
The affinities of Afrophoca libyca from basal Middle Miocene of Gebel Zelten, Libya
MARTIN PICKFORD and CHRISTIAN DE MUIZON
Re-interpretation of the holotype and only known specimen of Afrophoca libyca reveals that it represents a medium-sized anthracothere, Afromeryx zelteni, a species that is common in the basal Middle Miocene deposits at Gebel Zelten, Libya. This re-identification affects several recently published papers that have accepted it as the earliest known phocid, with repercussions on biogeographic scenarios and phylogeny reconstructions.
Introduction
The holotype, and only known specimen, of Afrophoca libyca Koretsky & Domning, 2014, is a right mandible fragment (ELNRP 2Z131) extending from the posterior wall of the canine alveolus mesially to the posterior alveolus of the m1 distally. It retains almost complete but damaged p3 and p4 and the anterior root of the m1. The specimen is from Gebel Zelten (also known as Jabal Zaltan), Libya, a basal Middle Miocene locality (Koretsky and Domning 2014). Afrophoca libyca has been interpreted by these authors as the oldest known representative of the family Phocidae in the Old World, and included in the subfamily Monachinae.
However, examination of the fossil reveals that it belongs, in fact, to the medium-sized anthracothere, Afromeryx zelteni, Pickford, 1991, of which the type locality is also Gebel Zelten. The attribution of the holotype and only known specimen of Afrophoca libyca to Afromeryx zelteni impacts on recent literature that accepted the original identification of Koretsky and Domning (2014) rendering the biogeographic and biochronological scenarios obsolete, and the phylogenetic analyses of no relevance for understanding the origins of Phocidae.
Institutional abbreviations.—ELNRP, East Libya Neogene Research Project, University of Garyounis, Benghazi, Libya; MNHN, Muséum national d’Histoire naturelle, Paris, France; NHMUK, Natural History Museum of the United Kingdom, London, UK; USNM, National Museum of Natural History, Washington, DC, USA.
Material and methods
A surface scan and published images of the holotype of Afrophoca libyca were compared with original fossils of anthracotheres from Gebel Zelten curated in the NHMUK and MNHN.
We follow standard convention in abbreviating tooth families as I, C, P, and M, with upper and lower case letters referring to upper and lower teeth, respectively.
Results
Several features that support the attribution of ELNRP 2Z131 to Afromeryx zelteni, that is common at Gebel Zelten, serve to distance it from Phocidae (Figs. 1–3). The seven features labelled in Fig. 1 are discussed below.
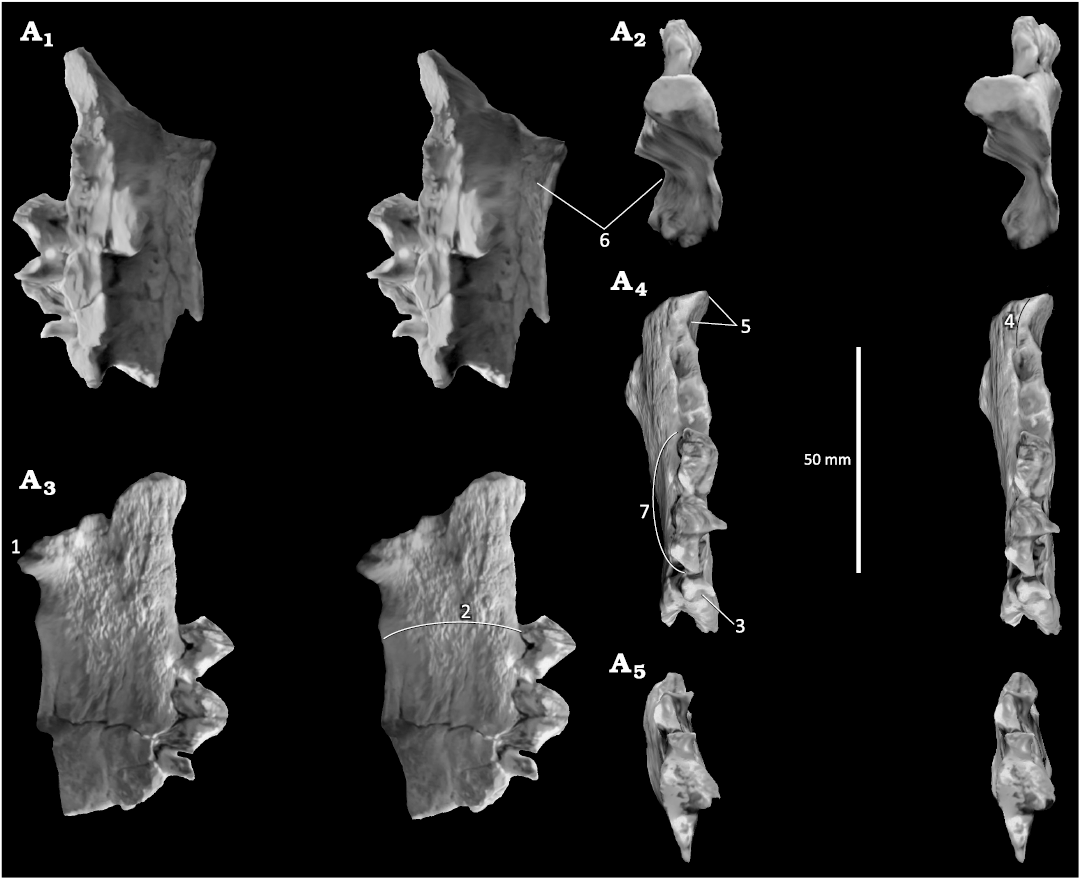
Fig. 1. Stereo images extracted from a scan of the holotype of Afrophoca libyca Koretsky & Domning, 2014, from basal Middle Miocene of Gebel Zelten, Libya. Mandible (ELNRP 2Z131) in labial (A1), anterior (A2), lingual (A3), occlusal (A4), and posterior (A5) views. 1–7 correspond to characters discussed in the text.
1.—The genial spine of the mandible (= mental process in Koretsky and Domning 2014) is prominent and is positioned beneath the p3. The rear of the symphysis (which is damaged in the holotype, leaving some doubt about its original extent) is reported to terminate beneath the p2 (beneath p1–p2 in Afromeryx). The presence of a genial spine in anthracotheres is related to the early fusion of the dentaries at the symphysis. Phocids, unlike anthracotheres, have no genial spine since their dentaries are not fused at the symphysis but are held together by ligaments.
2.—The reported extreme depth and thickness of the mandible of Afrophoca libyca fall within the range of variation of Afromeryx zelteni. Phocid mandibles are generally considerably shallower and more slender (personal observation), with the exception of the massive mandible of the durophagous monachine Hadrokirus martini Amson & Muizon, 2014, from the latest Miocene of Peru.
3.—The remnant of the m1 in the holotype of Afrophoca libyca indicates that its anterior root is wider than long (i.e., it is oval in section and transversely elongated); furthermore, the root has a reniform outline because of the presence of a deep posterior groove. Therefore, the morphology of the anterior root of the m1 of Afrophoca libyca suggests that it consists of two medio-laterally coalescent roots. This condition is also observed in Afromeryx zelteni, whereas, the single anterior and posterior roots of phocid m1 have circular outlines. Furthermore, the anterior alveolus of the m1 of Afrophoca libyca is narrower labio-lingually than that of the p4, a condition, which is also typical of Afromeryx zelteni.
4.—There is a diastema between the alveoli of the cannine and the p1 that was estimated to be about 10 mm long from the figure in Koretsky and Domning (2014: fig. 1E) and ca. 11 mm from the scaled scan. In Afromeryx zelteni the diastema is 11–12 mm in length according to Pickford (1991). In phocids there is generally no diastema between the canine and the p1 but, when present, it is shorter than the length of the alveolus of the p1 (e.g., in the holotype of Hadrokirus martini Amson & Muizon 2014. Even in the case of Acrophoca longirostris Muizon, 1981, which has a very long and slender rostrum with long diastemata between all post-canine teeth, no diastema (or a very short one) is observed between C/c and P1/p1, respectively. In contrast, in the holotype jaw of Afrophoca libyca, the diastema is approximately twice as long as the p1 alveolus (p1 alveolus length: 6 mm; diastema length: ca. 11 mm) as is observed in Afromeryx zelteni (Fig. 2).
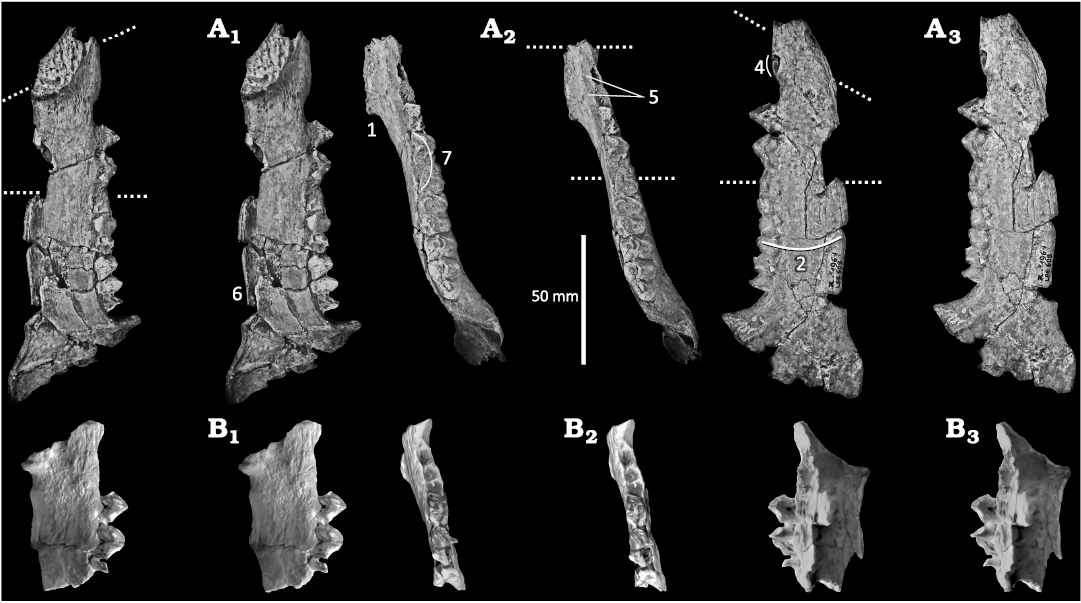
Fig. 2. Comparison between mandibles of Afromeryx zelteni Pickford, 1991 (A, MNHN Z-1961, LBE 608, Gebel Zelten, Libya) and Afrophoca libyca Koretsky & Domning, 2014 (B, ELNRP 2Z131, Gebel Zelten, Libya. Stereopairs in lingual (A1, B1), occlusal (A2, B2), and labial (A3, B3) views. The portion of the Afrophoca mandible corresponding to the jaw of Afromeryx lies between the dotted lines in A. 1–7 correspond to characters discussed in the text. Character 3 which is not visible on MNHN.Z-1961, LBE 608 is illustrated on Fig. 3.
5.—The labial edge of the canine alveolus is strongly reflexed laterally, suggesting the presence of an enlarged canine. Phocids generally possess rather small canines that do not produce such a large deviation of the lateral profile of the mandible (personal observation). Furthermore, as a consequence of the enlargement of the canine, the dentary is deeply excavated labial to the diastema, a condition which is never observed in phocids.
6.—The position of the mental foramina was not reported by Koretsky and Domning (2014) since the labial surface of the holotype mandible of Afrophoca libyca is damaged. However, a capacious mandibular canal is observed beneath the tooth roots (Fig. 1A2). In material of Afromeryx zelteni, mental foramina are positioned beneath p2, p4, and m1, with some variation in precise location and a capacious mandibular canal is present beneath the tooth roots, the latter being typical of anthracotheres in general. In carnivores, in contrast, the mandibular canal is narrow, and the mental foramina are small (slit-like in Phocidae).
7.—In their diagnosis of Afrophoca libyca, Koretsky and Domning (2014: 224–227) stated that the p3 and p4 of the holotype mandible are large, have a sub-rectangular occlusal outline, have a weakly developed basal cingulum and supplementary cheek tooth cusps (in fact, the latter two structures may actually be absent in the holotype of Afrophoca libyca). According to Koretsky and Domning (2014) these four features in Afrophoca libyca differ from all other fossil and extant monachines. Nevertheless, surprisingly, these authors placed Afrophoca within the phocid subfamily Monachinae. In contrast, it is noteworthy that these features perfectly correspond to the premolar morphology observed in the anthracothere Afromeryx zelteni.
The dimensions of the teeth of Afrophoca libyca were estimated by Koretsky and Domning (2014). Table 1 reveals that the published dimensions of the teeth in the holotype of Afrophoca libya (in italics in brackets) contradict the other measurements, especially the length of tooth row p1–p4 (43.5 mm, Koretsky and Domning 2014) and they are inconsistent with the scaled scan. Accepting that the published length of the p1–p4 is accurate, estimates of the lengths of the p3 and p4 can be made. The same can be done with the scan (in bold in Table 1). These measurements plot slightly below the lower end of, or within the range of metric variation of Afromeryx zelteni (Pickford 1991). Note that measurements of the p1 and p2 published by Koretsky and Domning (2014) were estimated from their alveoli and those of the m1 from the damaged remnant of the tooth. For this reason, the measurements are imprecise, because the crowns of mammalian cheek teeth are generally longer and broader than their alveoli. Note also that there is inconsistency between the measurements provided and the scale in their fig. 1 as well as between the dimensions of the p3 and the p4. The published measurements provided by Koretsky and Domning (2014: table 1) indicates that p3 is longer than p4, yet the image clearly shows the opposite (even allowing for the damage that the p4 has suffered).
Table 1. Measurements (in mm) of lower cheek teeth (p3, p4, and m1) of Afromeryx zelteni (maximum and minimum from Pickford 1994) and Afrophoca libyca from Koretsky and Domning 2014 (in brackets). The two measurements of Afrophoca libyca (in bold) have been estimated from the published length of p1–p4 and from the scaled scan, respectively. The latter are possibly the more accurate, but re-measurement of the original fossil is required. Abbreviations: ?, estimation is not possible (tooth damaged); e, estimated from remnant.
|
Tooth |
Afromeryx |
Afrophoca |
Afromeryx zelteni |
Afrophoca libyca |
| |
Length |
Length |
Breadth |
Breadth |
|
p3 |
12.9–13.5 |
12.4–12.9 (11.8) |
8.0 |
6.6 (8.3) |
|
p4 |
14.7–18.5 |
14.2–14.8 (11.4) |
8.9–10.7 |
? (8.0) |
|
m1 |
15.3–17.0 |
? (9.7e) |
8.5–10.9 |
? (7.0e) |
Discussion
Koretsky and Domning (2014) discussed a suite of features that they took to distinguish the holotype mandible of Afrophoca libyca from those of other Phocidae, on which basis the authors erected a new genus and species. All the features listed in the diagnosis (single-rooted p1, rectangular crowns of p3 and p4, presence and position of mental process, depth of mandible, thickness of mandible, absence of diastemata between the cheek teeth, alveolus of m1 narrower than that of p4, and the position of the distal termination of the symphysis beneath p2), some of them being regarded by the authors as unusual for phocids, are all typical of the small anthracothere Afromeryx zelteni Pickford, 1991, which is common at Gebel Zelten.
The presence of a diastema between the lower canine and the p1 is a feature typical of Afromeryx that distinguishes it from the similar sized Sivameryx africanus (Andrews, 1914) that also occurs at Gebel Zelten (Pickford 1991) and which has an exceptionally elongated diastema. Phocids have no diastema between the canine and the p1 or, when present, it is shorter than the length of the alveolus of p1.
In the holotype of Afrophoca libyca, the p1 is single-rooted, and there is no diastema between the p1 and the succeeding cheek tooth. Furthermore, the roots of p2–p4 are separated from each other. If such a condition is almost always observed in phocids (except in Acrophoca), it is also present in Afromeryx and therefore does not differentiate Afrophoca from Afromeryx.
The robust aspect of the teeth and the form of the postero-lateral margin of the canine alveolus, which is strongly reflexed laterally (Koretsky and Domning 2014: fig. 1E) do not resemble the situation in Phocidae. The latter feature seems to indicate a canine of much greater dimensions than those customarily occurring in seals, and as a consequence suggests that the individual was a male (Pickford 2006, 2009). Furthermore, the deep concavity of the dentary labial to the c–p1 diastema is unknown in phocids but is common in anthracotheres.
The dimensions of the holotype jaw of Afrophoca libyca accord with those of Afromeryx zelteni.
On the basis of its morphology and dimensions it is concluded that the holotype jaw of Afrophoca libyca belongs to Afromeryx zelteni, and does not represent a hitherto unknown genus of Phocidae.
Finally, the age of 21 Ma estimated for Gebel Zelten by Koretsky and Rahmat (2013, 2018) and Rahmat et al. (2017) is far too old, the deposits dating from the base of the Middle Miocene ca. 16.5–16.0 Ma (Pickford 1991, 2009).
Phylogenetic, biogeographic and biostratigraphic scenarios based on the assumption that Afrophoca is a seal (Koretsky and Rahmat 2013, 2018; Koretsky and Domning 2014; Berta et al. 2015, 2018; Churchill 2015; Koretsky et al. 2016; Celis 2017; Dewaele et al. 2017, 2018a, b; Koper et al. 2017; Rahmat et al. 2017, 2018; Keil et al. 2022) are hereby rendered obsolete because the holotype and only known specimen of Afrophoca represents the artiodactyl family Anthracotheriidae.
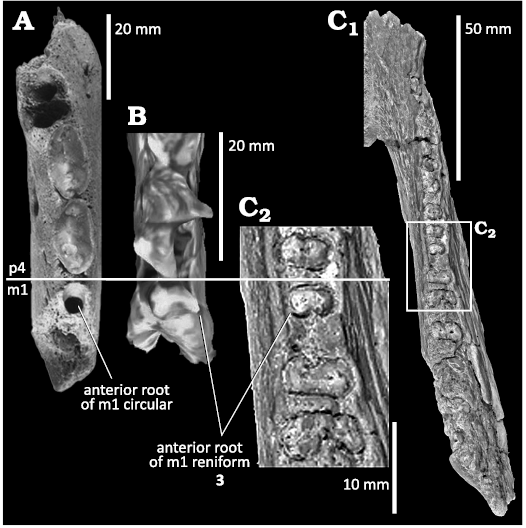
Fig. 3. Comparison between superior views of the anterior root of m1 in a phocid and the anthracotheres from Middle Miocene of Gebel Zelten, Libya. A. Pliophoca etrusca Tavani, 1941, USNM 206176, jaw fragment in occlusal view (from Koretsky and Domning 2014: fig. 1F). B. Afrophoca libyca Koretsky & Domning, 2014, ELNRP 2Z131 (holotype) in occlusal view. C. Afromeryx zelteni Pickford, 1991, NHMUK PV M 82236 right mandible (X66 in Pickford 1991: pl. 10: 3), superior view of the complete specimen (C1) and the part enlarged for comparative purposes (C2). Number 3 corresponds to character discussed in the text.
Conclusions
The holotype mandible fragment of Afrophoca libyca is shown to belong to a small anthracothere, Afromeryx zelteni Pickford, 1991, which is common at Gebel Zelten. Afrophoca libyca is therefore a junior synonym of Afromeryx zelteni. This re-identification means that recent discussions on the evolution of Phocidae, which included Afrophoca in the family, require revision.
Acknowledgements.—Thanks to Daryl Domning (Howard University, Washington DC, USA) for providing access to a scan of the holotype of Afrophoca libyca and to Neil Adams (NHMUK) for the image of Afromeryx zelteni used in Fig. 3 and for curatorial assistance. Thanks also to two anonymous referees whose comments helped improve the discussion.
References
Amson, E. and Muizon, C. de 2014. A new durophagous phocid (Mammalia, Carnivora) from the Neogene of Peru and considerations on monachines phylogeny. Journal of Systematic Palaeontology 12: 523–548. Crossref
Andrews, C.W. 1914. On the lower Miocene vertebrates from British East Africa, collected by Dr Felix Oswald. Quarterly Journal of the Geological Society of London 70: 163–186. Crossref
Berta, A., Churchill, M., and Boessenecker, R.W. 2018. The origin and evolutionary biology of pinnipeds: seals, sea lions, and walruses. Annual Review of Earth and Planetary Sciences 46: 203–228. Crossref
Berta, A., Kienle, S., Bianucci G., and Sorbi, S. 2015. A reevaluation of Pliophoca etrusca (Pinnipedia, Phocidae) from the Pliocene of Italy: phylogenetic and biogeographic implications. Journal of Vertebrate Paleontology 45 (1): e889144. Crossref
Churchill, M. 2015. Fossil Focus: Seals, sea lions and walruses. Palaeontology Online 5: art. 9.
De Celis, A. 2017. Fossil calibrations and mitochondrial genes provide new data about divergence timing of pinnipeds (Carnivora, Pinnipedia). In: S. Barrios de Pedro, C.B. Moreno, A. de Celis, J. Colmenar, E. Cuesta, D.G. Martínez, F. Gascó, A. Jacinto, E. Malafaia, M.M. Jiménez, C. de Miguel Chaves, P. Mocho, V. Pais, A.P. Blázquez, S. Pereira, A.S. Martínez, and D. Vida (eds.), A Glimpse of the Past. In: Abstract Book of the XV Encuentro de Jóvenes Investigadores en Paleontología, 131–134. XV Encontro de Jovens Investigadores em Paleontologia, Pombal.
Dewaele, L., Lambert, O., and Louwye, S. 2017. On Prophoca and Leptophoca (Pinnipedia, Phocidae) from the Miocene of the North Atlantic realm: redescription, phylogenetic affinities and paleobiogeographic implications. PeerJ 5: e3024. Crossref
Dewaele, L., Lambert, O., and Louwye, S. 2018a. A critical revision of the fossil record, stratigraphy and diversity of the Neogene seal genus Monotherium (Carnivora, Phocidae). Royal Society Open Science 5 (5): 171669. Crossref
Dewaele, L., Peredo, C.M., Meyvisch, P., and Louwye, S. 2018b. Diversity of late Neogene Monachinae (Carnivora, Phocidae) from the North Atlantic, with the description of two new species. Royal Society of Open Science 5 (3): 172437. Crossref
Kiel, S., Goedert, J.L., and Tsai, C.-H. 2022. Seals, whales and the Cenozoic decline of nautiloid cephalopods. Journal of Biogeography 49: 1903–1910. Crossref
Koper, L., Rahmat, S., and Koretsky, I. 2017. Origin and dispersal of true seals (family Phocidae) based on recent fossil evidence. In: 77th Annual Meeting Program & Abstracts, Poster Session IV, 143–144. Society of Vertebrate Paleontology, Calgary.
Koretsky, I.A. and Domning, D.P. 2014. One of the oldest seals (Carnivora, Phocidae) from the Old World, Journal of Vertebrate Paleontology 34: 224–229. Crossref
Koretsky, I.A. and Rahmat, S. 2013. First record of fossil Cystophorinae (Carnivora, Phocidae): Middle Miocene seals from the Northern Paratethys. Rivista Italiana di Paleontologia e Stratigrafia 119: 325–350.
Koretsky, I.A. and Rahmat, S. 2018. Origin and dispersal of true seals (family Phocidae) based on Recent fossil evidence. In: 5th International Palaeontological Congress, Abstracts Volume, 1–84. IPC5, Paris.
Koretsky, I.A., Barnes, L.G., and Rahmat, S.J. 2016. Re-evaluation of morphological characters questions current views of pinniped origins. Vestnik zoologii 50: 327–354. Crossref
Muizon, C. de 1981. Les vertébrés fossiles de la Formation Pisco (Pérou). Première partie: Deux nouveaux Monachinae (Phocidae, Mammalia) du Pliocène de Sud-Sacaco. Institut Français d’Études Andines, Mémoire 6: 20–161.
Pickford, M. 1991. Revision of the Neogene Anthracotheriidae of Africa. In: M.J. Salem, O.S. Hammuda, and M.T. Eliagoubi (eds.), The Geology of Libya, Vol. 4, 1491–1525. Elsevier, Amsterdam.
Pickford, M. 1994. Anthracotheriidae from the Albertine Rift Valley. In: B. Senut and M. Pickford (eds.), Geology and Palaeobiology of the Albertine Rift Valley, Uganda-Zaire. Vol. 2: Palaeobiology-Paléobiologie. Occasional Publication CIFEG Orléans 29: 309–319.
Pickford, M. 2006. Sexual and individual morphometric variation in Libycosaurus (Mammalia, Anthracotheriidae) from the Maghreb and Libya. Geobios 39: 267–310. Crossref
Pickford, M. 2009. Metric variation in Afromeryx and Libycosaurus (Anthracotheriidae: Mammalia) and its utility for biochronology. Revista Española de Paleontologia 24: 107–120. Crossref
Rahmat, S.J., Koretsky, I.A., Osborne, J.E., and Alford, A.A. 2017. New Miocene Monachinae from the Western Shore of Chesapeake Bay (Maryland, USA). Vestnik zoologii 51: 221–242. Crossref
Rahmat, S.J., Muniz, F., Toscana, A., Esperante, R., and Koretsky, I. 2018. First European record of Homiphoca (Phocidae: Monacinae: Lobodontini) and its bearing on the paleobiogeography of the genus. Historical Biology 32: 561–569. Crossref
Tavani, G. 1941. Revisione dei resti del Pinnipede conservato nel Museo di Geologia di Pisa. Palaeontographia Italica 40: 97–113.
Martin Pickford [martin.pickford@mnhn.fr; ORCID: https://orcid.org/0000-0002-9017-1107 ] and Christian de Muizon [christian.jourdain-de-muizon@mnhn.fr; ORCID: https://orcid.org/0000-0002-1247-8867 ], Sorbonne Université (CR2P, MNHN, CNRS, UPMC-Paris VI) 8, rue Buffon, 75005, Paris, France.
Received 1 March 2024, accepted 8 April 2024, published online 6 June 2024.
Copyright © 2024 M. Pickford and C. de Muizon. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Acta Palaeontol. Pol. 69 (2): 243–247, 2024
https://doi.org/10.4202/app.01152.2024