

A new symmetrodont mammal from the Early Cretaceous Jehol Biota of Ningcheng Basin, Inner Mongolia, Northeast China
HONGGANG ZHANG, CHANG-FU ZHOU, and ZHE-XI LUO
Zhang, H., Zhou, C.-F., and Luo, Z.-X. 2024. A new symmetrodont mammal from the Early Cretaceous Jehol Biota of Ningcheng Basin, Inner Mongolia, Northeast China. Acta Palaeontologica Polonica 69 (2): 315–327.
Recently, a dinosaur assemblage with three-dimensional preservation of the Early Cretaceous Jehol Biota has been discovered in the Ningcheng Basin, Inner Mongolia, China. Of the vertebrate fossils, a new spalacotherioid “symmetrodont” mammal is described here as Ningchengodon foxi gen. et sp. nov. The specimen is an incomplete right dentary characterized by a lower dental formula of i3/c1/p3/m5, with molar cusps in acute-triangulation. Cusps e and f embraced cusp d of the preceding molar forming a molar interlocking, reported here for the first time for acute-angled spalacotherioids. Phylogenetically, the new taxon is placed in the clade of Spalacotherioidea, and clustered at the base of the paraphyletic zhangheotheriids although its relationship is not further resolved with other zhangheotheriids. Additionally, the ultimate premolar replacement is simultaneous with the m5 eruption as in Origolestes lii and Zhangheotherium quinquecuspidens. This discovery enriches the biodiversity of spalacotherioids, and further highlights the complexity of the Zhangheotheriidae in evolution.
Key words: Symmetrodonta, Spalacotherioidea, Zhangheotheriidae, Ningchengodon foxi gen. et sp. nov., Jehol Biota, Cretaceous, Ningcheng Basin.
Honggang Zhang [13700017969@126.com; ORCID: https://orcid.org/0009-0008-2587-0770 ], College of Earth Science and Engineering, Shandong University of Science & Technology, Qingdao 266590, China; Paleontological Institute, Shenyang Normal University, Shenyang 110034, China; Paleontological Museum of Liaoning, Shenyang 110034, China.
Chang-Fu Zhou [zhoucf528@sdust.edu.cn; ORCID: https://orcid.org/0000-0002-3744-7573 ] (corresponding author), College of Earth Science and Engineering, Shandong University of Science & Technology, Qingdao 266590, China.
Zhe-Xi Luo [zxluo@uchicago.edu; ORCID: https://orcid.org/0000-0003-2170-8879 ] (corresponding author), Department of Organismal Biology and Anatomy, The University of Chicago, Chicago, IL 60637, USA.
Received 6 March 2024, accepted 8 May 2024, published online 19 June 2024.
Copyright © 2024 H. Zhang et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
“Symmetrodont” mammaliaforms are characterized by having a simple reversed-triangle molar pattern in which the three main cusps have a triangular arrangement, and opened labially on upper molars and lingually on lower molars (e.g., Kielan-Jaworowska et al. 2004; Martin 2018). Based on the degree of triangulation, “symmetrodont” mammaliaforms are roughly subdivided into the obtuse-angled and acute-angled morphotypes, which existed from the Late Triassic to the Late Cretaceous (e.g., Kielan-Jaworowska et al. 2004). Mammaliaforms with obtuse-angled molars include Kuehneotheriidae and Tinodontidae, but are paraphyletic (Luo et al. 2002; Lopatin et al. 2005; Martin 2018). By contrast, the acute-angled “symmetrodonts” form a monophyletic clade of the superfamily Spalacotherioidea (including Zhangheotheriidae and Spalacotheriidae), a phylogeny widely supported by previous analyses (e.g., Li and Luo 2006; Sweetman 2008; Bi et al. 2016; Han and Meng 2016).
The spalacotheriids are more derived in having more acute-angled molars, and are more broadly distributed geographically in Laurasia (e.g., Clemens 1963; Fox 1976; Cifelli and Madsen 1986, 1999; Ensom and Sigogneau-Russell 2000; Gill 2004a; Tsubamoto et al. 2004; Hu et al. 2005; Li and Luo 2006; Sweetman 2008; Han and Meng 2016; Mao et al. 2022; Martin et al. 2022). In contrast, the zhangheotheriids are clustered as a basal group within the Spalacotherioidea, and are known from the Early Cretaceous of eastern Asia, especially from western Liaoning, China (Hu et al. 1997, 2005; Rougier et al. 2003; Luo and Ji 2005; Ji et al. 2009; Bi et al. 2016; Plogschties and Martin 2020; Mao et al. 2020; Zhang et al. 2022), but also from western Siberia of Russia (Lopatin et al. 2010). In the Jehol Biota of western Liaoning, four genera and five species have been established, such as Zhangheotherium quinquecuspidens (Hu et al. 1997) and Maotherium sinense (Rougier et al. 2003), which are preserved in two-dimensional slabs, and fossils in three-dimensional preservation of Maotherium asiaticum (Ji et al. 2009), Anebodon luoi (Bi et al. 2016), and Origolestes lii (Mao et al. 2020). The latter species, M. asiaticum, A. luoi, and O. lii, are only known from the Lujiatun beds of the Yixian Formation at the Lujiatun and Jinlingshi sites, famous for diverse fossil vertebrates in three-dimensional preservation. The 3D fossils of mammals offer more valuable anatomical information for our studies of early mammalian evolution (Ji et al. 2009; Zhou et al. 2019; Mao et al. 2020).
Recently, a dinosaur assemblage from the Jehol Biota in the Xidayingzi site in the Ningcheng Basin, Inner Mongolia (Zhang et al. 2022, 2024) has been discovered. This site has also yielded three-dimensionally preserved fossils. The sole mammal fossil from this relatively new site is a partial right dentary, which has been preliminarily assigned to the Zhangheotheriidae (Zhang et al. 2022). Here, we provide a full description of its dental characteristics, to support its status as a new genus and species, and its phylogenetic affinities to the well-known zhangheotheriids.
Institutional abbreviations.—CAGS, Chinese Academy of Geological Sciences, Institute of Geology, Beijing, China; HGM, Henan Geological Museum, Zhengzhou, China; IVPP, Institute of Vertebrate Palaeontology and Palaeoanthropology, Chinese Academy of Sciences, Beijing, China; JZT, Jizantang Paleontological Museum, Chaoyang, China; PMOL, Paleontological Museum of Liaoning, Shenyang Normal University, Shenyang, China; YFGP, Yizhou Fossil & Geology Park, Jinzhou, China.
Other abbreviations.—We follow standard convention in abbreviating tooth families as I, C, P, and M, with upper and lower case letters referring to upper and lower teeth, respectively.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:66AE99DA-09C4-4562-8E05-93C7225F104C
Material and methods
The fossil (PMOL-AM00036; Fig. 1) is a partial right dentary. It is three-dimensionally (3D) preserved and heavily fractured. Due to the fragile condition, the fossil could not be well exposed by manual preparation, but has been CT-scanned at Zhejiang University (Hangzhou, China) using a High-Resolution X-ray CT scanner (Nikon XT H 320). The scanning process was performed without a filter and produced 2500 slices at 100 kV and 100 μA. Voxel size is 0.00687×0.00687×0.00687 mm. The 3D reconstruction was processed using VG Studio 3.0 (Volume Graphics). The comparative specimen of Origolestes lii (JZT-DB0064) was CT-scanned at China University of Geosciences (Beijing, China) using a High-Resolution CT scanner (Nikon XT T225) (Zhou et al. 2019).

Fig. 1. Zhangheotheriid mammal Ningchengodon foxi gen. et sp. nov. (Mammalia, Spalacotherioidea). Holotype (PMOL-AM00036) from the Barremian–Aptian, Lower Cretaceous; Yixian Formation of Xidayingzi, Ningcheng Basin, Inner Mongolia, China. Photography of the partial right dentary with teeth in lingual view, as exposed from rock matrix.
Phylogenetic analysis is based on the data matrix of Bi et al. (2016), which has been updated from Sweetman (2008); the latter was originally derived from Li and Luo (2006) with characters developed by Cifelli and Madsen (1999). Here, we combine the coding of Lactodens sheni (Han and Meng 2016), and add our new taxon and O. lii to the matrix; the coding of O. lii is based on the detailed description from Mao et al. (2020, 2022) and our own CT scans of fossils. The matrix includes 20 taxa and 30 characters (see SOM, Supplementary Online Material available at http://app.pan.pl/SOM/app69-Zhang_etal_SOM.pdf) and was performed with PAUP 4.0b10 (Swofford 1993) under the heuristic search option. All characters are treated as unordered and are equally weighted. Five most parsimonious trees were recovered with a length of 61 steps (consistency index = 0.5738, retention index = 0.8045, rescaled consistency index = 0.4616).
Systematic palaeontology
Mammalia Linnaeus, 1758
Trechnotheria McKenna, 1975
Spalacotherioidea Marsh, 1887
Zhangheotheriidae Rougier et al., 2003
Genus Ningchengodon nov.
Zoobank LSID: urn:lsid:zoobank.org:act:B6DCF54E-BF1B-4EC5-8583-BEDC50A44E86
Etymology: Combination of ningcheng, for Ningcheng Basin of Inner Mongolia where the type locality of Xidayingzi is located, and odon, tooth, a common suffix for fossil mammals.
Type species: Ningchengodon foxi sp. nov., by monotypy, see below.
Diagnosis.—As for the type and only known species.
Stratigraphic and geographic range.—Yixian Formation, Barremian–Aptian (Lower Cretaceous), Ningcheng Basin, Inner Mongolia, China.
Ningchengodon foxi sp. nov.
Figs. 1–10.
Zoobank LSID: urn:lsid:zoobank.org:act:00308FC8-03C1-48F4-802B-4156F7B673EF
Etymology: In honour of the late Professor Richard Carr Fox (1933–2022) for his great contribution to the knowledge of fossil mammals.
Holotype: PMOL-AM00036, a partial right dentary preserved with ultimate premolar and five molars plus tooth loci of other lower premolars, canine and incisors, as represented by tooth alveoli.
Type locality: Duanjiagou, Xidayingzi Village, Bisiyingzi Township, Ningcheng County, Inner Mongolia, China.
Type horizon: Yixian Formation, Barremian–Aptian (Lower Cretaceous).
Diagnosis.—A new zhangheotheriid with a lower dental formula of i3/c1/p3/m5, lack of distinct diastema between premolar loci, ultimate premolar molariform with a symmetrical crown and without cingulid lingually or buccally, acute-triangulation of main cusps of lower molars (m1, 82°; m2, ?; m3, 66°; m4, 88°), cusps e and f of anterior molars (m1–m3) embraced preceding cusp d to form e-f-d interlocking between lower molars m1 through m3, ultimate molar (m5) reduced and obtuse-angled (122°).
Ningchengodon foxi gen. et sp. nov. differs from all known zhangheotheriids (including Anebodon luoi Bi et al., 2016, Maotherium sinense Rougier et al., 2003, Maotherium asiaticum Ji et al., 2009, Origolestes lii Mao et al., 2020, and Zhangheotherium quinquecuspidens Hu et al., 1997) in the absence of major diastemata between the canine and the first premolar and between premolars (Figs. 2–4). It differs from the species of Maotherium and O. lii in having four evenly spaced mental foramina on the lateral side of the dentary (Figs. 3 and 4). It differs from most zhangheotheriids by a more vertical orientation of posterior incisors, canine and anterior premolars (Figs. 2–4). Ningchengodon foxi gen. et sp. nov. differs from O. lii in having three incisors, although this is a plesiomorphic feature of most zhangheotheriids. Ningchengodon foxi gen. et sp. nov. has a symmetrical permanent ultimate premolar (Figs. 4 and 5), different from the asymmetrical ultimate premolar of O. lii. Ningchengodon foxi gen. et sp. nov. differs from all spalacotheriids in lacking a distinct and crest-like lingual cingulid, and in having a molar interlock formed by the distal cusp d of a preceding molar and the mesial cusps e and f of the succeeding molar (Figs. 6 and 7).
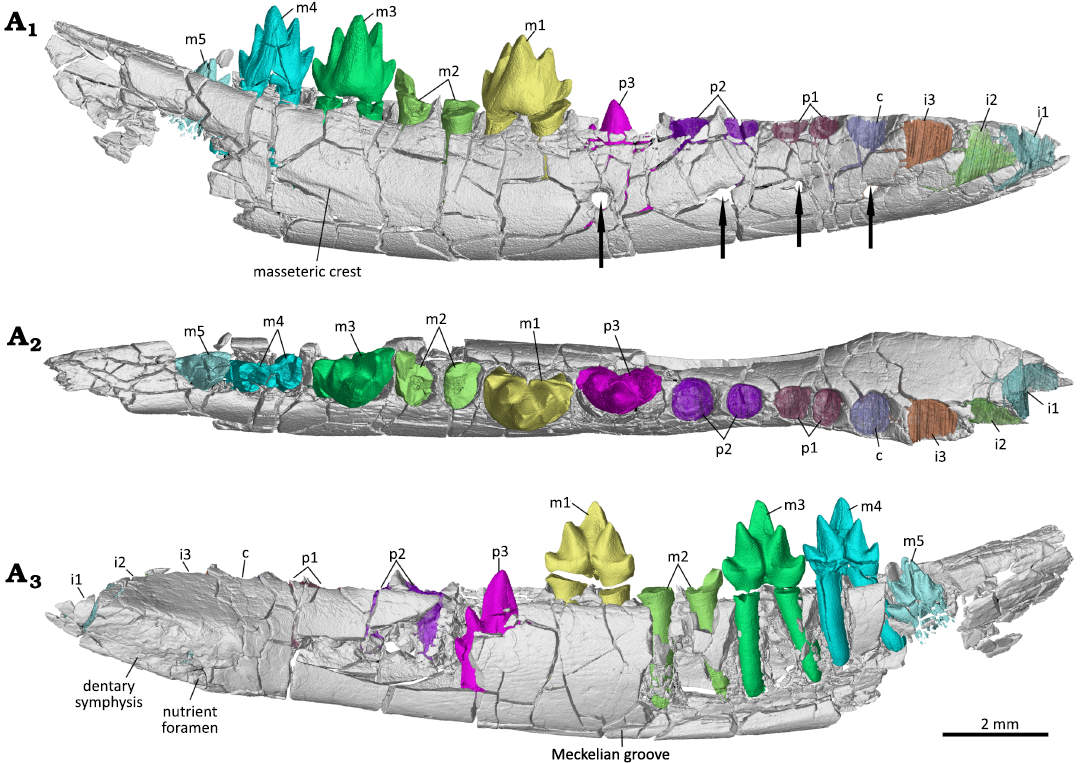
Fig. 2. Digital reconstruction of the partial right dentary with dentition of the zhangheotheriid mammal Ningchengodon foxi gen. et sp. nov. (PMOL-AM00036, holotype) from the Barremian–Aptian, Lower Cretaceous; Yixian Formation of Xidayingzi, Ningcheng Basin, Inner Mongolia, China. Dentary, teeth, and alveoli visualized from CT scans, in labial (A1), occlusal (A2), and lingual (A3) views. The lower m4 crown is separated as found and digitally re-attached to the roots in A1 and A3. Lower m4 crown was repositioned to its roots in A1 and A3, but not in A2. Arrows point to the mental foramina.
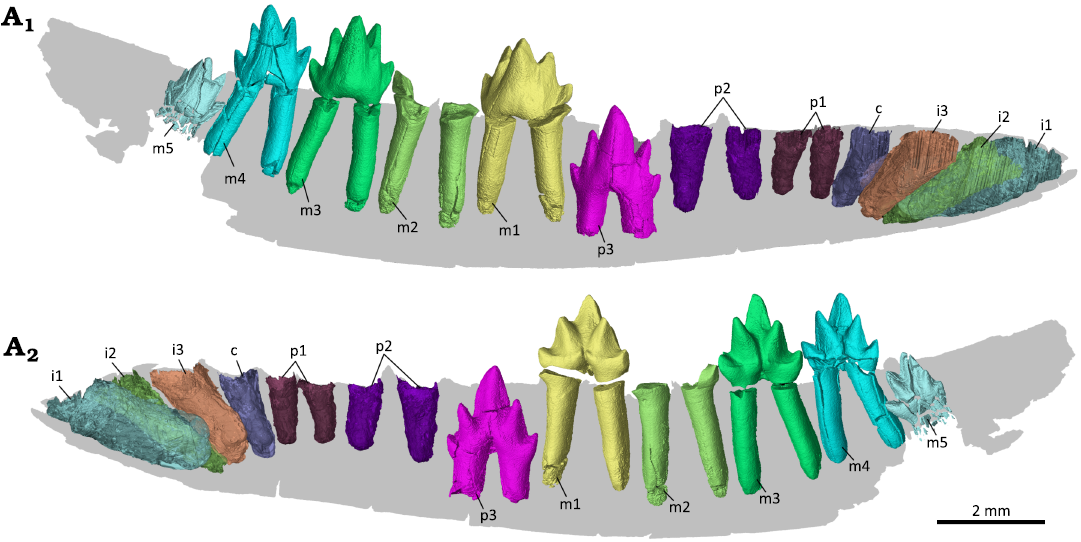
Fig. 3. Digital reconstruction of the dentition of the zhangheotheriid mammal Ningchengodon foxi gen. et sp. nov. (PMOL-AM00036, holotype) from the Barremian–Aptian, Lower Cretaceous; Yixian Formation of Xidayingzi, Ningcheng Basin, Inner Mongolia, China. Teeth and alveoli visualized from CT scans, rendered in labial (A1) and lingual (A2) views. The m4 crown is digitally repositioned to the roots. Individual teeth are illustrated in greater details in Figs. 5–9.
Description.—Dentary: The partial right dentary is preserved with six intact cheek teeth in alveoli (Figs. 2–4), representing a late subadult stage of dental development (details described below). Of the dentary only the horizontal body is preserved. The preserved dentary is fractured, and partially broken. The medial aspect of the dentary body is damaged with some tooth roots exposed. The horizontal body is slender and low, as in Maotherium spp., and slightly more robust than that of O. lii. The ventral margin is convex. The dentary tapers rostrally in lateral view. Its dorsal border or alveolar margin is nearly horizontal, slightly concave, and sculptured by the alveoli of the tooth row. The alveoli are more exposed laterally, because the labial side of alveoli is lower than the lingual side. In occlusal view, the dentary is nearly straight. The symphysis is elongate, oval and procumbent, being set at about 18° relative to the alveolar border. It terminates at the level of the posterior margin of the alveolus of the third incisor position. The symphysis is perforated by a nutrient foramen at its posteroventral corner, as in O. lii (Mao et al. 2020: fig. S6) and Yermakia domitor (Lopatin et al. 2005). The foramen opens anteriorly. Posterior to the symphysis, the dentary is slightly compressed medio-laterally in occlusal view, rather than distinctly constricted as in Maotherium asiaticum (Ji et al. 2009). A thin Meckelian groove is identified under the ultimate premolar and the first molar, and close to the ventral border of the mandible, although the groove along the rest of the mandible cannot be interpreted due to damage. On the labial side of the dentary, four evenly spaced mental foramina are lined along the dentary. The first foramen is positioned under the lower canine alveolus. This mental foramen opens more anteriorly than laterally, and it continues anteriorly as a shallow groove. The groove is fading out antero-dorsally. The other three mental foramina open laterally. The second foramen is slightly smaller than the other foramina, and is positioned in equal distance to the first foramen and the third foramen. The third foramen is slightly larger than the second one. It is positioned under the penultimate premolar. The posterior-most mental foramen is under the ultimate premolar. The foramen is larger than the anterior three foramina. A weak anterior crest of the masseteric fossa is developed, which extends under the third and fourth molars, suggesting an anterior extension of the masseteric fossa on the lateral side of the dentary body. Posterior to the tooth row, the dentary rises gradually, setting an angle of about 22° with the alveolar border, which suggests that an oblique coronoid process was present in this animal, as in other zhangheotheriids.
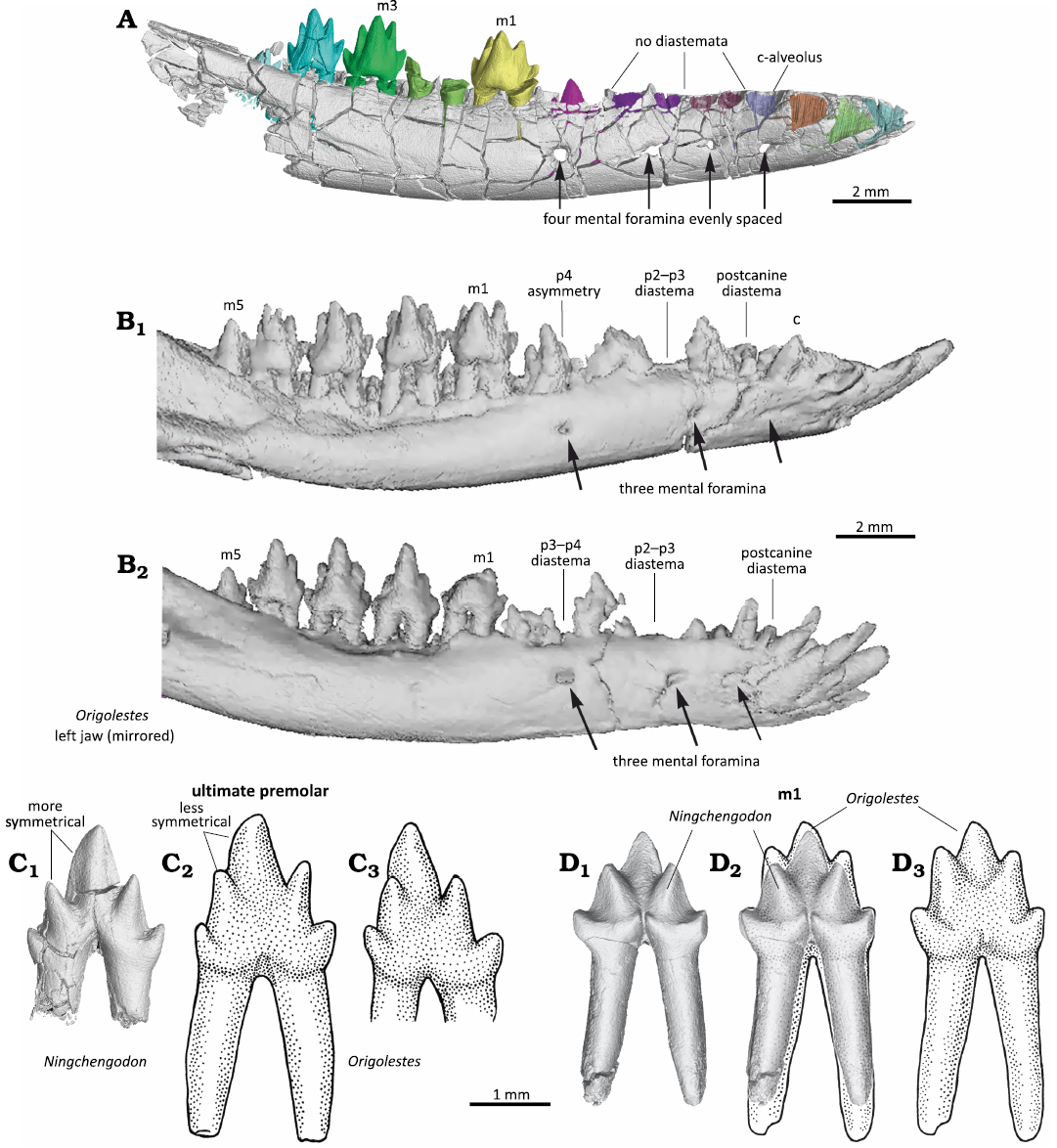
Fig. 4. Comparison of zhangheotheriid mammals Ningchengodon foxi gen. et sp. nov. (PMOL-AM00036) from the Barremian–Aptian, Lower Cretaceous; Yixian Formation of Xidayingzi, Ningcheng Basin, Inner Mongolia, China (A, C1, D1, D2) and selected specimens of Origolestes lii Mao et al., 2020: JZT-DB0064 from Lower Cretaceous of Yixian Formation of Lujiatun, Beipiao, Liaoning, China (B, C3) and IVPP V14383-1 from Lower Cretaceous of Yixian Formation of Lujiatun, Beipiao, Liaoning, China (C2, D2, D3). A. PMOL-AM00036 (holotype); right mandible and teeth in labial view. B. JZT-DB0064; right mandible and teeth in labial view (B1), left mandible in labial view (mirrored, B2). C. Differences in premolars: PMOL-AM00036, right ultimate premolar p3 (C1); IVPP V14383-1 (type specimen), left p4 in lingual view (mirrored, C2); and JZT-DB0064, right p4 in lingual view (C3). D. Differences in first molars: PMOL-AM00036, right m1 in lingual view (D1); PMOL-AM00036 (CT reconstruction), and IVPP V14383-1 (drawing) m1 in lingual views (D2); IVPP V14383-1, left m1 in lingual view (D3). C, D, CT reconstructions and drawings of teeth on same scale, showing m1 of O. lii 4–9% larger than that in N. foxi gen. et sp. nov. A, B, C1, D1, D2, CT reconstructions; C2, C3, D2, D3, stylistic stipple-drawings based on images published by Mao et al. 2020.
Dentition: The whole lower tooth row can be identified by the intact teeth in situ (including un-erupted crowns of p3 and m5) and the empty alveoli for the rest of the teeth that are lost (not preserved). These alveoli represent four single-rooted teeth (three lower incisors and one canine) and two double-rooted teeth (the first and second premolars). The four single-rooted teeth, which are rostrally positioned, are closely comparable to incisors and canine in the known zhangheotheriid species: Anebodon luoi, Maotherium sinense, Maotherium asiaticum, Zhangheotherium quinquecuspidens. In these species, the single-rooted teeth are identified as three incisors and one canine (Hu et al. 1997, 1998; Rougier et al. 2003; Luo and Ji 2005; Ji et al. 2009; Bi et al. 2016).
The anterior teeth of Ningchengodon foxi gen. et sp. nov. (PMOL-AM00036, holotype) have distinct characteristics, by which it is distinguishable from Origolestes lii. In O. lii (Mao et al. 2020), the anterior-most four single-rooted teeth are identified, respectively, as two incisors, one canine (single-rooted), and one single-rooted premolar. The single-rooted lower canine is in occlusion with the two-rooted upper canine in O. lii, and thus the identification of the canine position is well supported by the upper and lower canine crowns and roots that are visualized through CT scans (Mao et al. 2020: fig. S4). Given the position of the single-rooted canine in O. lii, it is a well-justified interpretation that O. lii has two single-rooted incisors anterior to the canine, and a single-rooted first premolar (Mao et al. 2020: fig. S4). In these features, O. lii differs from the specimen of M. asiaticum (HGM 41H-III-0321, holotype), which is also from the Lujiatun site (the same locality as O. lii), as previously discussed (Mao et al. 2020). Additionally, N. foxi gen. et sp. nov. lacks the large diastemata (Fig. 4). By contrast, the premolar diastemata are prominently developed, between canine and p1, between p1 and p2, and between p3 and p4 in the specimens of O. lii (Mao et al. 2020: fig. S4). The diastemata are wider in other known zhangheotheriids, although to a varying degree. For instance, there is a large diastema between p1 and p2 in Z. quinquecuspidens and M. sinense (Hu et al. 1997, 1998; Rougier et al. 2003; Luo and Ji 2005; Plogschties and Martin 2020), or between the canine and the single ultimate premolar in M. asiaticum (Ji et al. 2009: fig. 1). In A. luoi, there are only moderate gaps between p1, p2, and p3 (Bi et al. 2016: fig. 2).
Our identification of three incisors and one canine for N. foxi gen. et sp. nov. is consistent with the features in M. asiaticum, which is characterized by three single-rooted lower incisors, and one single-rooted lower canine matched by a two-rooted upper canine (Ji et al. 2009: fig. 1). The incisor count of N. foxi gen. et sp. nov. and M. asiaticum is a common pattern in zhangheotheriids (Hu et al. 1997, 1998; Rougier et al. 2003; Lopatin et al. 2010; Luo and Ji 2005; Ji et al. 2009; Bi et al. 2016). However, N. foxi gen. et sp. nov. is clearly different from other zhangheotheriids in premolar features in its three two-rooted premolars (indicated by their respective alveoli), including p1 that is closely packed to the canine, without a postcanine diastema. Its full complement of premolars differs from that of M. asiaticum, which has no anterior premolars (although still retaining a single lower premolar) in the ultimate premolar position. Based on this comparison, we determine that the lower dentition of PMOL-AM00036 has three incisors, one canine, three premolars and five molars (i3, c1, p3, m5).
Based on the shape and orientation of the alveoli, the three incisors appear to be large and procumbent. We infer that the first incisor is the largest (based on the largest size of its alveolus), as in most other known zhangheotheriids (Z. quinquecuspidens, Luo and Ji 2005; A. luoi, Bi et al. 2016; and O. lii, Mao et al. 2020), and differing from Kiyatherium cardiodens, in which the first incisor is smaller than the two posterior incisors (Lopatin et al. 2010). Ningchengodon foxi gen. et sp. nov. also differs from M. asiaticum, where the second incisor is much larger than the third (Ji et al. 2009). The canine, as the fourth single-rooted tooth, is more vertically positioned in N. foxi gen. et sp. nov. The canine, third incisor and second incisor form a gradient from the more vertical orientation to less vertical orientation of these teeth (as inferred from the orientation of their alveoli), which is similar to that in M. asiaticum (Ji et al. 2009). But these teeth are distinctive from the more procumbent orientation of the canine and incisors of O. lii (Mao et al. 2020: fig. S4).
Three premolars (p1–p3) are identified. The permanent p3 is still mostly inside the dentary (Figs. 4 and 5), and the two anterior premolars are represented by two pairs of alveoli. The first two paired alveoli are converged toward each other, which we interpret to correspond to the double-rooted p1. The next two alveoli, which appear to be paired, are also convergent dorsally. We interpret these two alveoli to be for p2. The alveoli of the second premolar are stronger, deeper, and more vertically oriented, relative to those of the first premolar. From this we interpret that the second premolar was larger, and slightly more vertical than the first premolar. These premolar alveoli are aligned longitudinally to those of the incisors and canine, and they are closely packed, indicating the lack of distinct diastemata. As expected from other zhangheotheriids for which the root morphologies are known, the alveoli of the two premolars are shallower than the alveoli of the incisors and canine, and also shorter than the roots of the molars (e.g., O. lii, Mao et al. 2020).
The ultimate lower premolar (p3) is in the process of eruption and just about to break through the alveolar margin (Figs. 2–5). Its crown is molariform (Fig. 5). The crown is symmetric mesio-distally and compressed bucco-lingually. The main cusps are well developed. Of these, the protoconid (cusp a) is much larger than the paraconid (cusp b) and the metaconid (cusp c); the latter two are comparable in size, and slightly lingually positioned relative to the protoconid, forming an obtuse triangle of 129°. Cusp a is robust and buccally more convex than lingually. Its tip is slightly curved distally and two crests extend down from the tip in mesial and distal direction. The cusps b and c are conical, and bucco-lingually more expanded than mesio-distally. Cusp b is well separated from cusp a. Cusp c is positioned slightly higher than cusp b, and moderately separated from cusp a. The hypoconid (cusp d) and mesial cusp e are present. They are nearly aligned mesio-distally with the cusps b and c. Cusp d is slightly larger than cusp e. A small and short crest extends buccally from cusp d. The base (or cingular part) of the crown is smooth and lacks a cingulid on both the lingual side and buccal side of the crown, distinct from the weak continuous lingual cingulid in A. luoi and O. lii (Bi et al. 2016; Mao et al. 2020). The doubled roots of the growing tooth are not yet fully formed and only developed to about half of the crown height. The roots are divergent and the root tips are open. As in Maotherium sinense (YFGP 1724; Plogschties and Martin 2020), the anterior root is nearly cylindrical, while the posterior root is mesio-distally compressed to form an oval outline in horizontal section.
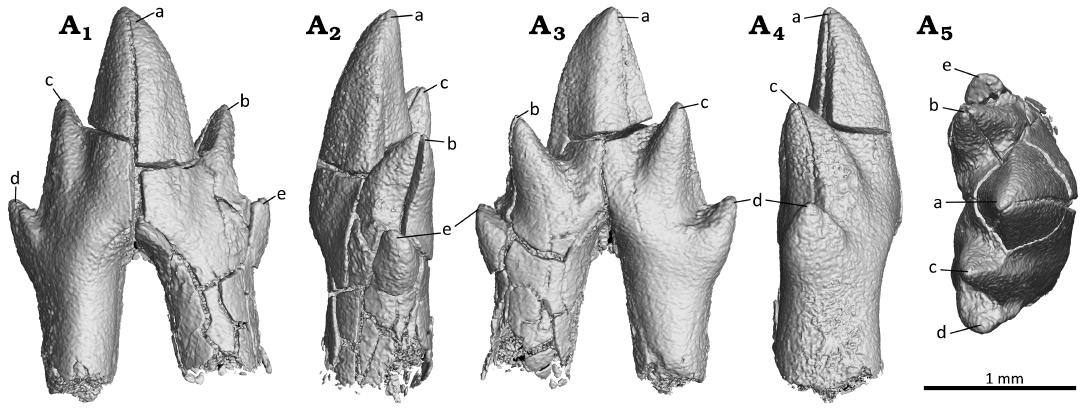
Fig. 5. Digital reconstruction of the ultimate premolar (p3) of the zhangheotheriid mammal Ningchengodon foxi gen. et sp. nov. (PMOL-AM00036, holotype) from the Barremian–Aptian, Lower Cretaceous; Yixian Formation of Xidayingzi, Ningcheng Basin, Inner Mongolia, China. Tooth in labial (A1), mesial (A2), lingual (A3), distal (A4), and occlusal (A5) views.
Five molars (m1–m5) are preserved in situ (Figs. 6–10), but the second molar (m2) has lost its crown. The crown of m4 is broken off, but could be re-associated back to its two roots, and in original position (Figs. 2–4, 8). The m5 is not yet fully developed with cusp a just breaching the dentary bone. The preserved crowns of the molars (m1, m3, m4, and m5) are bucco-lingually wide, but still longer than wide (Table 1). Cusps a, b, c, d, and e are cone-like and well separated from one another. These cusps are mesio-distally symmetric relative to the cusp a in m1, m3, and m4. In m1, cusps b and c are comparable in size, while cusp b is higher than cusp c in m3 and m4. Moreover, the main cusps form an acute trigonid in m1, m3, and m4 (Table 1). As in A. luoi and O. lii, the trigonid of the first molar (m1) has an acute angle about 82°, different from the obtuse-angled condition in K. cardiodens, M. sinense, M. asiaticus, and Z. quinquecuspidens (Hu et al. 1997; Luo and Ji 2005; Ji et al. 2009; Lopatin et al. 2010; Plogschties and Martin 2020). The trigonid angle decreases in the third molar (m3) to about 66°, and then increases in the fourth molar (m4) to 88°. Finally, in the ultimate molar (m5), the trigonid angle becomes obtuse at 122°. The b-a-c triangle on m5 is more asymmetrical than that of preceding molar.
Table 1. Measurements (in mm) of the lower premolar and molars of Ningchengodon foxi gen. et sp. nov. (PMOL-AM00036). ?, unknown.
|
Tooth |
Width |
Length |
Trigonid angle |
|
p3 |
0.82 |
1.71 |
129° |
|
m1 |
1.02 |
1.71 |
82° |
|
m2 |
? |
? |
? |
|
m3 |
1.02 |
1.65 |
66° |
|
m4 |
0.84 |
1.48 |
88° |
|
m5 |
0.72 |
1.17 |
122° |
The lower molar interlocking system is a noteworthy feature of the new taxon Ningchengodon foxi. In m1 and m3, the mesial cusp e is large, and it is either subequal to the associated distal cusp d, or even larger. Close to cusp e, an additional mesial cingular cuspule, cusp f, is present and positioned buccally to cusp e. For m1 and m3, an interlocking mechanism is developed such that cusp d of the preceding p3 fits in to the mesial notch of m1, or a molar fits into the notch (or embayment) between cusps e and f of the succeeding molar. The m2 has a damaged crown, and only its cusp d is preserved and positioned between cusps e and f of m3. Because the part of the crown where cusps e and f are to be expected is damaged in m2, we can only hypothesize the presence of a cusp d-e-f interlock between m1 and m2.
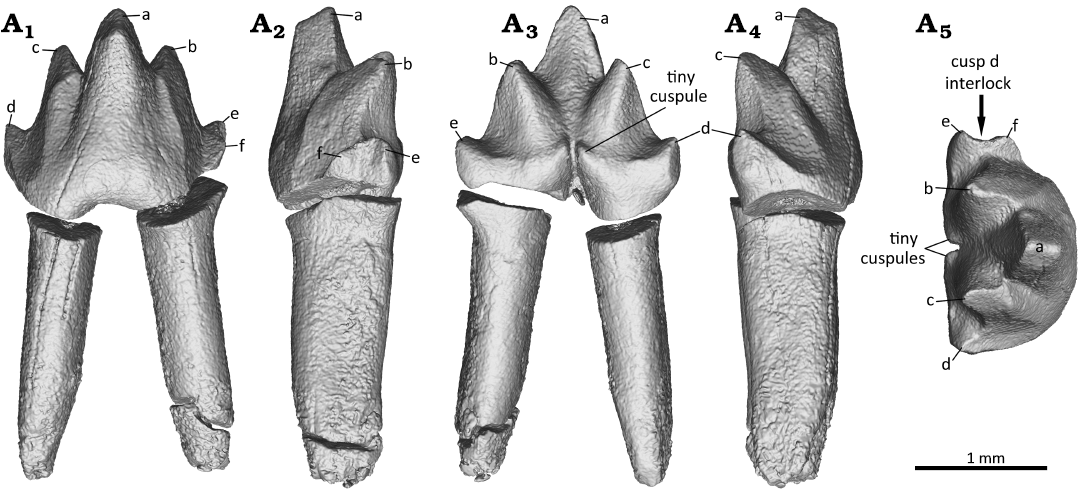
Fig. 6. Digital reconstruction of the right first molar (m1) of the zhangheotheriid mammal Ningchengodon foxi gen. et sp. nov. (PMOL-AM00036, holotype) from the Barremian–Aptian, Lower Cretaceous; Yixian Formation of Xidayingzi, Ningcheng Basin, Inner Mongolia, China. Tooth in labial (A1), mesial (A2), lingual (A3), distal (A4), and occlusal (A5) views.
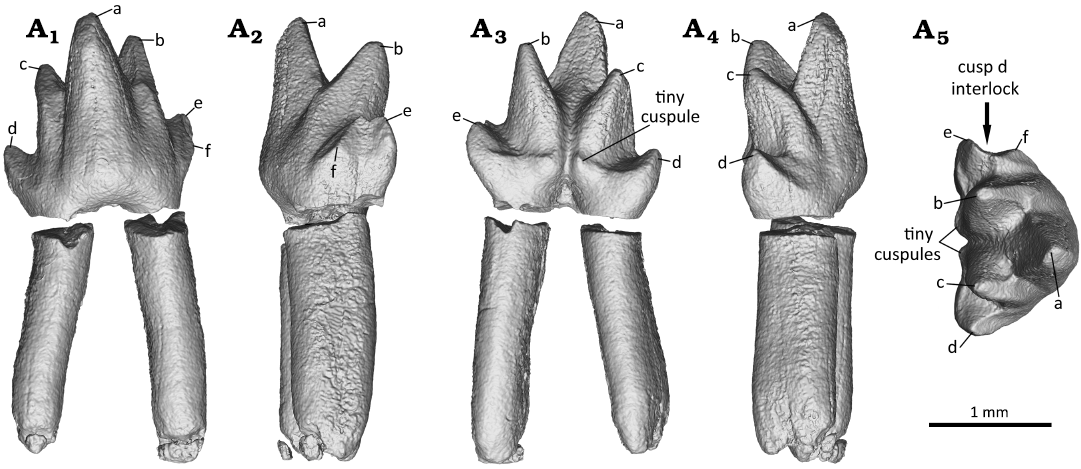
Fig. 7. Digital reconstruction of the right third molar (m3) of the zhangheotheriid mammal Ningchengodon foxi gen. et sp. nov. (PMOL-AM00036, holotype) from the Barremian–Aptian, Lower Cretaceous; Yixian Formation of Xidayingzi, Ningcheng Basin, Inner Mongolia, China. Tooth in labial (A1), mesial (A2), lingual (A3), distal (A4), and occlusal (A5) views.
In m4, the cusp f is reduced to a short blunt ridge, forming a shallow notch with cusp e (Fig. 8). Nearly parallel to the ridge, a comparable crest extends buccally from cusp e. The crest is not developed in m1 and m3; it is either absent, or possibly obliterated by the extensive wear facets between cusps b and f (Fig. 10). In m5, cusp f is vestigial and confluent to the crest extending buccally from cusp e. From the available information, and with caveats, we infer that there is a morphological gradient along the molar row of the molar interlocking: it starts with p3–m1, and is most developed in m1–m3, but its development is progressively less prominent in m4 and m5, toward the posterior end of molar row.
Molar interlocking structure first evolved with some of the early divergent mammaliaforms, such as morganucodontans and kuehneotheriids (e.g., Crompton 1974; Luo 1994; Gill 2004b; Jäger et al. 2019). However, Kuehneotherium species are distinct from other mammaliaforms such as morganucodontans in having cusps d-e-f interlocking, formed as the consequence of development of a neomorphic cusp f (Kielan-Jaworowska et al. 2004). This leads to the imbricating interlock in which cusp d of the preceding molar wedged between cusps e and f of the succeeding molar. Also, a similar interlocking is frequently present in obtuse angled “symmetrodonts” geologically younger than Kuehneotherium species, including Gobiotheriodon infinitus, Tinodon spp., Yermakia domitor (e.g., Kielan-Jaworowska et al. 2004). However, it was either absent, or simply unknown in spalacotherioids, because spalacotherioid species have not been examined for this feature in enough details (Hu et al. 1997; Averianov 2002; Kielan-Jaworowska et al. 2004; Lopatin et al. 2010; Luo and Ji 2005; Ji et al. 2009; Plogschties and Martin 2020; Mao et al. 2020). Ningchengodon foxi gen. et sp. nov. represents the first evidence of the lower molar interlock in a spalacotherioid, and shows a morphological gradient of the interlocking along the molar row, better developed in m1 through m3 (Figs. 6 and 7), than m4 and m5 (Figs. 8 and 9).
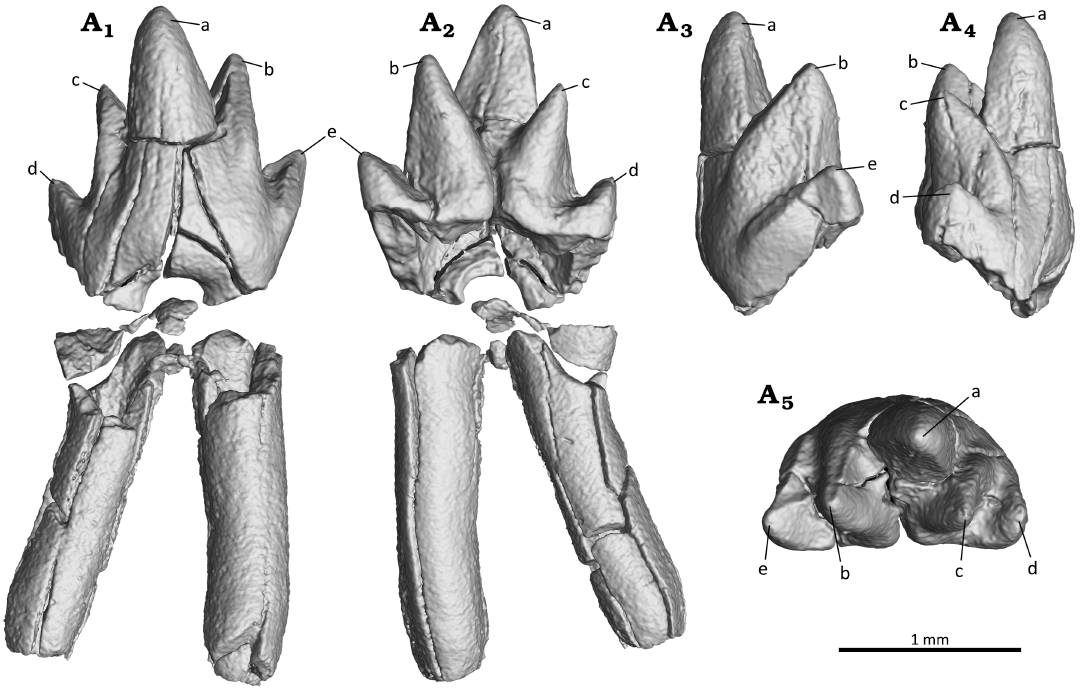
Fig. 8. Digital reconstruction of the fourth molar (m4) of the zhangheotheriid mammal Ningchengodon foxi gen. et sp. nov. (PMOL-AM00036, holotype) from the Barremian–Aptian, Lower Cretaceous; Yixian Formation of Xidayingzi, Ningcheng Basin, Inner Mongolia, China. The crown is digitally re-attached to the roots. Tooth in labial (A1), mesial (A2), lingual (A3), distal (A4), and occlusal (A5) views.
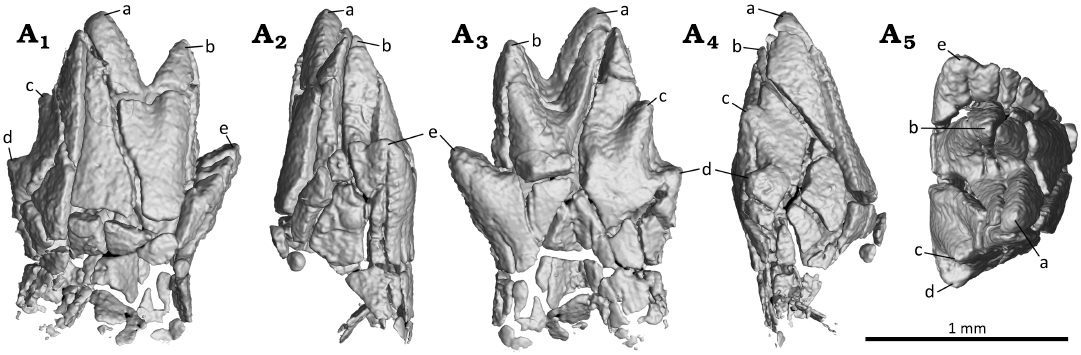
Fig. 9. Digital reconstruction of the ultimate molar (m5) of the zhangheotheriid mammal Ningchengodon foxi gen. et sp. nov. (PMOL-AM00036, holotype) from the Barremian–Aptian, Lower Cretaceous; Yixian Formation of Xidayingzi, Ningcheng Basin, Inner Mongolia, China. The m5 is still in development inside the dentary. Tooth in labial (A1), mesial (A2), lingual (A3), distal (A4), and occlusal (A5) views.
The molar crowns are smooth on the buccal side, and lack a labial cingulid around the base of the crown. On the other side of the crown, the lingual cingulid is present, although weak and discontinuous between two tiny cuspules that are positioned near the mid-length of the crown, respectively associated with the base of cusp b and base of cusp c in m1 and m3. The cuspules are comparable in size and in their symmetrical position near the mid-length of the molar, as the two cuspules in some molars of Z. quinquecuspidens (Hu et al. 1997, 1998; Luo and Ji 2005). A short and distinct sulcus separates the two cuspules, and extends upward to become confluent with the lingually-opened trigonid basin. The tiny cuspules are absent in m4 and m5. A continuous lingual cingulid of the lower molars is developed in K. cardiodens and A. luoi (Lopatin et al. 2010; Bi et al. 2016), but absent in other zhangheotheriids (e.g., Z. quinquecuspidens, Maotherium spp., and O. lii; Hu et al. 1997, 1998; Luo and Ji 2005; Ji et al. 2009; Plogschties and Martin 2020; Mao et al. 2020). A continuous and crest-like lingual cingulid is also present in spalacotheriids (e.g., Cifellitherium suderlandicum, Martin et al. 2022).
The anterior four molars (m1–m4) are double-rooted. The roots are long, straight, and tapered basally. They are compressed mesio-distally to show an oval outline in horizontal section. Paired roots are divergent towards the root tips. The roots are the longest in m2, and become successively shorter in more posterior molars along the molar series. A similar pattern is also present in Maotherium sinense and O. lii (Plogschties and Martin 2020; Mao et al. 2020). This posteriorly decreasing size (length) gradient of molar roots is also present in cladotherian mammals, and is likely because the teeth erupting earlier in development had more time for growth and lengthening the roots, than younger teeth that erupted later in molar row (Luo and Martin 2023).
The ultimate molar (m5) is partially erupted (Figs. 2 and 3). Its crown is complete, but the roots are not yet fully formed (Figs. 3 and 9). Its crown is reduced in size, comparable to the reduced ultimate molar in the relatively complete molar series of other zhangheotheriids, except for A. luoi, in which the ultimate molar is comparable in size with the preceding molars (Bi et al. 2016). The m5 crown is asymmetrical due to the extremely reduced cusps c and d (Fig. 9), which are much smaller than the mesial cusps b and e. Cusp b is also well separated from cusp a, and nearly lined with the latter. Behind m5, no tooth germ could be identified on the CT scan, implying that m5 represents the ultimate molar. The trigonid angle of m5 is obtuse at 122°, which is wider (more obtuse) than m1 and m3. The widening of the trigonid angle in the last one or two teeth also occurs in other zhangheotheriids (Luo and Ji 2005; Chen and Luo 2008; Plogschtie and Martin 2020; Mao et al. 2020). By comparison, in the spalacotheriids, the ultimate lower molars or the last two molars tend to have the three main cusps set at a more acute triangle (Cifelli and Madsen 1999; Kielan-Jaworowska et al. 2004; Chen and Luo 2008; Han and Meng 2016).
Wear facets on the prevallid and postvallid are well developed on the mesio-buccal and disto-buccal side of the first molar (m1; Fig. 10A). The prevallid shearing surfaces are positioned between cusp a and cusp b, and these facets are corresponding to wear facets 2, A, and B on lower molar of Kuehneotherium praecursoris as interpreted by Crompton (1971: fig. 7). Wear facet 2 is apically positioned at cusp a, and oriented nearly vertical along the mesial side of cusp a. This facet is discontinuous from wear facet A that located on the oblique buccal side of cusp b. Facet A is set at an angle with wear facet B. The latter is well developed and located basally between cusps b and f, forming a distinct notch on m1, although this notch was not yet developed on m3. The postvallid shearing surfaces positioned between the taller cusp a and lower cusp c, corresponding to facets 1 and 3 of lower molar of K. praecursoris as interpreted by Crompton (1971). Based on this comparison, we interpret the postvallid facets as wear facets 1 and 3. Wear facet 1 is located on the periapical sides of cusp a and cusp c. Wear facet 3 is positioned distal to facet 1, and is more extensive and positioned basally between cusps c and d, forming a V-shaped outline on m1, although not yet fully developed on m3. In the third molar (m3; Fig. 10B), only wear facet A of the prevallid shearing surface is weakly developed on the buccal side of cusp b. The pattern of discontinuous prevallid shearing surfaces, as evidenced in N. foxi gen. et sp. nov. here, is more widely present in primitive “symmetrodonts”. In contrast, the continuous shearing surface is a diagnostic feature for derived spalacotheriids (e.g., Cifelli and Madsen 1999; Kielan-Jaworowska et al. 2004; Tsubamoto et al. 2004; Martin et al. 2022).
Stratigraphic and geographic range.—Type locality and horizon only.
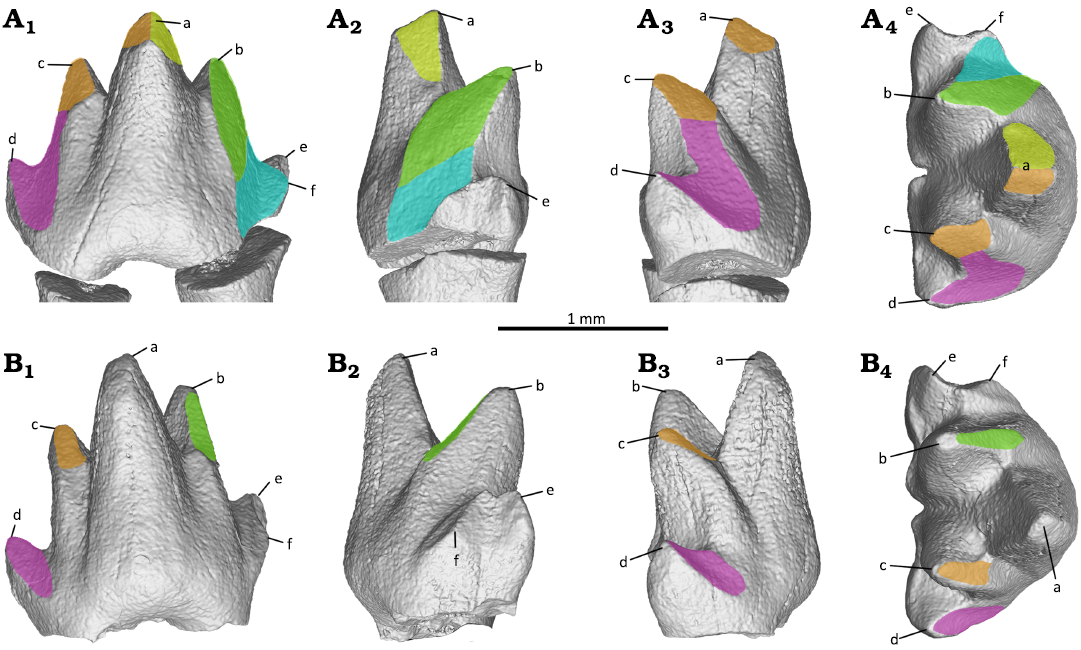
Fig. 10. Tooth wear facets of the first (A) and third (B) molars of the zhangheotheriid mammal Ningchengodon foxi gen. et sp. nov. (PMOL-AM00036, holotype) from the Barremian–Aptian, Lower Cretaceous; Yixian Formation of Xidayingzi, Ningcheng Basin, Inner Mongolia, China. Teeth in labial (A1, B1), mesial (A2, B2), distal (A3, B3), and occlusal A4, B4) views. Brown, wear facet 1; yellow, wear facet 2; green, wear facet A; blue, wear facet B; purple, wear facet 3.
Discussion
The holotype specimen of Ningchengodon foxi gen. et sp. nov. (PMOL-AM00036) was still undergoing development at the time of death, although it already reached a late stage of growth close to adulthood, as evidenced by the partially erupted ultimate premolar (p3) and the ultimate molar (m5) with a formed crown, but not yet fully developed roots.
Phylogenetically, the new taxon N. foxi forms a basal polytomy with other “zhangheotheriids” within the clade of Spalacotherioidea (all of the acute-angled “symmetrodonts”; Fig. 11). Zhangheotheriidae is paraphyletic as in most previous analyses (e.g., Sweetman 2008; Bi et al. 2016; Han and Meng 2016), although its monophyly was supported by some analyses that had sampled fewer taxa (e.g., Sweetman 2008; Mao et al. 2020). Furthermore, N. foxi gen. et sp. nov. has several derived features of dental morphology, and is clearly distinguishable as a new taxon, among “zhangheotheriids” (A. luoi, K. cardiodens, Maotherium spp., O. lii, and Z. quinquecuspidens). For example, N. foxi gen. et sp. nov. lacks a distinct diastema which is common among other “zhangheotheriids”, and plesiomorphic for this group. The ultimate premolar is molariform and lacks a lingual or buccal cingulid, different from the weak lingual cingulid seen in A. luoi and O. lii (Bi et al. 2016; Mao et al. 2020). Ningchengodon foxi gen. et sp. nov. has five molars, one more molar than A. luoi and Kiyatherium cardiodens, and lacks a continuous lingual cingulid typical of the latter taxa (Lopatin et al. 2010; Bi et al. 2016) and of spalacotheriids (Martin et al. 2022); the first molar (m1) is acute-angled, in contrast to the obtuse-angled patterns in K. cardiodens, Maotherium spp., and Z. quinquecuspidens (Lopatin et al. 2010; Luo and Ji 2005; Plogschties and Martin 2020); the molars of N. foxi gen. et sp. nov. are less than half the size of the corresponding teeth in A. luoi (Bi et al. 2016), but comparable with those of other “zhangheotheriids” (Hu et al. 1997; Luo and Ji 2005; Ji et al. 2009; Lopatin et al. 2010; Plogschties and Martin 2020; Mao et al. 2020). More importantly, the molar interlocking pattern of N. foxi gen. et sp. nov., characterized by cusps e and f of the anterior molars embracing cusp d of the preceding p3 for p3–m1 interlocking, and molar in m1–m3 positions (Figs. 6 and 7), is a new finding for the clade of Spalacotherioidea, although it frequently occurs in obtuse-angled “symmetrodonts” such as Gobiotheriodon infinitus, Kuehneotherium spp., Tinodon spp., Yermakia domitor (Luo et al. 2002; Kielan-Jaworowska et al. 2004; Gill 2004b; Lopatin et al. 2005).

Fig. 11. Phylogenetic relationship of Ningchengodon foxi gen. et sp. nov. among spalacotherioids and mammaliaform outgroups. The strict consensus tree of five most parsimonious trees (tree length = 61 steps; consistency index = 0.5738, retention index = 0.8045, rescaled consistency index = 0.4616).
The specimen of N. foxi gen. et sp. nov. shows two features related to diphyodont tooth replacement: (i) its ultimate premolar (p3) is preserved in situ and has erupted partially with cusp a breaching the gum surface, just having breached the alveolar margin; (ii) the replacement of the ultimate premolar (p3) occurs at the same time as the eruption of the ultimate molar (m5). Both these features have also been reported for Z. quinquecuspidens (CAGS97-07352, Luo and Ji 2005). Furthermore, it has been reported for Z. quinquecuspidens that premolar replacement occurred in alternative positions (p1→p3→p2) (Luo and Ji 2005). Because the anterior premolars are not preserved in N. foxi gen. et sp. nov. (Figs. 1 and 2), it is unclear if it had a similar alternating replacement pattern of the premolars as seen in Z. quinquecuspidens (Luo and Ji 2005), or a sequential pattern as in other mammaliaforms (Luo et al. 2004).
In a recent study on O. lii, Mao and colleagues (2020) have illustrated that the permanent tooth co-existed with the deciduous tooth during eruption in the p2 position in one specimen (IVPP V14383-1). In another specimen (JZT-DB0064), the second incisor and the first premolar are in the process of replacement. At the ultimate premolar (p4) locus, the timing of eruption of the permanent premolar may be variable, relative to the eruption of m5 (Mao et al. 2020: fig. S4). In O. lii (JZT-DB0064, Mao et al. 2020: fig. S4), the ultimate premolar is not fully erupted relative to the well-erupted ultimate molar (m5). Meanwhile, the holotype of O. lii (IVPP V14383-1, Mao et al. 2020: fig. S4) shows a slightly different pattern: the ultimate permanent premolar appears to have already erupted and to be fully in place, earlier than the ultimate molar (m5) that is still inside the mandible. Timing of the premolar replacement relative to eruption of the ultimate molar may be more variable among individuals of the same species than previously suspected. Despite this variability, the timing of the ultimate premolar replacement and simultaneous eruption with the m5 eruption is broadly similar in N. foxi gen. et sp. nov., O. lii, and Z. quinquecuspidens. But it is not clear whether N. foxi gen. et sp. nov. and O. lii had the same alternate pattern of premolar replacement as Z. quinquecuspidens, or if their replacement pattern is sequential. Lastly, it appears that in A. luoi, the ultimate premolar is less erupted due to its position lower than the adjacent molars and the ultimate molar (m4) of this species is already fully erupted (Bi et al. 2016: fig. 3). It is possible that A. luoi had a later timing in replacement of the ultimate premolar locus than the eruption of the ultimate molar locus, in comparison to N. foxi gen. et sp. nov. and Z. quinquecuspidens.
Acknowledgements
We thank Qiang Yang, Shurui Yang, Shuai Shao, Peng Yu, and Yuguo Yang (all PMOL) for their help in field excavations and fossil preparation, and Rui Pei (IVPP), Yalei Yin, Yuhui Feng, and Yang Li (all PMOL), and Dongxiang Yu (Shandong University of Science and Technology, Qingdao, China) for their helpful suggestion and discussions in this dinosaur assemblage, and Yu Peng (Zhejiang University, Hangzhou, China) for his help with CT scanning of these fossils. Our work also benefited from review comments of Thomas Martin (University of Bonn, Germany) and Nao Kusuhashi (Ehime University, Matsuyama, Japan), although any errors potentially remaining are our own. This work was supported by the National Natural Science Foundation of China (42161134003); Shandong Provincial Natural Science Foundation (ZR2017MD031); Program for Innovative Research Team of Excellent Talents in University of Shandong Province (2019KJH004); Taishan Scholar Program of Shandong Province (tsqn201812070).
References
Averianov, A.O. 2002. Early Cretaceous “symmetrodont” mammal Gobiotheriodon from Mongolia and the classification of “Symmetrodonta”. Acta Palaeontologica Polonica 47: 705–716.
Bi, S.-D., Zheng, X.-T., Meng, J., Wang, X.-L., Robinson, N., and Davis, B. 2016. A new symmetrodont mammal (Trechnotheria: Zhangheotheriidae) from the Early Cretaceous of China and trechnotherian character evolution. Scientific Reports 6: 26668. Crossref
Chen, M. and Luo, Z.-X. 2008. Morphology of dentition and postcranial skeleton of Akidolestes. Acta Geologica Sinica (Chinese Edition) 82: 155–164.
Cifelli, R.L. and Madsen, S.K. 1986. An Upper Cretaceous symmetrodont (Mammalia) from southern Utah. Journal of Vertebrate Paleontology 6: 258–263. Crossref
Cifelli, R.L. and Madsen, S.K. 1999. Spalacotheriid symmetrodonts (Mammalia) from the medial Cretaceous (upper Albian or lower Cenomanian) Mussentuchit local fauna, Cedar Mountain Formation, Utah, USA. Geodiversitas 21: 167–214.
Clemens, W.A. 1963. Late Jurassic mammalian fossils in the Sedgwick Museum, Cambridge. Palaeontology 6: 373–377.
Crompton, A.W. 1971. The origin of the tribosphenic molar. In: D.M. Kermack and K.A. Kermack (eds.), Early Mammals. Zoological Journal of the Linnean Society 50 (Supplement 1): 65–87.
Crompton, A.W. 1974. The dentition and relationships of the southern African Triassic mammals, Erythrotherium parringtoni and Megazostrodon rudnerae. Bulletin of the British Museum (Natural History), Geology 24: 397–437.
Ensom, P. and Sigogneau-Russell, D. 2000. New symmetrodonts (Mammalia, Theria) from the Purbeck Limestone Group, Lower Cretaceous of southern England. Cretaceous Research 21: 767–779. Crossref
Fox, R.C. 1976. Additions to the mammalian local fauna from the Upper Milk River Formation (Upper Cretaceous), Alberta. Canadian Journal of Earth Sciences 13: 1105–1118. Crossref
Gill, P.G. 2004a. A new symmetrodont from the Early Cretaceous of England. Journal of Vertebrate Paleontology 24: 748–752. Crossref
Gill, P.G. 2004b. Kuehneotherium from the Mesozoic Fissure Fillings of South Wales. 286 pp. Ph.D. Dissertation, University of Bristol, Bristol.
Han, G. and Meng, J. 2016. A new spalacolestine mammal from the Early Cretaceous Jehol Biota and implications for the morphology, phylogeny, and palaeobiology of Laurasian “symmetrodontans”. Zoological Journal of the Linnean Society 178: 343–380. Crossref
Hu, Y.-M., Wang, Y.-Q., Luo, Z.-X., and Li, C.-K. 1997. A new symmetrodont mammal from China and its implications for mammalian evolution. Nature 390: 137–142. Crossref
Hu, Y.-M., Wang, Y.-Q., Li, C.-K., and Luo, Z.-X. 1998. Morphology of dentition and forelimb of Zhangheotherium. Vertebrata PalAsiatica 36: 102–125.
Hu, Y.-M., Fox, R.C., Wang, Y.-Q., and Li, C.-K. 2005. A new spalacotheriid symmetrodont from the Early Cretaceous of northeastern China. American Museum Novitates 3475: 1–20. Crossref
Jäger, K.R.K., Gill, P.G., Corfe, I., and Martin, T. 2019. 3D occlusion and dental function of Morganucodon and Megazostrodon. Journal of Vertebrate Paleontology 39: e1635135. Crossref
Ji, Q., Luo, Z.-X., Zhang, X.-L., Yuan, C.-X., and Xu, L. 2009. Evolutionary development of the middle ear in Mesozoic therian mammals. Science 326: 278–281. Crossref
Kielan-Jaworowska, Z., Cifelli, R.L., and Luo, Z.-X. 2004. Mammals from the Age of Dinosaurs. Origins, Evolution, and Structure. 630 pp. Columbia University Press, New York. Crossref
Li, G. and Luo, Z.-X. 2006. A Cretaceous symmetrodont therian with some monotreme-like postcranial features. Nature 439: 195–200. Crossref
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Vol. 1: Regnum animale. Editio decima, reformata. 824 pp. Laurentii Salvii, Stockholm. Crossref
Lopatin, A.V., Maschenko, E.N., Averianov, A.O., Rezvyi, A.S., Skutschas, P.P., and Leshchinskiy, S.V. 2005. Early Cretaceous mammals of western Siberia: 1. Tinodontidae. Paleontological Journal 39: 523–534.
Lopatin, A.V., Averianov, A.O., Maschenko, E.N., and Leshchinskiy, S.V. 2010. Early Cretaceous mammals of western Siberia: 3. Zhangheotheriidae. Paleontological Journal 44: 573–583. Crossref
Luo, Z.-X. 1994. Sister taxon relationships of mammals and the transformation of the diagnostic mammalian characters. In: N.C. Fraser and H.-D. Sues (eds.), The Shadow of Dinosaurs: Early Mesozoic Tetrapods, 98–128. Cambridge University Press, Cambridge.
Luo, Z.-X. and Ji, Q. 2005. New study on dental and skeletal features of the Cretaceous “Symmetrodontan” mammal Zhangheotherium. Journal of Mammalian Evolution 12: 337–357. Crossref
Luo, Z.-X., Kielan-Jaworowska, Z., and Cifelli, R.L. 2002. In quest for a phylogeny of Mesozoic mammals. Acta Palaeontologica Polonica 47: 1–78.
Luo, Z.-X., Kielan-Jaworowska, Z., and Cifelli, R.L. 2004. Evolution of dental replacement in mammals. Bulletin of Carnegie Museum of Natural History 36: 159–175. Crossref
Luo, Z.-X. and Martin, T. 2023. Mandibular and dental characteristics of the Late Jurassic Mammal Henkelotherium guimarotae (Paurodontidae, Dryolestida). Paläontologische Zeitschrift 97: 569–619. Crossref
Mao, F.-Y., Hu, Y.-M., Li, C.-K., Wang, Y.-Q., Chase, M.H., Smith, A.K., and Meng, J. 2020. Integrated hearing and chewing modules decoupled in a Cretaceous stem therian mammal. Science 367: 305–308. Crossref
Mao, F.-Y., Liu C.-Y., and Meng, J. 2022. New material of the trechnotherian mammal Lactodens from the Early Cretaceous Jehol Biota: Comparison with Origolestes and implications for mammal evolution. Acta Palaeontologica Polonica 67: 135–153. Crossref
Martin, T. 2018. Mesozoic mammals—early mammalian diversity and ecomorphological adaptations. In: F.E. Zachos and R.J. Asher (eds.), Handbook of Zoology Mammalia, Mammalian Evolution, Diversity and Systematics, 199–299. De Gruyter, Berlin. Crossref
Martin, T., Averianov, A.O., Schultz, J.A., Schellhorn, R., and Schwermann, A.H. 2022. First spalacotheriid and dryolestid mammals from the Cretaceous of Germany. Acta Palaeontologica Polonica 67: 155–175. Crossref
Marsh, O.C. 1887. American Jurassic mammals. American Journal of Science 33: 326–348. Crossref
McKenna, M.C. 1975. Towards a phylogenetic classification of the Mammalia. In: W.P. Luckett and F.S. Szalay (eds.), Phylogeny of the Primates, 21–46. Plenum Press, New York. Crossref
Plogschties, T. and Martin, T. 2020. New information on the maxilla, dentary, and dentition of Maotherium sinense, with comments on the zhangheotheriid dental formulae. Paläontologische Zeitschrift 94: 155–165. Crossref
Rougier, G.W., Ji, Q., and Novacek, M.J. 2003. A new symmetrodont mammal with fur impressions from the Mesozoic of China. Acta Geologica Sinica‐English Edition 77: 7–14. Crossref
Sweetman, S.C. 2008. A spalacolestine spalacotheriid (Mammalia, Trechnotheria) from the Early Cretaceous (Barremian) of southern England and its bearing on spalacotheriid evolution. Palaeontology 51: 1367–1385. Crossref
Swofford, D.L. 1993. PAUP: Phylogenetic Analysis Using Parsimony, Version 4.0b10. Champaign, IL: Computer Program Distributed by the Illinos Natural History Survey.
Tsubamoto, T., Rougier, G.W., Isaji, S., Manabe, M., and Forasiepi, A.M. 2004. New Early Cretaceous spalacotheriid “symmetrodont” mammal from Japan. Acta Palaeontologica Polonica 49: 329–346.
Zhang, H.-G., Yu, D.-X., Feng, Y.-H., Pei, R., and Zhou, C.-F. 2022. A Lujiatun-like dinosaurian assemblage of the Jehol Biota from Ningcheng, Inner Mongolia, Northeast China. Acta Palaeontologica Polonica 67: 617–621. Crossref
Zhang, H.-G., Yin, Y.-L., Pei, R., and Zhou, C.-F. 2024. Early-diverging titanosauriform (Dinosauria, Sauropoda) teeth from the Lower Cretaceous Yixian Formation of southeastern Inner Mongolia, Northeast China. Acta Geologica Sinica-English Edition 98: 303–310. Crossref
Zhou, C.-F. Bhullar, B.-A.S., Neander, A.I., Martin, T., and Luo, Z.-X. 2019. New Jurassic mammaliaform sheds light on early evolution of mammal-like hyoid bones. Science 365: 276–279. Crossref
Acta Palaeontol. Pol. 69 (2): 315–327, 2024
https://doi.org/10.4202/app.01154.2024