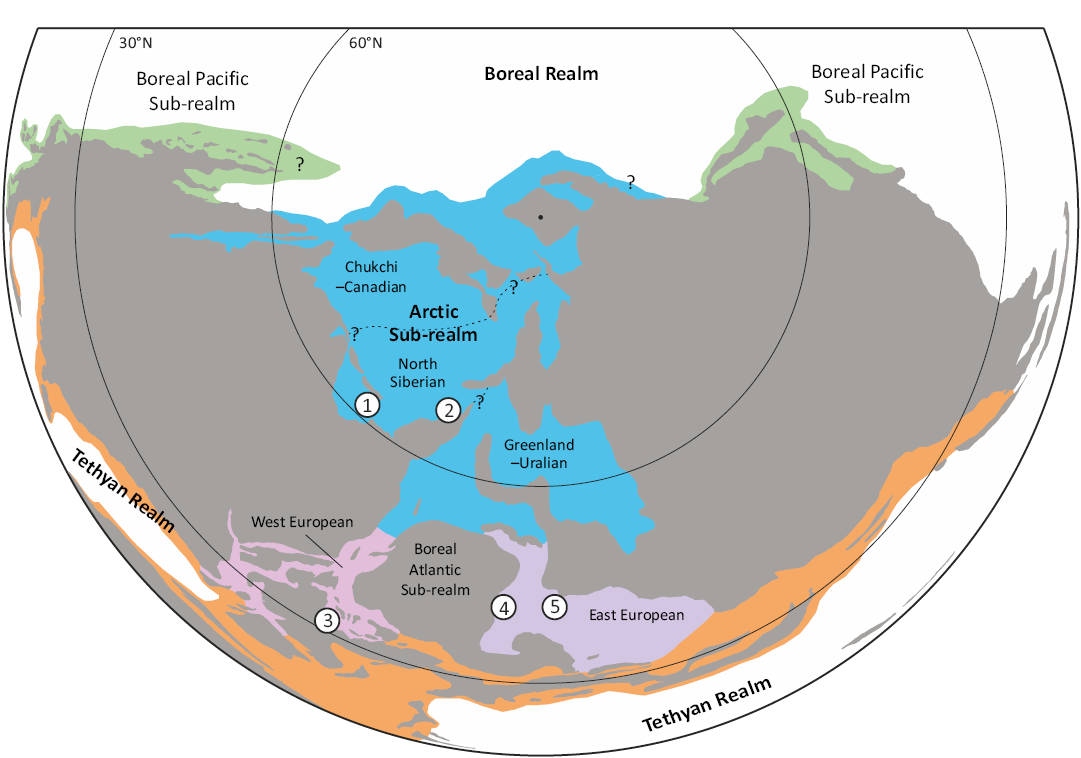
Fig. 11. Map with cryptoclidid occurrences in the Northern hemisphere. 1, Ellesmere Island; 2, Spitsbergen; 3, Great Britain; 4, 5, Russia (modified from Ingrams et al. 2021). Question marks indicate approximate boundaries of sub-realms and bio-provinces.
Cryptoclididae includes three or four lineages, of which only Colymbosaurinae, to which the genus Colymbosaurus belongs, has been formally named (Benson and Bowdler 2014; Roberts et al. 2020; Sachs et al. 2024). Based on the records of Abyssosaurus nataliae from the Lower Cretaceous of western Russia and several colymbosaurinae specimens from the uppermost Jurassic and possibly lowermost Cretaceous of Spitsbergen, Roberts et al. (2017) suggested that the group survived through the Jurassic–Cretaceous transition in the Boreal Realm. The Early Cretaceous specimen described here from Ellesmere Island verifies this hypothesis and expands the geographical range for the group.
Conclusions
This work describes a plesiosaur specimen from the Cretaceous on Ellesmere Island, seventy years after it was collected. The specimen preserves all four propodials, which is rare. Also preserved are several associated limb elements, vertebrae from several regions of the body, and some incomplete girdle elements. The specimen is assigned to Colymbosaurus sp., thus expanding the geographical and temporal range of this genus, and showing that the Sverdrup Basin held a fully developed marine ecosystem, including large predators, with connections to other ocean basins, at least for actively swimming animals.
Acknowledgments
The authors wish to thank Arden R. Bashforth and Bent Lindow (both Natural History Museum in Copenhagen, Denmark) for great support and collection access. Laura Cotton is warmly thanked for assistance and for taking some of the photographs together with Sophia K.G. Christensen (both Natural History Museum in Copenhagen). In addition, Louise I. Ahl (Natural History Museum in Copenhagen) is thanked for good help. For collection access at Smithsonian’s NMNH thanks to Hans-Dieter Sues. Aubrey J. Roberts (Natural History Museum, Oslo, Norway) is thanked for sharing her thoughts on cryptoclidids. Reviewers Sven Sachs (Naturkunde-Museum Bielefeld, Bielefeld, Germany) and Daniel Madzia (Institute of Paleobiology PAS, Warsaw, Poland) are thanked for comments that improved the manuscript. LLD wishes to acknowledge funding from the Palaeontological Association through a Stan Wood Award (PA-SW202001), Synthesys+ for a Transnational access grant, and a US-Norway Fulbright research grant.
References
Andrews, C.W. 1910. A Descriptive Catalogue of the Marine Reptiles of the Oxford Clay. Based on the Leeds Collection in the British Museum (Natural History), London. Part I. 205 pp. Trustees of the British Museum, London.
Araújo, R. and Smith, A.S. 2023. Recognising and quantifying the evolution of skeletal paedomorphosis in Plesiosauria. Fossil Record 26: 85–101. Crossref
Arkhangelsky, M.S., Zverkov, N.G., Rogov, M.A., Stenshin, I.M., and Baykina, E.M. 2020. Colymbosaurines from the Upper Jurassic of European Russia and their implication for palaeobiogeography of marine reptiles. Palaeobiodiversity and Palaeoenvironments 100: 197–218. Crossref
Balkwill, H.R. 1978. Evolution of Sverdrup Basin, Arctic Canada. The American Association of Petroleum Geologists Bulletin 62: 1004–1028. Crossref
Balkwill, H.R. 1983. Geology of Amund Ringnes, Cornwall and Haig-Thomas islands, District of Franklin. Geological Survey of Canada, Memoir 390: 1–76. Crossref
Balkwill, H.R., Wilson, D.G., and Wall, J.H. 1977. Ringnes Formation (Upper Jurassic), Sverdrup Basin, Canadian Arctic Archipelago. Bulletin of Canadian Petroleum Geology 25: 1115–1144.
Bardet, N., Falconnet, J., Fischer, V., Houssaye, A., Jouve, S., Suberbiola, X.P., Perez-Garcia, A., Rage, J.C., and Vincent, P. 2014. Mesozoic marine reptile palaeobiogeography in response to drifting plates. Gondwana Research 26: 869–887. Crossref
Bardet, N., Mazin, J.M., Cariou, E., Enay, R., and Krishna, J. 1991. Les Plesiosauria du Jurassique supérieur de la province de Kachchh (Inde). Comptes rendus de l’Academie des Sciences de Paris 313: 1343–1347.
Benson, R.B.J. and Bowdler, T. 2014. Anatomy of Colymbosaurus megadeirus (Reptilia, Plesiosauria) from the Kimmeridge Clay Formation of the U.K., and high diversity among Late Jurassic plesiosauroids. Journal of Vertebrate Paleontology 34: 1053–1071. Crossref
Benson, R.B.J. and Druckenmiller, P.S. 2014. Faunal turnover of marine tetrapods during the Jurassic–Cretaceous transition. Biological Reviews 89: 1–23. Crossref
Benson, R.B.J., Evans, M., and Druckenmiller, P.S. 2012. High diversity, low disparity and small body size in plesiosaurs (Reptilia, Sauropterygia) from the Triassic–Jurassic boundary. PLoS ONE 7 (3): e31838. Crossref
Benson, R.B.J., Ketchum, H.F., Naish, D., and Turner, L.E. 2013. A new leptocleidid (Sauropterygia, Plesiosauria) from the Vectis Formation (Early Barremian–early Aptian; Early Cretaceous) of the Isle of Wight and the evolution of Leptocleididae, a controversial clade. Journal of Systematic Palaeontology 11: 233–250. Crossref
Berezin, A.Y. 2011. A new plesiosaur of the family Aristonectidae from the early Cretaceous of the center of the Russian platform. Paleontological Journal 45: 648–660. Crossref
Berezin, A.Y. 2018. Craniology of the plesiosaur Abyssosaurus nataliae Berezin (Sauropterygia, Plesiosauria) from the Lower Cretaceous of the Central Russian Platform. Paleontological Journal 52: 328–341. Crossref
Bernard, A., Lécuyer, C., Vincent, P., Amiot, R., Bardet, N., Buffetaut, E., Cuny, G., Fourel, F., Martineau, F., Mazin, J.-M., and Prieur, A. 2010. Regulation of body temperature by some Mesozoic marine reptiles. Science 328: 1379–1382. Crossref
Brown, D.S. 1981. The English Upper Jurassic Plesiosauroidea (Reptilia) and a review of the phylogeny and classification of the Plesiosauria. Bulletin of the British Museum (Natural History) 35: 253–347.
Caldwell, M.W. 1997. Limb osteology and ossification patterns in Cryptoclidus (Reptilia: Plesiosauroidea) with a review of sauropterygian limbs. Journal of Vertebrate Paleontology 17: 295–307. Crossref
Cruickshank, A.R.I. 1997. A Lower Cretaceous pliosaurid from South Africa. Annals of the South African Museum 105: 207–226.
Davies, E.H. 1983. The dinoflagellate Oppel-zonation of the Jurassic– Lower Cretaceous sequence in the Sverdrup basin, Arctic Canada. Geological Survey of Canada Bulletin 359: 1–59. Crossref
de Blainville, M.H.D. 1835. Description de quelques espèces de reptiles de la Californie, précédée de l’analyse d’un systéme général d’erpétologie et d’amphibiologie. Nouvelles Annales du Muséum d’Histoire Naturelle 4: 233–296.
Delsett, L.L., Novis, L.K., Roberts, A.J., Koevoets, M.J., Hammer, Ø., Druckenmiller, P.S., and Hurum, J.H. 2016. The Slottsmøya marine reptile Lagerstätte: depositional environments, taphonomy and diagenesis. Geological Society Special Publications: Mesozoic Biotas of Scandinavia and its Arctic Territories 434: 165–188. Crossref
Delsett, L.L., Roberts, A.J., Druckenmiller, P.S., and Hurum, J.H. 2019. Osteology and phylogeny of Late Jurassic ichthyosaurs from the Slottsmøya Member Lagerstätte (Spitsbergen, Norway). Acta Palaeontologica Polonica 64: 717–743. Crossref
Döring, H. 1965. Die sporenpaläontologische Gliederung des Wealden in Westmecklenburg (Struktur Werle). Geologie, Beihefte 47: 1–118.
Embry, A. 1986. Stratigraphic subdivision of the Awingak Formation (Upper Jurassic) and revision of the Hiccles Cove Formation (Middle Jurassic), Sverdrup Basin, Arctic Islands. Geological Survey of Canada 86-1B: 341–349. Crossref
Embry, A. and Beauchamp, B. 2019. Sverdrup Basin. In: A.D. Miall (ed.), The Sedimentary Basins of the United States and Canada (Second Edition), 559–592. Elsevier, Amsterdam. Crossref
Embry, A., Beauchamp, B., Dewing, K., and Dixon, J. 2018. Episodic tectonics in the Phanerozoic succession of the Canadian High Arctic and the “10-million-year flood”. Geological Society of America Special Paper 541: 1–18. Crossref
Fleischle, C.V., Wintrich, T., and Sander, P.M. 2018. Quantitative histological models suggest endothermy in plesiosaurs. PeerJ 6: e4955. Crossref
Foffa, D., Young, M.T., Stubbs, T.L., Dexter, K.G., and Brusatte, S.L. 2018. The long-term ecology and evolution of marine reptiles in a Jurassic seaway. Nature Ecology & Evolution, 2, 1548–1555. Crossref
Fortier, Y.O., Blackadar, R.G., Glenister, B.F., Greiner, H.R., McLaren, D.J., McMillan, N.J., Norris, A.W., Roots, E.F., Souther, J.G., Thorsteinsson, R., and Tozer, E.T. 1963. Geology of the north-central part of the Arctic Archipelago, Northwest Territories (Operation Franklin). Geological Survey of Canada Memoir 320: 1–704. Crossref
Frebold, J. 1961. The Jurassic faunas of the Canadian Arctic. Middle and Upper Jurassic ammonites. Geological Survey of Canada Bulletin 74: 1–43. Crossref
Frebold, J. 1964. Illustrations of Canadian fossils, Jurassic of western and Arctic Canada. Geological Survey of Canada Paper 63: 1–104. Crossref
Fricker, P.E. 1963. Jacobsen-McGill Arctic Research Expedition 1959–1962, Geology, No. 1. Geology of the Expedition Area, Western Central Axel Heiberg Island, Canadian Arctic Archipelago. 156 pp. McGill University, Montreal.
Fürsich, F.T. 1982. Upper Jurassic bivalves from Milne Land, East Greenland. Bulletin, Grønlands Geologiske Undersøgelse 144: 1–126. Crossref
Fürsich, F.T. 1984. Benthic macroinvertebrate associations from the Boreal Upper Jurassic of Milne Land, central East Greenland. Bulletin, Grønlands Geologiske Undersøgelse 149: 1–72. Crossref
Galloway, J.M., Vickers, M.L., Price, G.D., Poulton, T.P., Grasby, S.E., Hadlari, T., Beauchamp, B., and Sulphur, K. 2020. Finding the VOICE: organic carbon isotope chemostratigraphy of Late Jurassic–Early Cretaceous Arctic Canada. Geological Magazine 157: 1643–1657. Crossref
Gasparini, Z., Bardet, N., and Iturralde-Vinent, M. 2002. A new cryptoclidid plesiosaur from the Oxfordian (Late Jurassic) of Cuba. Geobios 35 (2): 201–211. Crossref
Gasparini, Z. and Spalletti, L. 1993. First Callovian plesiosaurs from the Neuquen Basin, Argentina. Ameghiniana 30: 245–254.
Goloboff, P.A. and Morales, M.E. 2023. TNT, version 1.6, with a graphical interface for MacOS and Linux, including new routines in parallel. Cladistics 39: 144–153. Crossref
Hadlari, T., Midwinter, D., Galloway, J.M., Dewing, K., and Durbano, A.M. 2016. Mesozoic rift to post-rift tectonostratigraphy of the Sverdrup Basin, Canadian Arctic. Marine and Petroleum Geology 76: 148–158. Crossref
Hampe, O. 2013. The forgotten remains of a leptocleidid plesiosaur (Sauropterygia: Plesiosauroidea) from the Early Cretaceous of Gronau (Münsterland, Westphalia, Germany). Paläontologische Zeitschrift 87: 473–491. Crossref
Harding, I.C., Smith, G.A., Riding, J.B., and Wimbledon, W.A.P. 2011. Inter-regional correlation of Jurassic/Cretaceous boundary strata based on the Tithonian–Valanginian dinoflagellate cyst biostratigraphy of the Volga Basin, western Russia. Review of Palaeobotany and Palynology 167: 82–116. Crossref
Harrison, J.C., St-Onge, M.R., Petrov, O.V., Strelnikov, S.I., Lopatin, B.G., Wilson, F.H., Tella, S., Paul, D., Lynds, T., Shokalsky, S.P., Hults, C.K., Bergman, S., Jepsen, H.F., and Solli, A. 2011. Geological Map of the Arctic. Map 2159A. Geological Survey of Canada, Ottawa. Crossref
Hulke, J.W. 1870. Notes on some plesiosaurian remains obtained by J.C. Mansel, Esq., F.G.S., in Kimmeridge Bay, Dorset. Quarterly Journal of the Geological Society 26 (1–2): 611–622. Crossref
Ingrams, S., Jolley, D.W., and Schneider, S. 2021. High latitude stratigraphical palynology of the Jurassic–Cretaceous boundary interval, Sverdrup Basin, Arctic Canada. Cretaceous Research 126: 104922. Crossref
Ingrams, S., Jolley, D.W., and Schneider, S. 2022. Sequence stratigraphy and microplankton palaeoenvironmental dynamics across the Jurassic–Cretaceous transition in the Canadian Arctic. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 113: 159–174. Crossref
Jeletzky, J.A. 1964. Illustrations of Canadian fossils. Lower Cretaceous marine index fossils of the sedimentary basins of western and Arctic Canada. Geological Survey of Canada Paper 64: 1–101. Crossref
Jeletzky, J.A. 1965. Thorsteinssonoceras, a new craspeditid ammonite from the Valanginian of Ellesmere Island, Arctic Archipelago. Geological Survey of Canada Bulletin 120: 1–16. Crossref
Jeletzky, J.A. 1966. Upper Volgian (latest Jurassic) ammonites and buchias of Arctic Canada. Geological Survey of Canada 128: 1–51. Crossref
Jeletzky, J.A. 1970. Marine Cretaceous biotic provinces and paleogeography of western and Arctic Canada: Illustrated by a detailed study of ammonites. Geological Survey of Canada Paper 70: 1–92. Crossref
Jeletzky, J.A. 1971. Biochronology of Jurassic–Cretaceous transition beds in Canada. Geological Survey of Canada Paper 71: 1–8. Crossref
Jeletzky, J.A. 1973. Biochronology of the marine boreal latest Jurassic, Berriasian and Valanginian in Canada. In: R. Casey and P.F. Rawson (eds.), The Boreal Lower Cretaceous. Geological Journal Special Issue 5: 41–80.
Jeletzky, J.A. 1979. Eurasian craspeditid genera Temnoptychites and Tollia in the Lower Valanginian of Sverdrup Basin, District of Franklin, with comments on taxonomy and nomenclature of Craspeditidae. Geological Survey of Canada 299: 1–89. Crossref
Jeletzky, J.A. 1984. Jurassic–Cretaceous boundary beds of western and Arctic Canada and the problem of the Tithonian–Berrisian stages in the Boreal Realm. In: G.E.G. Westermann (ed.), Jurassic–Cretaceous Biochronology and Biogeography of North America. Geological Association of Canada, Special Paper 27: 175–255.
Jeletzky, J.A. 1986. Pseudoeuryptychites: a new polyptychitinid ammonite from the Lower Valanginian of the Canadian and Eurasian Arctic. Geological Survey of Canada Paper 86: 351–361. Crossref
Jeletzky, J.A. and Kemper, E. 1988. Comparative paleontology and stratigraphy of Valanginian Polyptychitinae and Simbirskitinae in Sverdrup Basin (Arctic Canada) and Lower Saxony Basin (northwest Germany). Geological Survey of Canada Bulletin 377: 1–355. Crossref
Jeletzky, J.A. and Poulton, T.P. 1987. A new genus and subgenus and two new species of latest Jurassic oxytomid bivalves from Arctic Canada. Canadian Journal of Earth Sciences 24: 711–722. Crossref
Kear, B.P. and Barrett, P.M. 2011. Reassessment of the Lower Cretaceous (Barremian) pliosauroid Leptocleidus superstes Andrews, 1922 and other plesiosaur remains from the nonmarine Wealden succession of southern England. Zoological Journal of the Linnean Society 161: 663–691. Crossref
Kemper, E. 1975. Upper Deer Bay Formation (Berriasian–Valanginian) of Sverdrup Basin and biostratigraphy of the Arctic Valanginian. Geological Survey of Canada 75-1B: 45–54. Crossref
Kemper, E. and Jeletzky, J.A. 1979. New stratigraphically and phylogenetically important olcostephanid (Ammonitida) taxa from the uppermost Lower and Upper Valanginian of Sverdrup Basin. Geological Survey of Canada Paper 79: 1–25. Crossref
Knutsen, E.M., Druckenmiller, P.S., and Hurum, J.H. 2012a. A new plesiosauroid (Reptilia: Sauropterygia) from the Agardhfjellet Formation (Middle Volgian) of central Spitsbergen, Norway. Norwegian Journal of Geology 92 (2–3): 213–234.
Knutsen, E.M., Druckenmiller, P.S., and Hurum, J.H. 2012b. A new species of Pliosaurus (Sauropterygia: Plesiosauria) from the Middle Volgian of central Spitsbergen, Norway. Norwegian Journal of Geology 92 (2–3): 235–258.
Knutsen, E.M., Druckenmiller, P.S., and Hurum, J.H. 2012c. Redescription and taxonomic clarification of “Tricleidus” svalbardensis based on new material from the Agardhfjellet Formation (Middle Volgian). Norwegian Journal of Geology 92 (2–3): 175–186.
Knutsen, E.M., Druckenmiller, P.S., and Hurum, J.H. 2012d. Two new species of long-necked plesiosaurians (Reptilia: Sauropterygia) from the Upper Jurassic (Middle Volgian) Agardhfjellet Formation of central Spitsbergen. Norwegian Journal of Geology 92 (2–3): 187–212.
Koevoets, M.J., Hurum, J.H., and Hammer, Ø. 2018. New Late Jurassic teleost remains from the Agardhfjellet Formation, Spitsbergen, Svalbard. Norwegian Journal of Geology 98 (2): 1–12. Crossref
Krahl, A. 2021. The locomotory apparatus and paraxial swimming in fossil and living marine reptiles: comparing Nothosauroidea, Plesiosauria, and Chelonioidea. PalZ 95: 483–501. Crossref
Krymgolts, G.Y., Petrova, G.T., and Pchelintsev, V.F. 1953. Stratigraphy and fauna of the marine Mesozoic deposits of Northern Siberia [in Russian]. Trudy naučno-issledovatel’skogo instituta geologii Arktiki Glavsevmorputi pri Sovete Ministrov SSSR 45, 1–104.
Lahusen, J. 1888. Über die russischen Aucellen. Mémoires du Comité Geologique 8: 1–46.
Lazo, D.G., Talevi, M., Cataldo, C.S., Aguirre-Urreta, B., and Fernández, M.S. 2018. Description of ichthyosaur remains from the Lower Cretaceous Agrio Formation (Neuquén Basin, west-central Argentina) and their paleobiological implications. Cretaceous Research 89: 8–21. Crossref
Liebe, L. and Hurum, J.H. 2012. Gross internal structure and microstructure of plesiosaur limb bones from the Late Jurassic, central Spitsbergen. Norwegian Journal of Geology 92 (2–3): 285–309.
Madzia, D. and Cau, A. 2020. Estimating the evolutionary rates in mosasauroids and plesiosaurs: discussion of niche occupation in Late Cretaceous seas. PeerJ 8: e8941. Crossref
Madzia, D., Szczygielski, T., and Wolniewicz, A. 2021. The giant pliosaurid that wasn’t—revising the marine reptiles from the Kimmeridgian, Upper Jurassic, of Krzyżanowice, Poland. Acta Palaeontologica Polonica 66 (1): 99–129. Crossref
Maxwell, E.E. 2010. Generic reassignment of an ichthyosaur from the Queen Elizabeth Islands, Northwest Territories, Canada. Journal of Vertebrate Paleontology 30: 403–415. Crossref
Meza-Vélez, I. and O’Gorman, J.P. 2020. First plesiosaurian record (Diapsida; Sauropterygia) from the La Herradura formation, (Valanginian–Hauterivian), Morro Solar, Peru. Cretaceous Research 106: 104247. Crossref
Mutterlose, J., Alsen, P., Iba, Y., and Schneider, S. 2020. Palaeobiogeography and palaeoecology of Early Cretaceous belemnites from the northern high latitudes. Proceedings of the Geologists’ Association 131: 278–286. Crossref
Mutterlose, J., Brumsack, H., Flögel, S., Hay, W., Klein, C., Langrock, U., Lipinski, M., Ricken, W., Söding, E., Stein, R., and Swientek, O. 2003. The Greenland-Norwegian Seaway: A key for understanding Late Jurassic to Early Cretaceous paleoenvironments. Paleoceanography 18: 1–26. Crossref
Norris, G. 1969. Miospores from the Purbeck Beds and marine Upper Jurassic of southern England. Palaeontology 12: 574–620.
O’Keefe, F.R. 2001. Ecomorphology of plesiosaur flipper geometry. Journal of Evolutionary Biology 14 (6): 987–991. Crossref
O’Keefe, F.R. and Street, H.P. 2009. Osteology of the cryptocleidoid plesiosaur Tatenectes laramiensis, with comments on the taxonomic status of the Cimoliasauridae. Journal of Vertebrate Paleontology 29: 48–57. Crossref
O’Keefe, F.R., Street, H.P., Cavigelli, J.P., Socha, J.J., and O’Keefe, R.D. 2009. A plesiosaur containing an ichthyosaur embryo as stomach contents from the Sundance Formation of the Bighorn Basin, Wyoming. Journal of Vertebrate Paleontology 29: 1306–1310. Crossref
O’Keefe, F.R., Street, H.P., Wilhelm, B.C., Richards, C.D., and Zhu, H. 2011. A new skeleton of the cryptoclidid plesiosaur Tatenectes laramiensis reveals a novel body shape among plesiosaurs. Journal of Vertebrate Paleontology 31: 330–339. Crossref
Otero, R.A., Alarcón-Muñoz, J., Soto-Acuña, S., Rojas, J., Rojas, O., and Ortíz, H. 2020. Cryptoclidid plesiosaurs (Sauropterygia, Plesiosauria) from the Upper Jurassic of the Atacama Desert. Journal of Vertebrate Paleontology 40 (1): e1764573. Crossref
Owen, R. 1860. On the orders of fossil and recent Reptilia, and their distribution through time. Report of the British Association for the Advancement of Science 29: 153–166.
Pavlow, A.P. 1914. Jurassic and Lower Cretaceous Cephalopoda of Northern Siberia [in Russian]. Zapiski Imperatorskoj Akademii Nauk po Fiziko-Matematičeskomu Otdeleniû, Ser. 8 = Mémoires de l’Académie Impériale des Sciences, Classe Physico-Mathématique, Sér. 8 31: 1–68.
Pestchevitskaya, E., Lebedeva, N., and Ryabokon, A. 2011. Uppermost Jurassic and lowermost Cretaceous dinocyst successions of Siberia, the Subarctic Urals and Russian Platform and their interregional correlation. Geologica Carpathica 62: 189–202. Crossref
Poulton, T.P. 1994. Jurassic stratigraphy and fossil occurrences—Melville, Prince Patrick, and Borden Islands. Geological Survey of Canada Bulletin 450: 161–193. Crossref
Roberts, A.J., Druckenmiller, P.S., Cordonnier, B., Delsett, L.L., and Hurum, J.H. 2020. A new plesiosaurian from the Jurassic–Cretaceous transitional interval of the Slottsmøya Member (Volgian), with insights into the cranial anatomy of cryptoclidids using computed tomography. PeerJ 8: e8652. Crossref
Roberts, A.J., Druckenmiller, P.S., Delsett, L.L., and Hurum, J.H. 2017. Osteology and relationships of Colymbosaurus Seeley, 1874, based on new material of C. svalbardensis from the Slottsmøya Member, Agardhfjellet Formation of central Spitsbergen. Journal of Vertebrate Paleontology 37 (1): e1278381. Crossref
Rogov, M.A. and Zakharov, V.A. 2009. Ammonite- and bivalve-based biostratigraphy and Panboreal correlation of the Volgian Stage. Science in China, Series D: Earth Sciences 52: 1890–1909. Crossref
Sachs, S. and Kear, B.P. 2017. Redescription of the elasmosaurid plesiosaurian Libonectes atlasense from the Upper Cretaceous of Morocco. Cretaceous Research 74: 205–222. Crossref
Sachs, S., Eggmaier, S., and Madzia, D. 2024. Exquisite skeletons of a new transitional plesiosaur fill gap in the evolutionary history of plesiosauroids. Frontiers in Earth Science 12: 1341470. Crossref
Sachs, S., Hornung, J.J., and Kear, B.P. 2016. Reappraisal of Europe’s most complete Early Cretaceous plesiosaurian: Brancasaurus brancai Wegner, 1914 from the “Wealden facies” of Germany. PeerJ 4: e2813. Crossref
Sachs, S., Hornung, J.J., Lallensack, J.N., and Kear, B.P. 2018. First evidence of a large predatory plesiosaurian from the Lower Cretaceous non-marine “Wealden facies” deposits of northwestern Germany. Alcheringa: An Australasian Journal of Palaeontology 42: 501–508. Crossref
Sachs, S., Kear, B.P., and Everhart, M.J. 2013. Revised vertebral count in the “longest-necked vertebrate” Elasmosaurus platyurus Cope 1868, and clarification of the cervical-dorsal transition in Plesiosauria. PLoS ONE 8 (8): e70877. Crossref
Schneider, S., Kelly, S.R.A., Mutterlose, J., Herrle, J.O., Hülse, P., Jolley, D.W., Schröder-Adams, C.J., and Lopez-Mir, B. 2020. Macrofauna and biostratigraphy of the Rollrock Section, northern Ellesmere Island, Canadian Arctic Islands—a comprehensive high latitude archive of the Jurassic–Cretaceous transition. Cretaceous Research 114: 1–35. Crossref
Seeley, H.G. 1874. Note on some of the Generic Modifications of the Plesiosaurian Pectoral Arch. Quarterly Journal of the Geological Society of London 30 (1–4): 436–449. Crossref
Stott, D.F. 1969. Ellef Ringnes Island, Canadian Arctic Archipelago. Geological Survey of Canada Paper 68: 1–44. Crossref
Tennant, J.P., Mannion, P.D., and Upchurch, P. 2016. Sea level regulated tetrapod diversity dynamics through the Jurassic/Cretaceous interval. Nature Communications 7: 1–11. Crossref
Thorsteinsson, R. 1971. Geology, Slidre Fiord, District of Franklin. Map 1298A. Geological Survey of Canada, Ottawa. Crossref
Tozer, E.T. and Thorsteinsson, R. 1964. Western Queen Elizabeth Islands, Arctic Archipelago. Geological Survey of Canada Memoir 332: 1–242. Crossref
Trautschold, H. 1868. A note on the fossils collected by Barbot-de-Marni at Vychegda [in Russian]. In: N.P. Barbot de Marny (ed.), Geognostičeskoe putešectvie v severnye gubernii Èvropejskoj Rossii, 46–55. Tipografiâ Imperatojskoj Akademii Nauk, St. Petersburg,
Troelsen, J.C. 1952. Geological investigations in Ellesmere Island, 1952. Arctic 5 (4): 198–210. Crossref
Troelsen, J.C. 1953. A Geological Study of Canyon Fiord, Ellesmere Island, with Particular Reference to the Geological History of the Franklinian Geosyncline. Progress Report. Project No. ONR-73. 5 pp. The Arctic Institute of North America, Calgary. Crossref
Troelsen, J.C. 1954. A Geological Study of Canyon Fiord, Ellesmere Island, With Particular Reference to the Geological History of the Franklinian Geosyncline. Project No. ONR-73. 24 pp. The Arctic Institute of North America, Calgary.
Tyborowski, D. and Błażejowski, B. 2019. New marine reptile fossils from the Late Jurassic of Poland with implications for vertebrate faunas palaeobiogeography. Proceedings of the Geologists’ Association 130: 741–751. Crossref
van der Meer, D.G., Scotese, C.R., Mills, B.J.W., Sluijs, A., van den Berg van Saparoea, A.-P., and van de Weg, R.M.B. 2022. Long-term Phanerozoic global mean sea level: Insights from strontium isotope variations and estimates of continental glaciation. Gondwana Research 111: 103–121. Crossref
Voronets, N.S. 1962. Stratigraphy and cephalopods of the Jurassic and Lower Cretaceous deposits of the Leno-Anabar region [in Russian]. Trudy Naučno-Issledovatel’skogo Instituta Geologii Arktiki; Ministerstva Geologii i Ochrany NEDR SSSR 110: 1–237.
Welles, S.P. 1943. Elasmosaurid plesiosaurs with description of new material from California and Colorado. Memoirs of the University of California 13 (3): 125–217.
Wierzbowski, A., Hryniewicz, K., Hammer, Ø., Nakrem, H.A., and Little, C.T.S. 2011. Ammonites from hydrocarbon seep carbonate bodies from the uppermost Jurassic–lowermost Cretaceous of Spitsbergen and their biostratigraphical importance. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 262: 267–288. Crossref
Wilhelm, B.C. and O’Keefe, F.R. 2010. A new partial skeleton of a cryptocleidoid plesiosaur from the Upper Jurassic Sundance Formation of Wyoming. Journal of Vertebrate Paleontology 30: 1736–1742. Crossref
Williston, S.W. 1925. Osteology of the Reptiles. 300 pp. Harvard University Press, Cambridge. Crossref
Wintrich, T., Hayashi, S., Houssaye, A., Nakajima, Y., and Sander, P.M. 2017. A Triassic plesiosaurian skeleton and bone histology inform on evolution of a unique body plan. Science Advances 3 (12): e1701144. Crossref
Zakharov, V.A. 1981. Buchiidae and biostratigraphy of the Boreal Upper Jurassic and Neocomian [in Russian]. Trudy Instituta Geologii i Geofiziki, Akademija Nauk SSR, Sibirskoe Otdelenie 458: 1–270.
Zakharov, V.A. and Rogov, M.A. 2020. High-resolution stratigraphy of buchiid bivalves and ammonites from the Jurassic–Cretaceous boundary beds in the Paskenta area (California). Cretaceous Research 110: 104422. Crossref
Zakharov, V.A., Meledina, S.V., and Shurygin, B.N. 2003. Paleobiochores of Jurassic Boreal basins. Russian Geology and Geophysics 44: 664–675.
Zverkov, N.G. and Prilepskaya, N.E. 2019. A prevalence of Arthropterygius (Ichthyosauria: Ophthalmosauridae) in the Late Jurassic—earliest Cretaceous of the Boreal Realm. PeerJ 7: e6799. Crossref
Zverkov, N.G., Fischer, V., Madzia, D., and Benson, R.B.J. 2018. Increased pliosaurid dental disparity across the Jurassic–Cretaceous transition. Palaeontology 61: 825–846. Crossref
Acta Palaeontol. Pol. 69 (4): 565–585, 2024
https://doi.org/10.4202/app.01148.2024