

Hettangian, Early Jurassic coleoids from West Somerset, SW England—filling a gap in the coleoid record of NW Europe
DAVID H. EVANS, CHRISTIAN KLUG, ANDREW H. KING, and KEVIN N. PAGE
Evans, D.H., Klug, C., King, A.H., and Page, K.N. 2024. Hettangian, Early Jurassic coleoids from West Somerset, SW England—filling a gap in the coleoid record of NW Europe. Acta Palaeontologica Polonica 69 (3): 425–445.
The semi-articulated remains of two specimens of the putative diplobelid coleoid species of Clarkeiteuthis and a coleoid of uncertain affinity (possibly a phragmoteuthid) are described from a single bed in the Blue Lias Formation (early Hettangian, Planorbis Subchronozone) of the Somerset coast (United Kingdom). Remains of Hettangian, (Early Jurassic) coleoids are generally rare worldwide and consist largely of disassociated hard parts. Previously, the oldest recorded putative diplobelids (Clarkeiteuthis) were known from the Sinemurian of Dorset. The Somerset specimens extend their range into the earliest Jurassic, shortly after the end Triassic mass extinction. Combined with the coleoid of uncertain affinity and a previously described belemnitid, this assemblage is significant in the light of the Mesozoic radiation of coleoids. Several isolated coleoid ink sacs are also known from the same bed, suggesting that this unit may have some of the properties of a Lagerstätte. The relatively frequent occurrence of coleoids in this bed combined with a record of an ichthyosaur stomach containing numerous arm hooks attributable to Clarkeiteuthis (Planorbis Chronozone of Dorset) suggest that coleoids may have been relatively abundant and diverse during the earliest Jurassic.
Key words: Cephalopoda, Coleoidea, Diplobelida, Early Jurassic, Hettangian, Blue Lias Formation, Somerset, United Kingdom.
David H. Evans [devanscephs@gmail.com; ORCID: https://orcid.org/0000-0002-9157-4610 ], 10 Liddymore Road, Watchet, Somerset, TA23 0DQ UK.
Christian Klug [chklug@pim.uzh.ch; ORCID: https://orcid.org/0000-0002-4099-7453 ], Paläontologisches Institut und Museum, University of Zurich, Karl Schmid-Strasse 6, 8006 Zurich, Switzerland.
Andrew King [andy@geckoella.co.uk; ORCID: https://orcid.org/0000-0002-4231-9012 ], Geckoella Ltd, East Quay, Harbour Road, Watchet, TA23 0AQ UK.
Kevin Page [kevinnpage@gmail.com], Camborne School of Mines, University of Exeter, Penryn, Cornwall, TR10 9FE, UK.
Received 15 May 2024, accepted 27 July 2024, published online 27 September 2024.
Copyright © 2024 D.H. Evans et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
As with much of the rest of the world, by comparison with younger Jurassic horizons, the record of early Hettangian (Planorbis and Liasicus chronozones) coleoids in southwest England is extremely sparse. In Dorset, with the exception of arm hooks in the stomach contents of ichthyosaurs (Pollard 1968, 1990), there appear to be no coleoids recorded from horizons older from the upper part of the lower Sinemurian (Hart et al. 2019: 331). A similar situation pertains to much of northern Europe where Riegraf (1996: table 3.2, 3.3) summarised the then known Hettangian coleoid fauna as consisting of a few aulacoceratids, a phragmocone attributed to ?Phragmoteuthis Mojsisovics, 1882, phragmocones of unspecified affinity, coleoid arm hooks, and rare occurrences of belemnite rostra belonging to Schwegleria Riegraf, 1980. The latter came from the Liasicus Chronozone of Nürtingen, southwest Germany (Riegraf 1996), and were subsequently reported from Fontenoille in southeast Belgium (Delsate et al. 2002). Despite the discovery and description (Zhu and Bian 1984) of the belemnites Sinobelemnites Zhu and Bian, 1984, and Sichuanobelus Zhu and Bian, 1984, from the Carnian of Sichuan, southwest China, their age was questioned (Doyle 1993, 1994; Weiss and Delsate 2006) and it generally continued to be accepted that the Belemnitida originated as small forms in northern Europe during the earliest Jurassic and were restricted to that area until the Pliensbachian reflected their origin in that region (Weiss and Delsate 2005, 2006). Subsequently, Iba et al. (2012) showed that the Chinese taxa co-occurred with Carnian ammonites while Iba et al. (2012, 2014) described Hettangian and Sinemurian belemnite assemblages from Japan, the diversity of which indicated a pre-Jurassic history and a probable Triassic origin for the Belemnitida. Additional occurrences of Sinobelemnites and Sichuanobelus from the Carnian of Yunnan, southwest China were reported by Ma et al. (2023), while Niko and Ehiro (2022) described the sinobelemnitid Tohokubelus Niko & Ehiro, 2022, from the Lower Triassic (Olenekian) of Japan, suggesting a possible Permian origin for the Belemnitida (Stevens et al. 2023), and a pre-Jurassic history of residence in the eastern Tethys.
The apparent paucity of earliest Jurassic coleoid records constitutes a significant gap, particularly as it succeeds the end Triassic mass extinction event and represents an interval covering part the Mesozoic radiation of coleoids, considered to have been initiated during the Triassic and Early Jurassic (Tanner et al. 2017; López-Córdova et al. 2022).
That coleoids were present in the lowermost Jurassic of the West Somerset coast has been known since the early 19th century when Buckland (1836: pl. 29:10) figured a coleoid ink sac collected from Watchet (West Somerset) by Mary Anning. More recently, a coleoid conotheca consisting of part of the phragmocone and complete proostracum from the Planorbis Chronozone at nearby Blue Anchor was described by Klug and Fuchs (2010) who described it as “the earliest record of an undoubted belemnitid proostracum of clear stratigraphic affinity” (Klug and Fuchs 2010: 181), although they were concerned by the lack of a rostrum, suggesting that it was either minute or had been lost during the specimens stratinomy. Subsequently, one of the authors (KP) discovered a coleoid with a phragmocone, fragmentary proostracum, ink sac and partial arm crown preserved in the Planorbis Chronozone at Watchet (Page and Hart 2018), prompting further investigation of the Planorbis Chronozone of the West Somerset coast. To date, several ink sacs, as well as three specimens consisting of partial arm crowns, ink sac and other structures (Table 1) have been recovered.
These three specimens, described in detail below, consist of a coleoid of uncertain affinity, although possibly a phragmoteuthid, and two individuals belonging to the putative diplobelid Clarkeiteuthis Fuchs et al., 2013. Together with probable Clarkeiteuthis arm hooks that were misattributed to Chondroteuthis Bode, 1933, by Dick et al. (2016: 427) in the stomach contents of an ichthyosaur from the Planorbis Chronozone of Lyme Regis (Pollard 1968: text-fig. 2a–c, 1990), these specimens are the oldest known examples of Clarkeiteuthis, and as such, the oldest (putative) members of the order Diplobelida worldwide. Moreover, these fossils may represent the oldest body-fossils of total-group decabranchians in general and highlights the significance of this assemblage. To this, it may be added that the specimen described below as a coleoid of uncertain affinities, differs from the specimen described by Klug and Fuchs (2010), and thus hints at a considerable diversity in this assemblage, suggesting that this horizon may yield additional material to future searches.
Institutional abbreviations.—PIMUZ, Paläontologisches Institut und Museum der Universität Zürich; LYMPH, Lyme Regis Phillpot Museum, Lyme Regis, UK; OUMNH, Oxford University Museum of Natural History, UK; BRMSG, Bristol Museums and Art Gallery, UK.
Geological setting
Description.—The Hettangian and lower Sinemurian (Lower Jurassic) successions of the southern margin of the eastern part of the Bristol Channel Basin are visible in many of the extensive cliff and foreshore exposures of the Blue Lias and Charmouth Mudstone formations on the West Somerset coast (Fig. 1). The lithological succession was recorded in detail (Palmer 1972; Whitaker and Green 1983; Warrington and Ivimey-Cook 1995; and reviewed by Simms 2004) and is supported by a high-resolution ammonite biochronology using biohorizons (Page 2002, 2003, 2005; Weedon et al. 2017, 2018; see Page 2017 for discussion of principles). The Blue Lias Formation comprises cyclic alternations of limestones with light and dark marls and laminated, organic rich mudstones, interpreted as of hemipelagic origin (Weedon 1986) and strongly influenced by sea-level and climate change (Weedon et al. 2018). In West Somerset, at a thickness of 170 m, the Blue Lias Formation is substantially thicker than on the Dorset/Devon coast, reflecting the greater rate of subsidence of the Bristol Channel Basin during the Hettangian and early Sinemurian (Simms 2004).
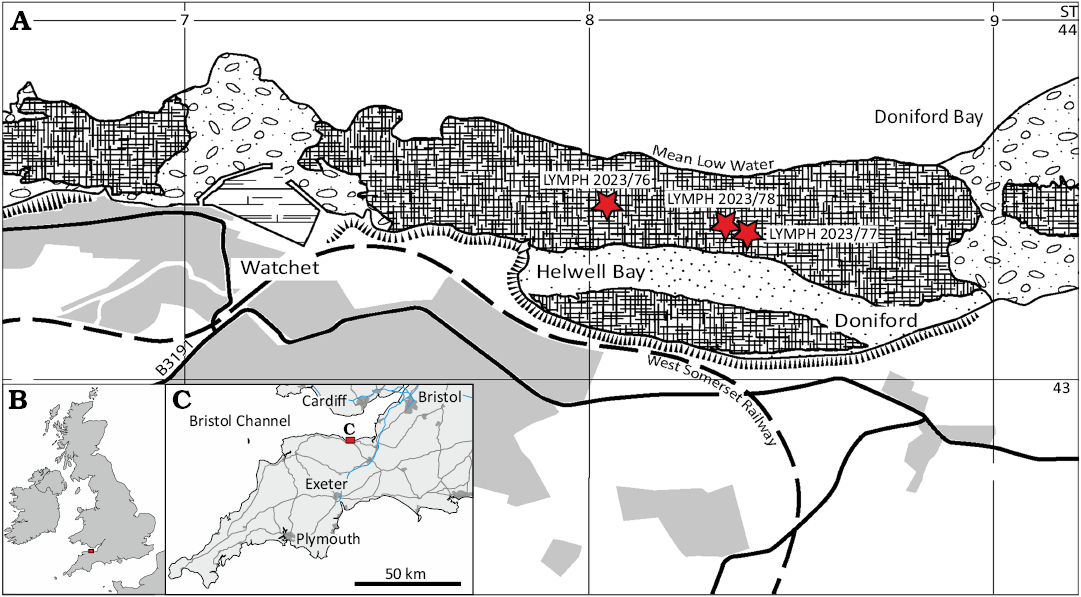
Fig. 1. Maps of Britain, Ireland, and north-western France (B) and south-west England (C) showing location of the study area near Watchet (red rectangle). Map of coast at Watchet (A) showing locations of the coleoids (stars) mentioned in the text.
On the West Somerset coast, the Planorbis Subchronzone (Planorbis Chronozone) is about 4 m thick (Page and Bloos 1998: fig. 2; Weedon et al. 2017, 2018; Fig. 2 herein). Although in general character its composition resembles typical Blue Lias Formation at higher levels, it includes several relatively indurated laminated mudstone horizons which weather to a pale grey-blue colour and yield ammonites with the aragonitic shell preserved. The general succession of beds recognisable only varies in minor detail along the coast (e.g., Blue Anchor-Watchet-St Audries Bay; see Page and Bloos 1998: fig. 2) although the quality of aragonite preservation does.
The coleoids were recovered from the basal part of Bed 24 (Whitaker and Green 1983: 65; Fig. 2 herein). Bed 23, underlying Bed 24, is a dark marl exhibiting relict lamination, largely disturbed, and obscured by burrows and more homogenous bioturbation. Except where disturbed by burrowing, fine broken shell debris rests parallel to the laminations. The finer-grained fragmented shell debris consists of moulds after aragonitic shell and a calcitic fraction composed of echinoderm stereom and echinoid spines. Coarser, more intact shell belonging to crushed Psiloceras ammonites, as well as bivalve shells and echinoid spines are dispersed through the sediment, and also as matrix supported concentrations. Although crushed, sections through Psiloceras conchs contain matrix and are not flattened to the degree seen in the laminated mudstones. Samples of Bed 23 effervesced in 10% HCl and disintegrated rapidly, leaving a residue of mud after several hours of dissolution.
The lower half of Bed 24 consists of about 1 m of laminated mudstones and is overlain by approximately 1 m of calcareous mudstones resting on the laminated mudstones. The former are lighter in colour than the laminated mudstones of the overlying Liasicus Chronozone, and although dark grey when fresh, weather to a pale grey blue. Close inspection of freshly broken sections through the mudstone near the base of Bed 24 exhibit laminae 0.05 mm thick, some of which appear to form bundles of 0.75–1.0 mm thick. Although dominated by clay minerals and probable organic material, many pale flecks and blebs interweaved into the laminae are most likely attributable to microspar after the neomophism of coccolith debris (see Weedon et al. 2018: 16, fig. 7). Flakes of the laminated mudstone effervesce gently in 10% HCl for at least 48 hours without disaggregation but lose their strength once decalcified.
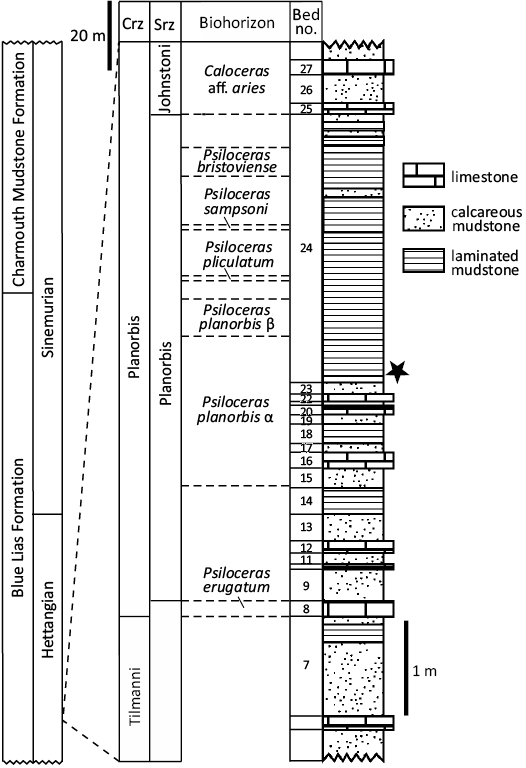
Fig. 2. Stratigraphy of the basal Blue Lias Formation in Doniford Bay showing the horizon from which the coleoids were recovered (star). Stratigraphic logs after Page and Bloos (1998: fig. 2), KPN (own data from 2022) and Simms et al. (2004). Abbreviations: Crz, Chronozone; Srz, Subchronozone.
Bedding plane surfaces of Bed 24 may be nearly devoid of macrofossils or strewn with flattened Psiloceras conchs ranging from 3–4 mm to 80 mm in diameter. Disarticulated bivalves, including the spat and juvenile shells of Liostrea, Pteria?, and Cuneigervillia? may also be present in large numbers, as are isolated echinoid spines. Angular fragments of broken shell also occur and may be abundant on some bedding planes. While black, organic debris occurs throughout Bed 24 it appears to be particularly abundant near its base and consists largely of wood fragments, conifer shoots, fish scales, and less frequently, coleoid ink sacs.
Although some bedding plane surfaces are devoid of shelly material, and the mean density of ammonites on other surfaces is estimated at about 50 per m2, some surfaces exhibit ammonites crowded together along a more or less straight line; similar such concentrations are known from the German Posidonia Shales (Seilacher 1982a, b). In such concentrations, the conchs both overlap and rest on each other, and their estimated density within the linear zone is approximately 1600 per m2. Abundant comminuted shell debris is also associated with these concentrations and where visible in cross section, three or four tiers of flattened conchs may be seen to be resting on each other. In cross section, the clusters have clearly defined margins. The clearly defined margins and linearity of these concentrations indicates that they are gutter fills.
Interpretation.—Weedon (1986), concluded that the Blue Lias Formation of southern England accumulated as hemipelagic sediment on an epicontinental shelf under a regime of climate forced cyclic sedimentation—and this model is fully updated and expanded by Weedon et al. (2017). The succession contains numerous hiatuses evidenced by the truncation of burrows at the tops of units, the presence of exotic sediment in burrows, the encrustation of shells by postmortem epicoles (Seilacher 1982a, b; Davis et al. 1999; Klug and Korn 2001), condensation of shells and shell debris through winnowing and scouring as well as aligned bands of shells interpreted as “gutter fills” (Weedon et al. 2018). Comparison of successions using Shaw plots (Weedon et al. 2017, 2018) facilitated the location of major changes in relative sedimentation rates while the utilisation of a highly resolved ammonite biostratigraphy (Weedon et al. 2018) further constrained the location and duration of hiatuses. One such hiatus, of an 0.3 Ma duration is present in Bed 24 (Weedon et al. 2018: fig. 14), approximately a meter above its base and above the horizons from which the coleoids were collected.
The presence in Bed 24 of numerous horizons containing comminuted and concentrated shell including concentrations of ammonites in gutter fills indicates relatively frequent winnowing and scouring by currents below fair-weather wave base most probably generated by storms (see also Seilacher 1982a, b). Evidence for the survival and continued growth of serpulids on ammonite conchs after the death of the ammonite and/or postmortem colonisation of empty conchs suggest that the conchs may have rested on the substrate undisturbed and largely free of cloaking sediment at intervals. Where there is evidence of extensive and heavy serpulid colonisation on both flanks, the empty conchs may have been turned over several times and/or transported from another location. Small articulated cidaroid echinoids (Diademopsis) have been recorded from Bed 18 and Bed 24 (Page and Paul 2017) in Doniford Bay, and Bed 24 at Lilstock (DHE personal observation 2024). Page and Paul (2017) considered that their preservation in an articulated state was likely to have arisen from rapid burial in fine sediment falling out of suspension from waning bottom currents; They also noted that the ammonite conchs could have supplied a substrate from which algae were grazed by the echinoids.
The laminated mudstone facies of the Planorbis Chronozone appear to differ from those of the upper Hettangian and lower Sinemurian in the apparently greater frequency of winnowed and scoured horizons. This may reflect the lower rates of pre-Liasicus Chronozone subsidence in the Bristol Channel Basin, leading to relatively frequent substrate disturbance prior to sufficiently deep burial to be isolated from disturbance by activity on the substrate.
The sedimentary environment represented by Bed 24 appears to reflect a regime at times dominated by the winnowing, scouring, and reworking of fines, with the concurrent concentration of coarser sediment (predominantly shell) as condensates and current transported accumulations. Fallout of fines from storm suspended sediment may have generated rapidly deposited blankets of carbonate mud largely devoid of coarser clasts, but some of this sediment probably represents a relatively steady background sedimentation. While there is some evidence of bioturbation, evidence of taxa representative of infaunal niches is sparse, indicating that the substrate, if not the sea floor, was frequently stressed and probably oxygen deficient, as may also be indicated by the low diversity of the epifauna. It is clear that times the sea floor was sufficiently firm to support the movement of cidaroid echinoids.
Material and methods
All the coleoid specimens, coming from intertidal exposures, were saturated with seawater. Prior to drying the specimens, they were left to desalinate in freshwater, replenished every week over a period of eight weeks. To circumvent the risk of delamination, once desalinated the slabs were left to dry slowly in a cool environment for two weeks. The specimens were then inspected to determine what preparation, if any, could be carried without risk of damage. Due to the relative hardness of the matrix, the brittleness of the remaining calcareous skeleton, and the softness of the sclerotized parts, it was decided to describe the specimens without attempting any prior mechanical preparation as the risk of serious damage resulting from such work was too high.
Values for the dimensions and characters of the phragmocones (Table 2) were taken from drawings (Klug and Fuchs 2010: fig. 2) of PIMUZ 27078 and the specimen described by Hölder (1973). Values for LYMPH 2023/77 were taken directly from the specimen. All three specimens are flattened, and brittle deformation is assumed. In all three specimens one lateral side of the phragmocone is missing and has been reconstructed by Klug and Fuchs (2010: fig. 2). Consequently, values for the diameter, length and apical angle represent minimums and that for cameral depth, a maximum. Formulae used to recover these values (Fig. 3) assume that the phragmocone cross-section was originally circular, or nearly so.
Study of the specimens was carried out under daylight and a GXM Zoom trinocular microscope at magnifications ranging from × 8 to × 50 and illuminated from a LED source. Specimens were photographed using a Canon EOS1300D equipped with a Canon AF 35 mm lens and a Canon EF 100 mm macro lens illuminated by a LED source. Microscope images were captured with a GXCAM-U3-18 microscope camera and processed using GXCaptureT software. Further processing and editing was carried out in Adobe Photoshop.

Fig. 3. Diagrams illustrating methods used to obtain values of the phragmocone diameter, length and apical angle for PIMUZ 27078 and the specimen described by Hölder 1973 (A) and LYMPH 2023/77 (B). The methods assume brittle deformation and originally circular phragmocone cross-section. See text for discussion.
All the locations referred to on the West Somerset coast lie within the Blue Anchor to Lilstock Coast Site of Special Scientific Interest (SSSI) and are formally protected under national conservation legislation. Much of the SSSI is privately owned. All sampling and collecting from in-situ exposure, requires both a permission form the landowner and may also require consent from the National Conservation Agency, Natural England (https://designatedsites.naturalengland.org.uk).
Preservation of coleoids.—All currently known coleoid remains from the Planorbis Subzone at Blue Anchor and Watchet are listed in Table 1. The precise horizon from which the Blue Anchor specimens originated is uncertain as LYMPH 2023/83 came from a fallen block and it seem likely that PIMUZ 27078 did also. All the ink sacs and more complete coleoids from Watchet came from the basal 200 mm of Bed 24 which is extensively exposed on the foreshore of Helwell and Doniford Bay. The precise location from which OUMNH PAL.J.003564, collected by Mary Anning, originated is unknown but the colour of the matrix and the presence of crushed smooth conchs Psiloceras indicates a likely origin in Bed 24. Their stratigraphical distribution appears to be limited. Despite searching other horizons within the Planorbis Subzone, no other remains of coleoids have been observed other than in Bed 24. Their geographically limited occurrence within Doniford and Helwell Bay may reflect local conditions at the time of deposition, but perhaps more likely, reflects the interaction of the varying wave climate in Doniford Bay with the changing dip and strike of Bed 24. This interaction limits the likelihood of discovering new fossil material to areas where erosion and scouring are sufficiently high to regularly refresh exposure, but not so rapid as to destroy exposed specimens over short intervals, permitting their discovery before being lost to erosion. The effects of scouring are particularly evident in LYMPH 2023/76 where the surface of the arm hooks are striated and planed down to the extent that the pseudopulp cavity is partially exposed in some of them.
Table 1. Coleoid specimens currently known from the Planorbis Subchronozone at Blue Anchor, Helwell Bay, and Watchet tabulating organs and skeletal elements preserved.
|
Locality |
Specimen No |
Arm hooks |
Jaws |
Proostracum |
Ink sac |
Mantle |
Phragmocone |
Rostrum |
|
Blue Anchor |
PIMUZ 27078 |
– |
– |
× |
× |
? |
× |
– |
|
Blue Anchor |
LYMPH 2023/83 |
– |
– |
× |
× |
– |
– |
– |
|
Helwell Bay |
LYMPH 2023/77 |
× |
? |
× |
× |
– |
× |
– |
|
Helwell Bay |
LYMPH 2023/76 |
× |
– |
× |
× |
– |
× |
– |
|
Helwell Bay |
LYMPH 2023/78 |
× |
? |
– |
× |
× |
– |
– |
|
Helwell Bay |
LYMPH 2023/79 |
– |
– |
– |
× |
– |
× |
– |
|
Helwell Bay |
LYMPH 2023/80 |
– |
– |
– |
× |
– |
– |
– |
|
Helwell Bay |
LYMPH 2023/81 |
– |
– |
– |
× |
– |
– |
– |
|
Helwell Bay |
LYMPH 2023/82 |
– |
– |
– |
× |
– |
– |
– |
|
Watchet |
OUMNH PAL-J.003564 |
– |
– |
– |
× |
– |
– |
– |
Table 2. Estimated dimensions and ratios of structures in PIMUZ 27078, LYMPH 2023/77, and Hölder’s (1973) specimen.. Measurements taken directly from LYMPH 2023/77 and Klug and Fuchs (2010: fig. 1). All values assume brittle deformation. *As the proostracum is largely missing in LYMPH 2023/77, the distance between the posterior of the brachial crown and the anterior of the phragmocone is taken as a substitute.
|
Specimen |
PIMUZ 27078 |
LYMPH 2023/77 |
|
|
Restored anterior diameter of phragmocone (mm) |
9.8 |
>17.4 |
6.68 |
|
Estimated length of phragmocone |
28.7 |
>31.0 |
19.1 |
|
Restored apical angle of phragmocone (degrees) |
18.5 |
31 |
19.1 |
|
Depth of camerae |
0.13 |
0.06 |
0.12 |
|
Length of proostracum (mm) |
50 |
69* |
34 |
With the exception of LYMPH 2023/78, in which mantle muscle is unequivocally present, the only soft tissue otherwise preserved consists of ink sacs and ink ducts. Of the coleoid remains currently known from this horizon, six (LYMPH 2023/79–83, OUMNH PAL-J.003564) consist of isolated ink sacs and three (LYMPH 2023/76–78) comprise of calcified (phragmocone) and sclerotised (arm hooks, jaws, proostracum) remains. The organisation of the remains of LYMPH 2023/76–78 suggests that although the soft tissue decayed completely in LYMPH 2023/76 and 77, other than sediment infiltration and the collapse of structures supported by soft tissue, very little disturbance or displacement of the body took place during the decay process. The lack of evidence for disturbance in these three specimens suggests that there were few, if any, scavengers either on or within the substrate, implying probable anoxic/dysoxic conditions inimical to their presence.
Although the brachial crowns as represented by the distribution of the arm hooks indicates that they are incomplete, the arm hooks present such consistent orientations and aspects to suggest that they are relatively undisturbed, particularly in the case of LYMPH 2023/77 (Fig. 4). This is less clear in LYMPH 2023/76, as the current state of preservation of many of the hooks is too fragmentary to perceive any order in their distribution, although there are hooks that are consistently paired and ordered and were clearly attached to the same arm. The clustering and overlapping of hooks in LYMPH 2023/77 (Fig. 5: cluster 1, rows a and b) could reflect a contraction of some of the arms or a denser distribution of hooks along the arms. Alternatively, this could have resulted from some of the arms resting at oblique angles to the substrate. If this were the case, the arms would have been buried and supported by sediment. Reduction in sediment thickness through compression and dewatering reduced the sediment thickness to millimetres, bringing hooks into close contact and overlapping each other. This also implies that the moribund animal or carcass was buried rapidly in sediment.

Fig. 4. Coleoidea indet. LYMPH 2023/77 from the Early Jurassic, Hettangian Stage of Doniford Bay, Watchet, UK. A1, image of whole specimen; A2, interpretative sketch mapping out the distribution of the surviving components of the organism; A3, broken edge of phragmocone wall overlain by two sets of lamellae (edges marked by white arrowheads) originating from the phragmocone wall and representing the remnants of probable septa with a thin layer of cameral deposit, edge of ink sac to lower left (black arrowhead); A4, margin of phragmocone with outer, iridescent layer and fine striations (arrowhead) possibly representing fragment of proostracum. A5, iridescent layer showing longitudinal striae (black arrowhead) outside phragmocone wall (proostracum) and partially overlain by smooth shell (white arrowhead) that may represent a fragment of the outer shell layer of the conotheca, primordial rostrum, or an anterior part of the orthorostrum; A6, phragmocone wall underlying ink sac with probable septa (arrowheads) (see A3) and showing sutures and striae representing probable growth increments; A7, cluster of closely packed arm hooks. A8, dark mass possibly representing jaw (see Fig. 8 for interpretation). A9, type A hooks with spurs (arrowheads), other hooks mainly type B; A10, enlargement of brachial crown showing rows and pairs of hooks; A11, type B hooks with strongly curved terminations of the uncinus.
The posterior part of the phragmocone is missing in all three specimens while LYMPH 2023/76 and 78 have some leaked ink within the mantle cavity. In discussing the factors controlling soft tissue preservation, Reitner and Urlichs (1983) pointed out that in order for this type of preservation to occur at all, the cephalopod carcass must have reached the sea floor before the decay process became advanced. They observed (Reitner and Urlichs 1983: 460) that in the case of belemnitids, damage to the phragmocone would have rendered the organism negatively buoyant (an example is given in Klug et al. 2021); they further noted that damage in specimens from the Posidonienschiefer, while attributed to compaction, might also be caused by predation as evidenced by specimens where breakage had been attributed to bites (Riegraf and Reitner 1979: 301). Breakage caused by a bite was noted by Reitner (2009) who observed that this may also have resulted in ink leaking into the mantle cavity. Similar damage was reported by Schweigert (2018). However, during decay experiments on squids and octopus carried out by Clements et al. (2017), the rupturing of the ink sac was observed to occur during the decay process. As a consequence, the presence of leaked ink, as evidence of predation is equivocal since it may also be attributed to the postmortem decay of the ink sac. Nevertheless, the loss of the apical portion of the phragmocone and the rupturing of the ink sac, particularly in LYMPH 2023/76 and 78 might be considered consistent with attempted predation, leaving the lethally injured prey to sink to the sea floor (a pabulite sensu Klug et al. 2021).
Other than the ink sacs and ink ducts present in all three specimens, only LYMPH 2023/78 has soft tissue preserved in the form of mantle muscle. Many recent decabrachian coleoids use ammonium to regulate their buoyancy, the presence of which during tissue decay may inhibit the pH from declining to a level below the carbonic acid dissociation constant for any great length of time, thus inhibiting and/or limiting calcium phosphate precipitation (Clements et al. 2017; Fuchs 2023). Some modern decabrachians (Sepia) possess a phragmocone and do not have significant levels of ammonium in their tissues (Denton 1974). This seems likely to also have been the case in phragmocone-bearing fossil decabrachians. Therefore, it may be no coincidence that in LYMPH 2023/78, where muscle tissue is preserved, the phragmocone appears to be entirely missing and could have been lost through dissolution under low pH conditions.
The isolated ink sacs (LYMPH 2023/80–82) from Helwell Bay all appear to be partially eroded. Only one (LYMPH 2023/80) preserves an ink sac and part of the ink duct. LYMPH 2023/81 and LYMPH 2023/82 are either ink ducts or extremely worn and elongate ink sacs with ducts that taper gently from the sac. Both the worn condition and isolated occurrences of the ink sacs suggest that they may have been prefossilised and reworked.
Systematic palaeontology
Subclass Coleoidea Bather, 1888
Order, family, genus and species uncertain
Coleoidea indet.
Figs. 4, 5.
Material.—Single specimen (LYMPH 2023/77) from the base of Bed 24 of the Planorbis Subchronozone, Planorbis Chronozone, Hettangian Stage, “Psiloceras Beds” of Blue Lias Formation; Foreshore of Doniford Bay [ST 0842 4337], Watchet, Somerset, UK.
Description.—General features: The length of the specimen, from the most distal arm hooks to the posterior of the remaining portion of the phragmocone is 115 mm. With the exception of the ink sac, the specimen is entirely flattened, and the phragmocone wall and septa heavily fractured. There is nothing preserved between the anterior of the ink sac and the dark mass to the posterior of the arm crown, a distance of 60 mm.
Arm hooks and arms: Many of the posterior arm hooks and the dark mass are partially buried in and obscured by a thin layer of finely crystalline material, interpreted as comminuted shell debris. The hooks range 0.6–1.4 mm in length and comprise two general types that may intergrade:
(i) type A: nearly straight, with a slender, faintly curved, short uncinus; a prominent spur is visible on some of the hooks (Figs. 4A8, 6A1) and where present, appears to be consistently located at a position 0.75% of the distance from the proximal end of the hook (Fig. 7). There are also several isolated spurs in the matrix which may have broken away from the hooks. The longest hook of this type is 1.17 mm and the shortest 0.56 mm. The proximal and distal angles (see Fuchs and Hoffmann 2017) measured on one hook are 10° and 36°, respectively;
(ii) type B: larger hooks (up to 1.7 mm), possessing a weakly curved shaft and a long, gently curved uncinus with a short, strongly curved distal end bearing a marked resemblance to a crochet hook (Figs. 4A11, 6A2, A3); although some of the hooks of this type appear to carry spurs, these are spurious and result from the compression of the shaft of one hook onto the uncinus of an underlying hook, creating a projection from the shaft of the upper hook that can be mistaken for a spur (Fig. 4A11); all the hooks of type B lack spurs; proximal and distal angles measured on one hook are 19° and 26°, respectively.
The wear caused by exposure to wave activity has resulted in the loss of the orbicular scar in all the hooks that are visible. Consequently, it is not possible to obtain proportionate measurements of the relative lengths of the uncinus, shaft and base.
The majority of hooks form three clusters anterior to the dark mass that might represent the buccal mass with the jaws. Each cluster consists of closely packed hooks that are often in contact and/or overlap with each other. Anterior to the clusters, discrete rows of isolated hooks extend forward (Fig. 5: d–h). Both the rows and clusters show consistent alignments and packing order, indicating that they are unlikely to be the product of minor transportation of the hooks, but largely reflect an original configuration; this is not surprising because cephalopod arms have the ability to extremely extend and contract. With few exceptions, the distal ends of the hooks face anteriorly, and the alignment of many of the hooks within the clusters suggests that the hooks form discrete rows. Potentially, nine rows of hooks (Fig. 5: a–i) may be discerned.

Fig. 5. Coleoidea indet. LYMPH 2023/77 from the Early Jurassic, Hettangian Stage of Doniford Bay, Watchet, UK. Sketch of distribution of arm hooks and interpretation of hook pairing and likely traces of arms. 1–3, clusters of arm hooks referred to in the text; a–i, paths of arm pairs as interpreted.

Fig. 6. Coleoidea indet. LYMPH 2023/77. A1, type A hook in lateral view drawn from an image showing the location of spur close to the distal end of the shaft, the most distal part of the uncinus is missing; A2, lateral view of apparently complete type B hook drawn from an image; A3, oblique view of type B hook showing a flatter “platform” zone on the inner surface of the shaft, base appears to be worn. Locations of base, shaft and uncinus marked, but exact lengths are estimates as the orbicular scar has been worn away in all examples.
The distal ends of the hooks face the anterior in all rows. In rows d–g the bases of the hooks face towards the axis of the brachial crown. In row h, the bases of the hooks, although also facing toward the axis of the crown, are reversed by comparison with rows d–g. This may be because the arm was torted, or it lay on the opposite side of the axis of the brachial crown, i.e., opposite to the other arms.
Hooks of type A appear to be in the minority and are associated with rows f, g and h. They also appear to be present on the left side of cluster 3, which is otherwise dominated by type B hooks. Cluster 1 consists almost entirely of large type B hooks, all of which lack spurs. The hooks are so closely packed that it is possible that spurs, if present, have been obscured from view. Cluster 2 contains a mixture of types A and B, including some that may be intermediate in form. With the exception of row h, cluster 3 also consists of type B hooks and intermediate forms. Hooks of both types appear to become progressively shorter and increasingly slender toward the proximal ends of the arms.
Each of the rows a–i probably represents an arm, but rows d–e and f–g are likely to represent pairs of hooks on the same arm. At a minimum, cluster 1 may represent parts of two arms, cluster 2, parts of three arms, and cluster 3, parts of two arms. Thus, the remains of up to six arms may be preserved. It is unclear whether the other arms were weathered away, remain still covered by sediment, or were detached and fell off during the decay of soft tissue and are represented by the more disordered portions of cluster 1 in particular.
Dark mass: The boundaries of this structure (Fig. 4A8) are largely obscured by the same layer of shell debris that cloaks some of the posterior arm hooks (Fig. 4A10). The structure is almost entirely flattened. The main body of the structure is comprised of material the same colour as that of the arm hooks, with a similarly reflective surface, although markedly pitted by comparison. This area is thought to be relatively thick. The margins of the structure, although appearing to comprise a similar material are generally less reflective and have the appearance of being thinner. Parts of both the core and margins are missing in several places and have either been broken off prior to burial or lost to recent erosion.
The core of the structure is 1.7 mm long and 1.5 mm wide. The main feature is a slender projection aligned with the long axis and tapering away from the main body. At the opposite end, a low ridge curves convex outwards symmetrically around the long axis in the plane in which the structure is flattened. The main ridge is associated with two striae or very low ridges parallel with and further outboard of the main ridge. Laterally, the main body extends out to form a pair of broad areas, the margins of which are not visible, but may be up to 0.6 mm long.
The projection is divided anteriorly by a fissure 0.1 mm wide and roughly aligned to the long axis of the structure. The fissure tapers and closes over a distance of 0.55 mm, but a shallow groove, in places filled with shell debris persists and deviates obliquely from the long axis for the length of the body.
Ink sac: The ink sac is 13.2 mm long by 7.7 mm wide and partially flattened. The remaining 4 mm of an ink duct extends from the anterior end of the ink sac. The surface of the ink sac is badly worn though wave erosion, but a number of depressed lobes and shallow groove may indicate that the surface of the sack had been folded prior to or during burial. The posterior end of the ink sac overlaps with the anterior part of the phragmocone by at least 6 mm. The surface of the ink sac is coated in places with calcite or aragonite that has a sucrose texture. These patches may represent the remains of crushed septa.
Conotheca: Nothing remains of the proostracum anterior of the ink sac, and where shell survives, as a consequence of the flattening and crushing of the conch, it is difficult to distinguish between potential proostracum, phragmocone wall or septa. The conotheca is heavily fractured, and matrix is present within the fractures so that there is continuity between the external and internal matrix. This suggests that fracturing probably took place during early burial while the sediment remained relatively mobile. The resultant jigsaw of overlapping fragments creates an impediment to interpretation of the structure of the conotheca.
Underlying and partially obscured by the ink sac, part of the inner surface of the phragmocone wall is visible around the perimeter of the ink sac. Here, the presence of sutures is marked by narrow, shallow grooves that are on average, 1.1 mm apart (Fig. 4A6). Where seen, the sutures are straight. Between the sutures, the inner surface of the phragmocone wall exhibits faint striae about 0.08 mm apart and parallel with the sutures. These are interpreted as growth increments.
On the inner side of the phragmocone wall, and in places extending onto the margins of the ink sac, two lamellae are visible, each of which is divisible into two fabrics. These are (i) an off-white, finely sucrose-textured fabric, and (ii) a denser, multi-layered fabric that has a nacreous appearance, the edges of which are indicated by arrows in Fig. 4A3. Fabric 1 coats both sides of fabric 2 but there appear to be traces of matrix between adjacent layers of fabric 1, suggesting the original presence of cavities between the laminae. The most anterior of the lamina composed of fabric 2 appears to be attached to the phragmocone wall at its anterior margin, as may also the second more posterior lamina. The distance between the contacts of the two laminae where they are in contact with the phragmocone wall is similar to the distance between the sutures, although it cannot be demonstrated that that they are inserted at the sutures. These structures are interpreted as the remains of two septa; fabric 2 may represent the septum and fabric 1, as questionable cameral deposits (see Bandel and Spaeth 1988; Doguzhaeva et al. 2003: fig. 9.10).
On the outer surface of the phragmocone wall there is a very thin layer of laminated, strongly iridescent material. Parts of the edge of the crushed conotheca can be traced along one side of the specimen although the opposing margin was lost during the extraction of the specimen. The most anterior segment of the conotheca margin appears to have been fractured longitudinally, and the two edges of the fracture are imbricated against each other. The upper facing surface of the conotheca is composed of a material of similar thickness and general fabric to that forming the outer surface of the phragmocone as seen around the ink sac and probably represents the same structure. The iridescent outer surface of the conotheca is covered in traces of striae 0.07 mm apart (Fig. 4A4) and oriented to face obliquely apically away from the mid-line of the conotheca.
The most posterior portion of the conotheca that is preserved comprises an isolated area 10 mm to the posterior of the rest of the remaining conotheca. This portion is composed of several layers of crushed shell consisting of a chaotic mix of septal and phragmocone wall fragments. The presence of matrix between the conch fragments and in continuity with the matrix external to the conotheca indicates that the walls were ruptured during burial while the sediment remained unconsolidated.
Although this part of the conotheca is so fragmented that it is difficult to identify any features, some fragments show the traces of sutures 0.85 mm apart, while traces of striae, probably representing growth increments are visible in places. Patches of an outer, iridescent layer are also present. One such patch (Fig. 4A5) shows striations about 0.1 mm distant, parallel to the longitudinal axis of the body.
Toward the left side (anterior down) of the posterior portion of conotheca there is a shallow, elongate trough. The trough is aligned approximately parallel to the longitudinal axis of the specimen. The surface of the trough is largely filled and obscured by matrix except at the margins. At the anterior end of the trough three low ridges each about 0.25 mm apart, form overlapping semi-circular arcs 0.95 mm in diameter which close to the anterior. The nature of this structure is unclear, but the trough may have been caused by flexure during compression of the conch. The anterior ridges may represent truncated layers of the phragmocone wall at the anterior end of the trough.
Since one edge of the flattened conotheca is missing, the width of the anterior end of the phragmocone, estimated as twice the distance between the approximate middle of the ink sac and the surviving margin of the flattened phragmocone is 13.7 mm. Assuming that an originally circular phragmocone cross-section was subjected to brittle deformation the restored diameter is estimated at 17.4 mm. Using the trace of the phragmocone margin and assuming both that the direction of the sutures is normal to the conch axis and that deformation was brittle, the restored alveolar/apical angle is estimated at about 31°. On this basis, the estimated length of the phragmocone is at least 31.5 mm, of which at a minimum, 5mm of the posterior of the phragmocone is missing.
Rostrum: There is little evidence that a rostrum was present, but at the posterior end of the preserved portion of the phragmocone there is a small patch of smooth shell that rests on the outer iridescent layer (Fig. 4A5), partially obscuring it. Although only a fraction of a millimetre thick, this layer might represent a remnant of the outer shell layer of the conotheca, the primordial rostrum or an anterior fragment of the orthorostrum.
Remarks.—Many of the arm hooks may be assigned to the parataxon Paraglycerites Eisenack, 1939, and bear a resemblance to hooks assigned to several species of Paraglycerites by Riegraf (1996: figs. 5, 6.1–7) from the bituminous mudstone at the base of the Psilonotenbank (Psilonotum Subchronozone of the Planorbis Chronozone) in the Tübingen area of south-west Germany. Direct comparison with the arm hooks from the Psilonotenbank is hampered by the relatively fragmentary state of preservation of the latter although they both possess a long base with a low angle between the shaft and base, a long shaft, and an uncinus at a high angle to base. They differ in that spurs appear to be less frequent in LYMPH 2023/77, and where visible, located at a relatively distal position (Fig. 7). The fragmentary preservation of the Psilonotenbank specimens makes it difficult to judge whether they might be ubiquitous in that material while the compression, partial embedment and crowding of hooks may be obscuring the presence of spurs present on other hooks in LYMPH 2023/77.

Fig. 7. Coleoidea indet. LYMPH 2023/77. Plot of relative location of spurs on Type A hooks.
Fuchs and Hoffmann (2017: 6) observed that the presence of spurs on arm hooks had at that time, only been reported in rostrum-bearing belemnitids. Engeser (1987) stated that the arm hooks of phragmoteuthids and belemnotheutids lack spurs, although Doguzhaeva et al. (2007: fig. 2b, c) reported the presence of spurs on the hooks in specimens attributed to Phragmoteuthis bisinuata (Bronn, 1859) (see also Hart et al. 2019); now reinterpreted as specimens of Mojsovicsteuthis sp., of uncertain order and family (Lukeneder et al. 2024). Lukeneder et al. (2024: fig. 3) made no mention of the presence of a spur in their hook type 4, observed by Doguzhaeva et al. (2007: 280) to “bear a pronounced spur on the internal side”. Instead they noted the bifurcated nature of the base of the hooks that here are interpreted as the very prominent inner process of a type of hooks that possesses well-developed inner and outer processes. Under this interpretation, it remains the case that the presence of spurs on arm hooks have only been reported in rostrum-bearing belemnitids. No inference can be drawn from this, however, that LYMPH 2023/77 is a rostrum-bearing belemnitid, as the affinities of many spur-bearing hooks remain unknown—including Paraglycerites (Alexander Pohle, personal communication 2014), and are only known from Acrocoelites and Passaloteuthis (Reitner and Urlichs 1983) while the hooks in Hibolithes lack spurs (Klug et al. 2010: figs. 2, 6a).
The location of the dark mass immediately posterior to the arm crown suggests that it may be associated with the buccal mass, and in particular the jaws. However, the extremely poor preservation makes this interpretation highly speculative (Fig. 8). The curved ridge(s) present at one end of the long axis might form a beak-like structure if retro-deformed. Comparison with the jaws of Hibolithes (Klug et al. 2010: fig. 5) from the Kimmeridgian of SW Germany suggests that such an interpretation is feasible if the structure represents the rostrum and part of the hood of the lower jaw while the lateral areas represent parts of the external lamellae. The size of the putative lower jaw is of the same magnitude as that of the Hibolithes although smaller, reflecting the smaller size of LYMPH 2023/77.
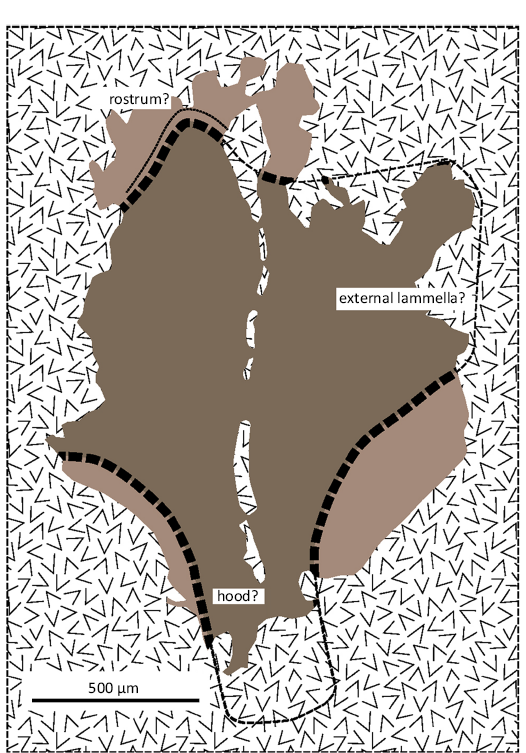
Fig. 8. Coleoidea indet. LYMPH 2023/77. Interpretation of the dark mass posterior to the brachial crown as part of a lower jaw. Dark brown represents heavily sclerotised zone surrounded by more lightly sclerotised (light brown) marginal zone. Except over the purported rostrum, the dashed lines marking the boundary between the two zones are largely speculative (see Fig. 5A8 for comparison). Hatched fill represents layer of comminuted shell debris that obscures parts of the structure and marks out a series of grooves and fissures aligned subparallel to the long axis.
The striae present in isolated patches of the outer, iridescent layer of the conotheca are likely to represent growth increments. Since the orientation of the striae on the margin of the conotheca are different from those visible on the posterior portion of the conotheca, these patches are here interpreted as remnants of the proostracum. The striae parallel to the body axis may represent a fragment of the left hyperbolar zone, and those observed on the lateral edge of the crushed conotheca could be a fragment of the right side of the median field, suggesting that the dorsolateral surface of the specimen faces upwards.
The lack of any conclusive evidence for the presence of a rostrum (or a lack thereof), combined with the fragmentary preservation of the conotheca means that ordinal determination of LYMPH 2023/77 is uncertain, while familial and generic determination is not possible. The possession of arm hooks assignable to Paraglycerites suggests a relationship to the specimens described by Riegraf (1996). The latter is known from the Psilonotum Subzone of Southwest Germany. Psiloceras sampsoni, present in the upper part of Bed 24 in West Somerset (Page and Bloos 1995: fig. 2) is considered a probable senior synonym of P. psilonotum (Bloos and Page 2000). LYMPH 2023/77, from the lower portion of Bed 24, is high in the Planorbis Subzone. Thus, there may be little difference in age between LYMPH 2023/77 and the German material. The belemnitid described by Klug and Fuchs (2010) from Blue Anchor (PIMUZ 27078) is associated with specimens of P. planorbis and is likely to be nearly coeval with LYMPH 2023/77.
Klug and Fuchs (2010) assigned the Blue Anchor specimen (PIMUZ 27078) to the Belemnitida on the basis of the morphology of the well-preserved proostracum. No arm hooks are known from PIMUZ 27078. The proostracum of LYMPH 2023/77 is so fragmentary that its overall morphology must remain speculative. Thus, LYMPH 2023/77 can only be compared with PIMUZ 27078 in the general proportions of the restored phragmocones (Tables 2, 3).
While noting differences which they attributed to the uncertainties of reconstruction and different modes of deformation, Klug and Fuchs (2010) recognised the similarity of PIMUZ 27078 to an Early Jurassic belemnitid specimen described by Hölder (1973) from the Angulata Chronozone of Bünde, Westphalia, Germany. Our attempts at reconstruction of PIMUZ 27078 and Hölder’s (1973) specimen also indicate the similarity of several characters (Table 3), suggesting that the two specimens might represent different growth stages of the same taxon.
Table 3. Comparison of ratios of selected measured characters between PIMUZ 27078, Hölder’s (1973) specimen, and LYMPH 2023/77. Note the similarities the ratios between PIMUZ 27078 and Hölder’s (1973) specimen despite the difference in overall size between the two specimens, and the discrepancies between PIMUZ 27078 and LYMPH 2023/77, Hölder’s (1973) specimen and LYMPH 2023/77.
| |
PIMUZ 27078/ LYMPH 2023/77 |
LYMPH 2023/77 |
PIMUZ 27078 |
|
Maximum phragmocone diameter |
0.56 |
0.38 |
0.68 |
|
Phragmocone length |
0.92 |
0.68 |
0.67 |
|
Proostracum length |
0.72 |
0.49 |
0.68 |
When compared with PIMUZ 27078 and Hölder’s (1973) specimen, LYMPH 2023/77 differs in the larger apical angle of the phragmocone and the depth of the camerae. Assuming that the approximate distance from the anterior end of the phragmocone to the dark mass (jaws?) may represent the length of the proostracum, then it is proportionately slightly shorter than those of PIMUZ 27078 and Hölder’s (1973) specimen although the uncertainty in measurement is likely to be larger than differences in proportionate length. Westermann (1975) observed that for theoretical models of ectocochleate cephalopods, there is an inverse relationship between cameral depth and apical angle. Although this does not hold consistently for fossil ectocochleates, it is possible (although untested) that the relationship holds for coleoids and may indicate that the wider apical angle of LYMPH 2023/77 is real rather than a consequence of the deformation of the phragmocone. Thus LYMPH 2023/77 differs from PIMUZ 27078, at least in the in the larger apical angle of the phragmocone and the depth of the camerae.
To date, it seems that no rostra have ever been reported from the Blue Lias Formation. Given the robust nature of the rostrum, it would be expected that after at least two centuries of collecting they would have been recorded if present. Neither can their absence be explained through dissolution since the originally aragonitic phragmocone is preserved in LYMPH 2023/77 and PIMUZ 27078 and would have been more soluble than the calcitic rostrum. Thus, it seems possible that there was no rostrum present or that it formed an extremely thin sheath in the form of a primordial rostrum.
As the differences in phragmocone morphology between LYMPH 2023/77 and PIMUZ 27078 appear to be real, the two specimens seem likely to be taxonomically distinct. Where known in the Phragmoteuthida, the apical angle is large (25–40°, see Fuchs and Donovan 2018) and also in most (but not all) Diplobelida (15–50°, see Fuchs 2019) while the camerae of both orders are generally narrow. The arm hooks of LYMPH 2023/77, however, are quite different to those of the two Early Jurassic, putative diplobelids, Clarkeiteuthis and Chondroteuthis. Instead, they bear some resemblance to the type 3 hooks of Lukeneder et al. (2024: fig. 3c) associated with Phragmoteuthis bisinuata Bronn, 1859, from the Upper Triassic of Austria (Carnian) Polzber Konservat-Lagerstätte of Lower Austria). Since Phragmoteuthis ranges into the Sinemurian (Donovan 2006; Fuchs and Donovan 2018: fig. 1), it is possible that LYMPH 2023/77 is a phragmoteuthid. Additional, better-preserved material, if discovered, may confirm such an assignment, but here since very little remains of the diagnostic proostracum, LYMPH 2023/77 is placed in open nomenclature.
Order Diplobelida Jeletzky, 1965
Family Chondroteuthidae Jeletzky, 1965
Genus Clarkeiteuthis Fuchs et al., 2013
Type species: Onychoteuthis conocauda Quenstedt, 1849, by original designation (Fuchs et al. 2013: 246). From a horizon in the Paltum Subzone, Tenuicostatum Zone, lower Toarcian, Lower Jurassic at Pliensbach, Holzmaden area, Baden-Württemberg, Germany
Clarkeiteuthis montefiori (Buckman, 1830)
Fig. 9–12.
Material.—Two specimens (LYMPH 2023/76 and LYMPH 2023/78) from the base of Bed 24 of the Planorbis Subchronozone, Planorbis Chronozone, Hettangian Stage, “Psiloceras Beds” of the Blue Lias Formation; foreshore exposures in Doniford Bay, Watchet, UK (LYMPH 2023/76, ST 0804 4349; LYMPH 2023/78, ST 0842 4339). Since there are differences in the preservation of the two specimens and they exhibit different features, they are described separately below.
Discussion
LYMPH 2023/76 (Fig. 9, 10): LYMPH 2023/76 is 152 mm long and consists of an incomplete and disturbed array of arm hooks extending over a distance of 24 mm; a proostracum, of which 60 mm is visible, an ink sac 42 mm long and 24 mm wide, and a 25 mm long portion of a crushed phragmocone that has an estimated anterior width of 27 mm and retains three septa. The posterior end of the ink sack overlaps the anterior end of the of the phragmocone by 12 mm. There is a 33 mm gap between the most anteriorly visible portion of the proostracum and the posterior end of the arm hook array.
Except for the ink sac, the specimen is entirely flattened and probably presents a ventral aspect since the proostracum appears to lie beneath the ink sac. Other than the ink sac, there appears to be no evidence of the presence of soft tissue. Exposure to wave activity has eroded large areas of the proostracum although its anterior and posterior ends, which remain under the matrix may conserve its full width.
Arm hooks: Approximately 60 arm hooks are visible, as well as the impressions of as many as 20 hooks that have been eroded away at the anterior end of the cluster. Many of the hooks are incomplete and most have been fractured, making it difficult to determine whether variation in curvature between hooks reflects a morphological diversity or is a consequence of crushing. Of those hooks sufficiently well-preserved to assess their morphology, all possess bilobed bases (or expand towards the base, suggesting the presence of bilobed bases) combined with a nearly straight to weakly curved shaft and an uncinus which becomes increasingly curved towards the tip in some individuals (Figs. 9A7, A9, A11, 11). Measured from tip to base, hooks range from 1.5–4.0 mm in length, and the base from 1.1–1.3 mm in width. Where preserved, the base appears to being relatively thicker and more robust in larger hooks. In the only hook suitable for measuring the angle between the base and the inflection on the outer side of the hook is 94°, and that between the inflection and the tip of the uncinus is 60°. The distal ends of several hook appear to be more strongly curved and the angle between the inflection and tip of uncinus will be less than 60°. The smaller hooks are more frequent at the posterior end of the cluster. Although many of the hooks appear to have been disturbed, pairs of hooks belonging to the same arm remain evident, and many as four rows of paired hooks may be present (Fig. 10).

Fig. 9. Coleoid cephalopod Clarkeiteuthis montefiori (Buckman, 1830) from the Early Jurassic, Hettangian Stage of Doniford Bay, Watchet, UK. A. LYMPH 2023/76. A1, image of whole specimen; A2, interpretative sketch mapping out the distribution of the surviving components of the organism; A3, enlargement of posterior end of specimen showing undifferentiated remains of septa and phragmocone wall; A4, detail of weathered surface of proostracum showing imbricate tabular layers; A5, posterior end of proostracum showing cracked surface, patches of carbonaceous material underlying the proostracum (white arrowhead), carbonaceous material in a major crack (black arrowhead), and one of the depressions present on the ventral surface of the proostracum (see also A10); A6, detail of depression on the ventral surface of the proostracum showing the arcuate ridges and furrows (arrowhead) probably formed by the migration of muscle attachments; A7, pair of worn and broken arm hooks (white arrowhead in A9) with strongly curved uncinus; A8, A9, explanatory drawing and image of arm hooks forming remains of brachial crown; A10, carbonaceous material underlying crack (black arrowhead in A5) showing linear features that may be corrugation of an organic membrane; A11, arm hook (black arrowhead in A9) showing broad base and basal processes.

Fig. 10. Coleoid cephalopod Clarkeiteuthis montefiori (Buckman, 1830). LYMPH 2023/76. Sketch of distribution of arm hooks and probable hook pairing with likely traces of arms (dashed lines).
Proostracum: The width of the remaining part of the proostracum is 29.3 mm at the posterior end and 18.9 mm at the anterior. It is at least 0.2 mm thick at the lateral margins. The width at the anterior end is likely to reflect the true width since anterior of this point, the proostracum is covered by matrix, so that the width observed immediately adjacent to the matrix cover is unlikely to have been modified by recent erosion. The dorsal surface of the proostracum faces into the matrix and is therefore not visible. The ventral surface is visibly heavily cracked; it consists of apparently semi-translucent material varying in colour from off-white to pale blue, through to a deep brown. It is iridescent in places. The surface, except where eroded, appears smooth. The brown tinted zone forms a longitudinal band about 2 mm wide. The major cracks are 2.0–1.0 mm apart, while more frequent cracks 0.5–1.0 mm distant form a lattice between the major cracks. The minor cracks are most densely distributed towards the lateral margins away from the brown-tinted zone. The distribution of the cracks suggests that either the cross-section of the proostracum was more strongly curved and/or thinner towards the lateral margins.
Under magnification, parts of the ventral surface show a very thin layer that appears to be extremely finely crystalline. This layer may have originally been extensive, covering the whole ventral surface, but has been lost to erosion. Eroded surfaces beneath this outer layer indicate the presence of a stack of thin laminae, each estimated to be no more than 0.02 mm (20 μm) in thickness. The laminae themselves are slightly inclined to the surface of the proostracum such that they are imbricate. Where the laminae intersect with the surface, they are aligned at approximately 60° to the long axis of the phragmocone and dip to the left anterior as viewed from the dorsal surface. Fine cracks, visible as white lines up to 10 mm long follow this alignment.
A pair of depressions are visible on the surface of the proostracum 8 mm in front of the anterior end of the ink sac. The depression on the right (as viewed from the dorsum) consists of series of curved ridges about 0.25 mm distant with furrows in between (Fig. 9A5, A6). The convex sides of the curves face the anterior and the more anterior ridges partially overlap those that are more posterior, particularly where directed inward from the margin of the proostracum. This structure is about 2 mm wide. The inward directed portions of the ridges look as though they should extend further across the proostracum, although they diminish in that direction. About 5 mm laterally from this structure, a second smaller, less prominent depression and set of curved ridges are visible with the convex sides facing the anterior. These are believed likely to form part of the same structure, which may be interpreted as comprising broadly curving ridges and furrows extending laterally across part of the proostracum.
Thin layers of dark, carbonaceous material are visible in between the larger cracks in the proostracum (Fig. 9A5). They also form small patches on its ventral surface. Whether this material represents originally organic tissue or leaked ink is unclear, but the carbonaceous material associated with the dorsal surface of the proostracum appears to line its surface. It does not extend into the matrix, and would, therefore, seem likely to represent an originally organic structure. Approximately halfway across the width of the preserved part of the proostracum, a narrow carbonaceous band extending the length of the proostracum is visible. This structure appears to be attached to the dorsal surface of the proostracum and is visible between and below a line of longitudinal cracks (Fig. 9A5, A10). The structure shows several ridges and grooves parallel to its axis suggesting that it may be crumpled and folded. This structure may be the same as that visible in Chondroteuthis wunnenbergi Bode, 1933 (Fuchs 2019: fig. 4c) and Clarkeiteuthis conocauda (Quenstedt, 1849) (Fuchs 2019: fig. 5b)
Ink sac: The ink sac is partially crushed, and its surface, as currently preserved suggests that the wall may have been folded or even ruptured during burial. Patches of matrix discoloured by carbonaceous material surrounding the ink sac may be evidence of ink leakage. Overall, the shape of the sac is prolate but tapers into a funnel at the anterior end.
Phragmocone: The phragmocone is poorly preserved. The apical portion is missing while the remaining septae and phragmocone wall are completely crushed and flattened (Fig. 9A3). Based on what little remains of the phragmocone, the apical angle is estimated at 20°. The depth of the two remaining camera varies markedly, with the anterior camera having a depth 0.27% of the phragmocone diameter, and the posterior, a depth of 0.16%, giving an average of 0.23%.
LYMPH 2023/78 (Figs. 11, 12): The specimen consists of an ink sac 25 mm long, ink duct, mantle, and traces of the body extending over a distance approximately 65 mm anterior to the ink sac. The phragmocone is not visible and any shell or tissue to the posterior of the ink sac was either lost prior to burial or to more recent wave action. With the exception of the ink sac and ink duct, the remaining part of the body is largely flattened. The outline of the mantle is 21 mm wide anterior to the ink sac and is constricted to 9 mm, 21 mm anteriorly. The remainder of the outline of the mantle maintains that width for the rest of its length. A jaw-like structure is visible 30 mm anterior to the ink sack toward the anterior extremity of the body. Part of the arm crown is exposed in a break oblique to the lamination of the matrix on the underside of the block. The length of the specimen from the posterior of the ink sac to the exposed part of the arm crown is about 110 mm.
Between the lateral margins of the presumed mantle, with the exception of the ink sac and ink duct, the body has been compressed to a thickness of 0.3–0.4 mm. However, the surface of the compressed body is uneven and consists of a pair of lateral ridges that outline the margins of the body and the mantle, and a pair of longitudinal ridges on either side of the median axis. The latter diverge posteriorly to join the ridges at the margins, creating a cavitiy either side of the anterior end of the ink sac. A similar feature is seen in a specimen of Clarkeiteuthis conocauda figured by Reitner (2009: 290, fig. 1) from the Toarcian, Lower Jurassic, of Swabia. Reitner (2009) described the remains of gills preserved within the cavities of the Swabian specimen.
The ridges and inter-areas that appear to comprise the remains of the body are composed of a black, hard waxy material that is indistinguishable from the that of the ink sac under reflected white light. Leakage of ink into the mantle cavity was reported by Reitner (2009) and Jenny et al. (2019) for another specimen. It is likely that the material lining the mantle cavity is leaked ink.
There are several flattened conchs of Psiloceras associated with the specimen. At least one individual is impressed into the probable leaked ink at the anterior end of the mantle leaving a mould of the of the conch surface.
Arm hooks: Since the arm hooks are only visible in an oblique fracture on the underside of the specimen, the area where they are visible is too small to determine whether they represent individual arms or a disturbed cluster, although at least one example of paired hooks is visible. Many of the arm hooks are represented by fragments, having been broken by the fracturing of the matrix. Of the less damaged hooks, most are partially obscured by matrix. Two better exposed hooks (Fig. 11A6), measure respectively 0.42 and 0.38 mm at the base of the shaft and the shafts 1.09 and 1.17 mm long. The basal processes have been broken off in both hooks. In a third hook (Fig. 11A6), the base is complete and has a width of 0.8 mm. The inner process is longer than the outer process. In the first hook, the uncinus is 0.85 mm long but is incomplete in the second hook. The curvature is moderate. The angle of the distal end of the uncinus is at 51° to the shaft in the first hook. Other incomplete hooks have a more strongly curved uncinus such as that in one hook (Fig. 11A6) the tip is at 70° to the shaft. Several of the hooks show grooves running along their lateral surfaces. These may have resulted from the compression of the hook and the collapse of the internal cavity associated with the basal opening but could also reflect a primary striation.
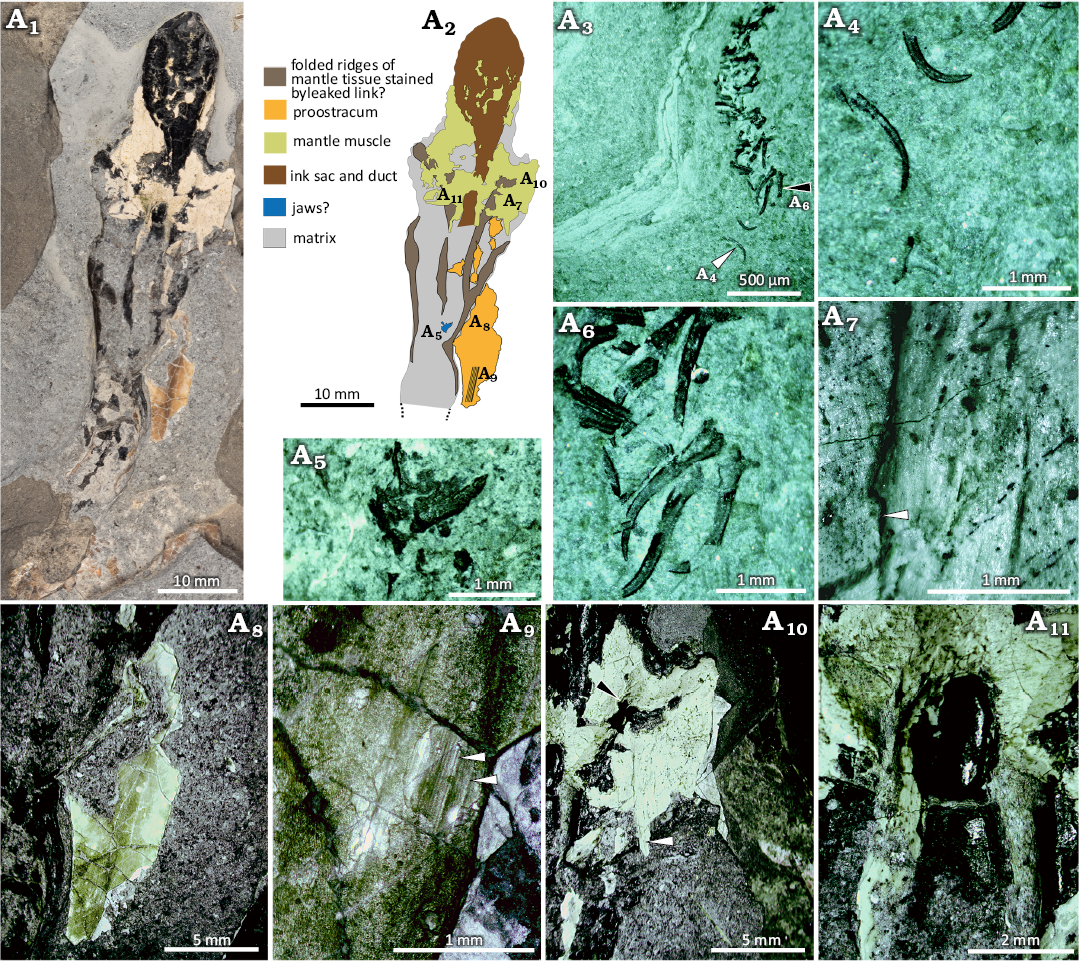
Fig. 11. Coleoid cephalopod Clarkeiteuthis montefiori (Buckman, 1830). LYMPH 2023/78 from the Early Jurassic, Hettangian Stage of Doniford Bay, Watchet, UK. A1, image of whole specimen; A2, interpretative sketch mapping out the distribution of the surviving preserved components of the organism; A3, oblique section through the brachial crown, partially exposed on the underside of the specimen; A4, isolated hooks with bases (white arrowhead in A3) and part of the shaft missing but with more strongly curved uncinus; A5, possible jaw with the rostrum below; A6, arm hooks (black arrowhead in A3) with moderately curved uncinus, long tapering shafts and bases that are broken but indicate the presence of broad basal processes; A7, detail of mantle muscle showing division into bands of finely striated muscle, note also the overlap and discontinuity (arrowhead) suggesting tearing or folding of the mantle; A8, fragment of proostracum overlapping and resting on main body of the mantle cavity and exhibiting longitudinal linear banding; A9, detail of proostacum with coarser linear ridges (arrowheads), between which numerous finely striate, parallel and linear longitudinal ridges are visible; A10, thin layer of mantle muscle on lateral flanks showing a nearly homogenous fabric but with some fain banding visible, banding appears to cross the image nearly horizontally while, a separate band (white arrowhead) exhibits vertical bands. One zone (black arrowhead) appears to be corrugated and twisted; A11, mantle muscle developed over and to either side of the ink duct showing a broadly circumferential grain over the ink duct but with a grain parallel to the duct further anterior, possibly representing longitudinal muscle.
Jaws?: About 30 mm in front of the anterior end of the ink sac, a structure tentatively interpreted as a jaw floats in the matrix (Fig. 11A5). The surface of this structure is worn and has probably been deformed while one lateral half remains largely obscured by matrix. If bilaterally symmetrical, the width of the structure would be 3 mm and its length 1.8 mm. The anterior edge is arcuate, but projects forward on either side of the plane of bilateral symmetry to form a sharp apex (compare Klug et al. 2020). Behind the anterior edge, the surface appears relatively smooth although sediment infills parts of the middle zone and is also present in a zone subparallel to the anterior edge of the structure. This suggests the presence of a cavity under the anterior area. In the posterior part of the structure, approximately along the plane of bilateral symmetry, a narrow ridge extends out to the posterior edge. Beyond the anterior apex and floating isolated in the matrix, there is a needle-like structure 0.35 mm long and 0.03 mm wide at its base. This structure points anteriorly and is roughly aligned with the apex of the anterior edge and the plane of bilateral symmetry.
Mantle and mantle muscle: Details of the outline, extent and general features of the mantle were described above. The mantle musculature is described below.
Mantle muscle is preserved anterior to the ink sac forming a band 18 mm long. Traces of muscle are also visible embedded onto the surface of the ink sac. A whitish layer approximately 0.05 mm thick, underlying the leaked ink, may also represent mantle muscle. If this is the case, then the preserved mantle muscle extends around the circumference of the body in this area. The surfaces of the mantle muscles are worn and pockmarked from the holdfasts of marine algae. Despite the wear, faint striations are visible (Fig. 11A7, A10, A11) which (Fig. 11A7) resolve into semi-translucent bands about 0.01 mm across at higher magnifications; these are bordered by bands of more opaque material tentatively interpreted as the remains of muscle fibres as illustrated by Kear et al. (1995; text-fig 4a, b). On the exposed side of the specimen, the muscle varies in thickness from a thin film of about 0.1 mm to zones where it may reach 0.25 mm.
The orientation of the striations indicates that most of the preserved muscle is probably represented by circular muscle (Fuchs et al. 2016). However, in several places the striations deviate so strongly from circumscribing the body axis that they are oriented longitudinally (Fig. 12). However, there appears to be no consistent pattern to the orientation of the muscle fibres. Much of the deviation from a circumscriptive distribution appears to be associated with the zones where the layer of muscle is thicker. The boundaries between zones where the muscles are thicker or thinner often exhibit cross-cutting relationships in the directions of the muscle fibres (Fig. 11A10). The thickening of the muscle combined with the relationships of the striations suggest that the mantle has been deformed and twisted in these zones, possibly caused by postmortem shrinking and tearing of the mantle (Clements et al. 2017). In narrow bands either side of the ink duct (Fig. 11A11), muscle tissue with a grain parallel to the body axis extends a short distance anteriorly.
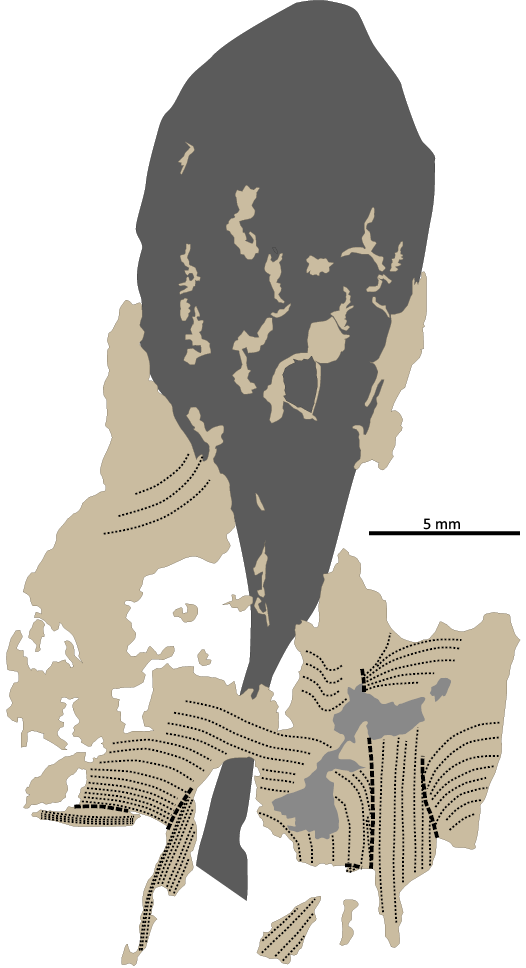
Fig. 12. Coleoid cephalopod Clarkeiteuthis montefiori (Buckman, 1830). LYMPH 2023/78. Orientation of mantle muscle fibres. Dark grey, ink sac; grey, leaked ink underneath mantle muscle; beige, mantle muscle; fine dashed lines, alignment of muscle fibres; coarse dashed lines, overfolds or tears in the mantle muscle.
Proostracum: Several small fragments of the proostracum remain. The largest of these (Fig. 11A8) is 20 mm long by 9 mm wide and is comprised of at least three layers. The lowest layer forms a sheet that is probably no thicker than 10–20 μm. The middle layer is a semitranslucent sheet of a thickness estimated to vary from 0.2–0.3 mm and appears to be thickest posteriorly and toward the middle of its width. Over much of the area, the upper surface of the middle layer is exposed as the uppermost layer is missing. The surface, however, consists of a series of linear striations that are oriented roughly parallel to the axis of the body. The striations have a distinct relief and a density of 40 per mm. At an approximately 0.2 mm spacing, the striae are more prominent, and form narrow longitudinal ribs. The outermost layer is extremely thin (possibly <10 μm) and preserved only in small patches where the prominence of the striae appears to be greater. The proostracum contains a network of large matrix-filled cracks as well as much narrower, incipient, and regularly spaced cracks oriented at approximately 30° to the striae. One edge of this fragment of proostracum overlaps with and appears to lie above the compressed body cavity. The other remaining patches of the proostracum consist of no more than it lowest layer. rest on the body cavity, apparently at the same level as the mantle muscle and although there appears to be a marginal overlap with the mantle muscle in places, it is not possible to determine the order in which they lie. It is possible that muscle tissue is preserved above and below the proostracum.
Ink sac and ink duct: The ink sac is 15 mm long and 9.3 mm wide. It is constricted anteriorly to form an ink duct 1.5 mm in diameter. The surfaces of both the ink sac and the ink duct are covered in a tessellation of cracks, indicating that the contents of both had largely solidified prior to the compression of the rest of the remains of the body.
Remarks.—As the remaining large fragment of proostracum (LYMPH 2023/78) may appear similar to the pale laminae interpreted as muscle tissue toward the posterior end of the specimen, the difference in fabric is emphasised here. The former consists of a dense and translucent, probably crystalline material, whereas the latter is only marginally translucent and exhibits a very fine striation that completely penetrates the material. Since this specimen lacks the phragmocone, and the proostracum is too fragmentary for its general overall shape to be deduced, identification relies upon the similarity of the arm hooks to those of Clarkeiteuthis conocauda and C. montefiorei (Buckman, 1880) as well as the characteristic shape of the body with the anterior constriction in the diameter of the mantle.
The muscular mantle is inserted into the lateral margins of the proostracum (Fuchs et al. 2016: 436). Jenny et al. (2016) referred to the mantle associated with the phragmocone of the specimen of a specimen they described C. conocauda as thin or “skinny” (Jenny et al. 2016: 3). Since the mantle muscle was not performing a locomotory function over such areas, powerful muscle was not required. Although in LYMPH 2023/78 the remains of muscle sheets appear to be at their thinnest over the ink sac and duct, where a few fragments proostracum are also present further adorally, it is not clear from the current state of preservation whether these fragments were located above or below the muscle sheet, or whether these proostracum fragments are at their original location. Furthermore, where muscle sheet is preserved over the ink sac, there is no sign of a proostracum located between the muscle sheet and the ink sac. This is difficult to understand and there may be several explanations that are not necessarily excusive to each other:
(i) the fabrics interpreted as muscle sheets are in fact proostracum; this seems unlikely given the strongly contrasting nature of the fabrics attributed to these structures;
(ii) the proostracum originally underlay the muscle sheet but became separated and was extruded during the partial decay of the mantle; there is some evidence for this in that the largest and apparently thickest remaining proostracum fragment appears to lie outside of the body cavity; and
(iii) the proostracum was originally located above the preserved muscle sheets and the muscle preserved below it was attached to its ventral side; note that the grain of some of the musculature in this region is oriented parallel to the body axis as it would if it represented head or funnel retractors.
Additional material may help resolve this conundrum.
Geographic and stratigraphic range.—Early Hettangian to late Sinemurian of Southwest England.
Discussion
The arm hooks observed in LYMPH 2023/76 and LYMPH 2023/78 with their bilobed bases, perpendicular shafts and weakly curved uncinus were regarded as unique to Clarkeiteuthis (Fuchs et al. 2013). Following Donovan (2006), Fuchs et al. (2013) did not attempt to synonymise Clarkeiteuthis montefiori with C. conocauda on the grounds that the Sinemurian C. montefiori remained insufficiently known and differed in age from the Toarcian C. conocauda. Hart and Hutchinson (2016) and Hart et al. (2019) regarded C. conocauda and C. montefiori as distinct species, observing that in C. montefiori, the type specimen “…clearly show pairs that have a long hook with a bi-lobed base alongside smaller, almost triangular hooks” (Hart and Hutchinson 2016: 37), while in a specimen from Bristol Museum and Art Gallery (BRMSG Ce12385) “…one of the pair has a long, slightly curved, shaft the other part of the pair is smaller and does not have an elongated shaft” (Hart and Hutchinson 2016: 37). Hart et al. (2019) further elaborated on BRMSG Ce12385, noting that one component of the pair was more variable with some that looked subtriangular, while others were more elongated, resembling those of the other complementary row. Although the hooks of each half pair do appear different, their differing orientations and variable degree of burial in the matrix makes the assessment of differences challenging. This is likely to be exacerbated in the type since it is preserved in a fissile mudstone and the hooks may have been compressed and deformed during burial. Both specimens show changes in hook size along the length of the arms, but this is also apparent in C. conocauda and is a common feature of belemnoids (e.g., Donovan and Crane 1992; Klug et al. 2010; Garassino and Donovan 2000: text-figs. 2–4, 7).
Jenny et al. (2019: fig. 1) described a jaw (possibly lower and upper jaw) in a specimen of Clarkeiteuthis conocauda from the Toarcian Posidonienschiefer of Holzmaden, Germany. At 10 mm in width the jaw(s) are substantially larger than LYMPH 2023/78, despite the two specimens being of a broadly similar size. It is possible that only the most strongly sclerotised part of the jaw is preserved in LYMPH 2023/78. Jenny et al. (2019: 6) noted a more strongly sclerotised zone, possibly associated with the inner and outer lamellae preserved in the Holzmaden specimen. The relative location of the structure in LYMPH 2023/78 also differs from that of the Holzmaden specimen as is located nearer the ink sac anterior to the ink sac. Since the purported jaw in LYMPH 2023/78 has been substantially compressed and probably rotated out of its original orientation, the broad angle of the anterior margin may reflect this compression, resulting in the shortening of the anterior-posterior axis. If interpreted as a lower jaw, the anterior margin would represent the outer lamella, the anterior apex and needle-like structure the rostrum, with the terminal portion broken away. It could thus have had an elongate rostrum as documented for a Middle Jurassic belemnite by Klug et al. (2020). The posterior ridge may be the crest and the areas to either side, the right and left wings of the inner lamella.
Although the proostracum is fragmentary and therefore undiagnostic in LYMPH 2023/78, the constriction of the anterior part of the body seen in this specimen is characteristic of Clarkeiteuthis (Fuchs et al. 2013). The poor preservation of the proostracum in LYMPH 2023/76, and in particular the lack of the lateral margins, casts doubt on this assignment. However, at 18.9 mm, the width of the anterior end of the preserved portion of the proostracum is similar to that of C. conocauda while the length/width ratio, based on the distance from the anterior end of the phragmocone to the posterior of the arm crown is 0.174, very close to 0.17 stated for C. conocauda (Fuchs et al. 2013: 250). The significance of other features of the proostracum of LYMPH 2023/76 are less clear. A feather-like pattern on the inner (ventral) surface of the proostracum was reported in several belemnitid genera (Doguzhaeva 2012) including a specimen described by Crick (1896) and assigned to Chondroteuthis (Donovan in Doguzhaeva et al. 2002; Doyle and Shakides 2004) indicates that this feature extends to putative diplobelids. Although much of the inner (ventral) surface of the proostracum of LYMPH 2023/76 appears to be smooth, the depressions and associated arcuate ridges and furrows apparent in one area (Fig. 9A6) bear a resemblance to those on the ventral surface of Chondroteuthis wunnenbergi (Doyle and Shakides 2004: text-fig. 3a) and is of the same relative scale. Moreover, if the longitudinal carbonaceous structure visible about halfway across the width of the proostracum is interpreted as a median line or ridge, then this structure is located in the same relative position on the proostracum and may be interpreted as representing a remnant of the feather-like pattern reported by Doguzhaeva (2012). Dirk Fuchs (personal communication 2024) indicated that these structures mar represent muscle attachment sites.
Fuchs et al. (2013) estimated apical angles of 30–35° for the specimens of Clarkeiteuthis conocauda they figured. Thus, the estimate of 20° for LYMPH 2023/76 seems somewhat low unless it represents a taxon distinct from C. conocauda. Donovan (2006) noted that the apical angle decreased with growth in a specimen from the Bucklandi Chronozone of south-west Germany described by Riegraf (1982). He estimated the apical angle of the mature part of the phragmocone in this specimen at 22°, similar to LYMPH 2023/76. Estimation of the apical angle in crushed material is reliant on the correct interpretation of the nature of deformation. Uncertainty as to whether deformation was brittle or plastic can lead to large under and overestimates of the apical angle, as well as other characters. The apical angle for two of the specimens illustrated by Fuchs et al. (2013: fig. 2a, b) may be estimated at 40° and 32° respectively if deformation was plastic, or 34° and 20° if brittle. It is likely that combinations of plastic and brittle deformation are involved in each case, so that these estimates are the upper and lower limits of the range. The value of 20° in LYMPH 2023/76 assumes plastic deformation. Assuming fully brittle deformation results in an angle of 13°, similar to that of a specimen of Chondroteuthis wunnenbergi figured by Doyle and Shakides (2004: fig. 3d, e). The nature of the arm hooks in LYMPH 2023/76 and LYMPH 2023/78 rule out the possibility that they belong to C. wunnenbergi. The only clear reference to cameral depth in specimens attributed to Clarkeiteuthis conocauda by Fuchs and Donovan (2013) refers to their fig. 2c–e. In measurements derived from the figure, cameral depth is respectively (anterior to posterior) 0.126 and 0.15 of the phragmocone diameter, and thus shorter than those apparent in LYMPH 2023/76. In conclusion, the lack of sufficiently well-preserved material attributed to either C. conocauda or C. montefiori makes recognition of distinctions between the two taxa equivocal. Thus, the position reached by Donovan (2006) and followed by Fuchs et al. (2013) that C. montefiori differed in stratigraphical age and was too poorly characterised to synonymise with C. conocauda is followed here and the specimens described above are assigned to C. montefiori. Future discovery of better-preserved material may clarify the relationship between these two taxa.
Conclusions
Bed 24 of the Planorbis Subchronozone of the Blue Lias Formation of the West Somerset coast rarely contains exceptionally well-preserved coleoid remains that include the diplobelid Clarkeiteuthis, an indeterminate coleoid taxon possessing some features that suggest it may be a phragmoteuthid and a possible belemnitid described by Klug and Fuchs (2010). Together with arm hooks attributable to diplobelids (found in the stomach contents of an ichthyosaur from the Planorbis Subchronozone of the Dorset coast) as well as arm hooks attributable to the arm hook type assigned to Paraglycerites (from the Psilonotum Subchronozone of SW Germany), these finds suggest that phragmoteuthds and putative diplobelids may have been relatively widespread in northern Europe during the early Hettangian, albeit with low diversities. Indirect evidence of their relative abundance may be reflected in the presence of large numbers of arm hooks that are probably attributable to Clarkeiteuthis in the stomach of an ichthyosaur (Pollard 1968, 1990), suggesting that they were probably regular prey items. The apparent lack of coleoid body fossils in the Hettangian sediments of northern Europe may result from a lack of appropriate preservational conditions and/or low expectations of finding them. The Somerset specimens unequivocally demonstrate their presence in sediments of this age and help plug a gap in knowledge, extending coleoid records into the earliest Jurassic, shortly after the end Triassic mass extinction event.
Acknowledgements
CK thanks Kenji Ikuno (Museum of Nature and Human Activities, Hyogo, Japan) for being of great help during our joint field work. The authors thank the Alexander Pohle (Ruhr-Universität Bochum, Germany) and Dirk Fuchs (Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany) for their constructive reviews which substantially improved the manuscript. KNP, DHE, and AK wish to thank the Orchard-Wyndham Estate for permission for scientific sampling of the foreshore area. KNP would also like to thank the Fundación María José Bello Villalba (Crediton, UK) for ongoing support.
References
Bandel, K. and Spaeth, C. 1988. Structural differences in the ontogeny of some belemnite rostra. In: J. Wiedemann and J. Kullmann (eds.), Cephalopods—Present and Past, 247–271. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart.
Bloos, G. and Page, K.N. 2000. The basal Jurassic ammonite succession in the NW European province—review and new results. GeoResearch Forum 6: 2–40.
Buckland, W. 1836. Geology and Mineralogy Considered with Reference to Natural Theology. 2+xvi+619; x+129 pp. William Pickering, London. Crossref
Clements, T., Colleary, C., De Baets, K., and Vinther, J. 2017. Buoyancy mechanisms limit preservation of coleoid cephalopod soft tissues in Mesozoic lagerstätten. Palaeontology 60: 1–14. Crossref
Crick, C.G. 1896. On the proöstracum of a belemnite. Proceedings of the Malacological Society of London 3: 117–119.
Delsate, D., Duffin, C.J., and Weis, R. 2002. A new microvertebrate fauna from the Middle Hettangian (Early Jurassic) of Fontenoille (Province of Luxembourg, south Belgium). Memoirs of the Geological Survey of Belgium 48: 1–84.
Davis, R.A., Mapes, R.H., and Klofak, S.M. 1999. Epizoa on externally shelled cephalopods. In: A.V. Rozanov and A.A. Shevyrev (eds.), Fossil Cephalopods: Recent Advances in Their Study, 32–51. Russian Academy of Sciences, Palaeontological Institute, Moscow.
Dick, D.G., Schweigert, G., and Maxwell, E.E. 2016. Trophic niche ontogeny and palaeoecology of early Toarcian Stenopterygius (Reptilia: Ichthyosauria). Palaeontology 59: 423–431. Crossref
Denton, E.J. 1974. On buoyancy and the lives of modern and fossil cephalopods. Proceedings of the Royal Society of London. Series B. Biological Sciences 185: 273–299. Crossref
Doguzhaeva, L.A. 2012. The original composition of the pro-ostracum of an early Sinemurian belemnite from Belgium deduced from the mode of preservation and ultrastructure. Palaeontology 55: 249–60. Crossref
Doguzhaeva, L.A., Mutvei, H., and Donavan, D.T. 2002. Pro-ostracum, muscular mantle and conotheca in the Middle Jurassic belemnite Megateuthis. In: H. Summesberger, K. Histon, and A. Daurer (eds.), Cephalopods—Present and Past. Abhandlungen der Geologischen Bundesanstalt Wien 57: 321–339.
Doguzhaeva, L.A., Mutvei, H., and Weitschat, W. 2003. The pro-ostracum and primordial rostrum at early ontogeny of Lower Jurassic belemnites from north-western Germany. Berliner Paläobiologische Abhandlungen 3: 79–89.
Doguzhaeva, L.A., Summesberger, H., Mutvei, H., and Brandstaetter, D. 2007. The mantle, ink sac, ink, arm hooks and soft body debris associated with the shells in Late Triassic coleoid cephalopod Phragmoteuthis from the Austrian Alps. Palaeoworld 16: 272–284. Crossref
Donovan, D.T. 2006. Phragmoteuthida (Cephalopoda: Coleoidea) from the Lower Jurassic of Dorset, England. Palaeontology 49: 673–684. Crossref
Donovan, D.T. and Crane, M.D. 1992. The type material of the Jurassic cephalopod Belemnotheutis. Palaeontology 35: 273–296.
Doyle, P. 1993. Mollusca: Cephalopoda (Coleoidea). In: M.J. Benton (ed.), The Fossil Record 2, 229–236. Chapman and Hall, London.
Doyle, P. 1994, Aspects of the distribution of Early Jurassic belemnites. Proceedings of the Third Pergola International Symposim: Palaeopelagos, Special Publication 1: 109–120.
Doyle, P. and Shakides, E.V. 2004. The Jurassic belemnite suborder Belemnotheutina. Palaeontology 47: 983–998. Crossref
Engeser, T. 1987. Belemnoid arm hooks (“onychites”) from the Swabian Jurassic: A review. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 176: 5–14. Crossref
Fuchs, D. 2019. Part M, Chapter 23E: Systematic descriptions: Diplobelida. Treatise Online 118: 1–8. Crossref
Fuchs, D. 2023. Part M, Chapter 23F: Systematic descriptions: Decabrachia. Treatise Online 171: 1–41.
Fuchs, D. and Donovan, D.T. 2018. Part M, Chapter 23C: Systematic descriptions: Phragmoteuthida. Treatise Online 111: 1–7. Crossref
Fuchs, D. and Hoffmann, R. 2017. Arm armature in belemnoid coleoids. Treatise Online 91: 1–20.
Fuchs, D., Donovan, D.T., and Keupp, H. 2013. Taxonomic revision of “Onychoteuthis” conocauda Quenstedt, 1849 (Cephalopoda: Coleoidea). Neues Jarbuch für Geologie und Paläontologie Abhandlungen 230: 245–255. Crossref
Fuchs, D., Iba, Y., Tschlinger, H., Keupp, H., and Klug, C. 2016. The locomotion system of Mesozoic Coleoidea (Cephalopoda) and its phylogenetic significance. Lethaia 49: 433–454. Crossref
Garassino, A. and Donovan, D.T. 2000. A new family of coleoids from the Lower Jurassic of Osteno, Northern Italy. Palaeontology 43: 1019–1038. Crossref
Hart, M.B. and Hutchinson, D. 2016. A newly described Jurassic “squid-like cephalopod” from the Lias Group of Dorset. Geoscience in South-West England 14: 35–38.
Hart, M.B., Hughes, Z., Page, K.N., Price, G.D., and Smart, C.W. 2019. Arm hooks of coleoid cephalopods from the Jurassic succession of the Wessex Basin, Southern England. Proceedings of the Geologists’ Association 130: 326–338. Crossref
Hölder, H. 1973. Miscellanea cephalopodica. V. Ein belemnitisches proostracum aus dem unteren Lias. Münstersche Forschungen zur Geologie und Paläontologie 29: 63–76.
Iba, Y., Sano, S., and Mutterlose, J. 2014. The early evolutionary history of belemnites: new data from Japan. PLOS ONE 9 (5): e95632 Crossref
Iba, Y., Sano, S., Mutterlose, J., and Kondo, Y. 2012. Belemnites originated in the Triassic—a new look at an old group. Geology 40: 911–914. Crossref
Jenny, D., Fuchs, D., Arkhipkin, A.I., Hauff, R.B., Fritschi, B., and Klug, C. 2019. Predatory behaviour and taphonomy of a Jurassic belemnoid coleoid (Diplobelida, Cephalopoda). Scientific Reports 9: 7944. Crossref
Kear, A.J., Briggs, D.E.G., and Donovan, D.T. 1995. Decay and fossilization of non-mineralized tissue in coleoid cephalopods. Palaeontology 38: 105–131.
Klug, C. and Fuchs, D. 2010. An earliest Hettangian (Jurassic) belemnitid from Great Britain with a preserved proostracum. In: K. Tanabe, Y. Shigeta, T. Sasaki, and H. Hirano (eds.), Cephalopods—Present and Past, 181–185. Tokai University Press, Tokyo.
Klug, C. and Korn, D. 2001. Epizoa and post-mortem epicoles on cephalopod shells—examples from the Devonian and Carboniferous of Morocco. Berliner geowissenschaftliche Abhandlungen (E) 36: 145–155.
Klug, C., Etter, W, Hoffmann, R., Fuchs, D., and De Baets, K. 2020. Large belemnite jaws from the Aalenian (Middle Jurassic) of Switzerland. Swiss Journal of Palaeontology 139: 1–11. Crossref
Klug, C., Schweigert, G., Fuchs, D., and Dietle, G. 2010. First record of a belemnite preserved with beaks, arms and ink sac from the Nusplingen Lithographic Limestone (Kimmeridgian, SW Germany). Lethaia 43: 445–456. Crossref
Klug, C., Schweigert, G., Hoffmann, R., Weis, R., and De Baets, K. 2021. Fossilized leftover falls as sources of palaeoecological data: a “pabulite” comprising a crustacean, a belemnite and a vertebrate from the Early Jurassic Posidonia Shale. Swiss Journal of Palaeontology 140: art. 10(2021). Crossref
López-Córdova, D.A., Avaria-Llautureo, J., Ulloa, P.M., Braid, H.E., Revell, L.J., Fuchs, D., and Ibáñez, C.M. 2022. Mesozoic origin of coleoid cephalopods and their abrupt shifts of diversification patterns. Molecular Phylogenetics and Evolution 166: 107331. Crossref
Lukeneder, P., Fuchs, D., and Lukeneder, A. 2024. Morphology, taxonomy and trophic interactions of rostrum-less coleoids from the Late Triassic Polzberg Konservat-Lagerstätte (Lower Austria). Swiss Journal of Palaeontology 143: art. 21(2024). Crossref
Ma, Z., Zhang, T., Chen, J., Popa, M. E., Li, H., Li, S., Zeng, J., and Zhang, X. 2023. The earliest belemnite linked with the Carnian Pluvial Episode. Frontiers in Ecology and Evolution 11: 1236222. Crossref
Niko, S. and Ehiro, M. 2022. Tohokubelus gen. nov., the oldest belemnite from the Olenekian (Lower Triassic) of Northeast Japan. Paleontological Research 26: 115–123. Crossref
Page, K.N. 2002. A review of the ammonite faunas and standard zonation of the Hettangian (Lower Jurassic) of the east Devon coast (south-west England). Geoscience in south-west England 10: 293–303.
Page, K.N. 2003. The Lower Jurassic of Europe: its subdivision and correlation. Geological Survey of Denmark and Greenland Bulletin 1: 23–59. Crossref
Page, K.N. 2005. The Hettangian ammonite faunas of the West Somerset Coast (South West England) and their significance for the correlation of the candidate GSSP (Global Stratotype and Point) for the base of the Jurassic System at St. Audries Bay. In: M. Hanzo (coord.), Còlloque l’Hettangien à Hettange de la science au patrimoine, 15–19. Université Henri Poincarré, Nancy.
Page, K.N. 2017. From Oppel to Callomon (and beyond!): Building a high resolution ammonite-based biochronology for the Jurassic System. Lethaia 50: 336–355. Crossref
Page, K.N. and Bloos, G. 1995. The base of the Jurassic system in West Somerset, south-west England—new observations on the succession of ammonite faunas of the lowest Hettangian Stage. Geoscience in South-West England 9: 231–235.
Page, K.N. and Hart, M.B. 2018. A new record of a basal Jurassic belemnoid squid from Somerset, south-west England. Geoscience in South-West England 14: 243.
Page, K. and Paul, C.R.C. 2017. Articulated echinoids from the basal Blue Lias Formation (Lower Jurassic) near Watchet, Somerset, England. Geoscience in South-West England 14: 161–166.
Palmer, C.P. 1972. The Lower Lias (Lower Jurassic) between Watchet and Lilstock in North Somerset (United Kingdom). Newsletters on Stratigraphy 2: 1–30. Crossref
Pollard, J.E. 1968. The gastric contents of an ichthyosaur from the Lower Lias of Lyme Regis, Dorset. Palaeontology 11: 376–388.
Pollard, J.E. 1990. Evidence for diet. In: Briggs, D.E.G. and Crowther, P.R. (eds.), Palaeobiology a Synthesis, 363–367. Blackwell Scientific Publications, Oxford.
Reitner, J. 2009. Preserved gill remains in Phragmoteuthis conocauda (Quenstedt, 1846–49) (Toarcian, Southern Western Germany). Berliner paläobiologische Abhandlungen 10: 289–295.
Reitner, J. and Urlichs, M. 1983. Echte Weichteilbelemniten aus dem Untertoarcium (Posidonienschiefer) Südwestdeutschlands. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 165: 450–465. Crossref
Riegraf, W. 1982. New Coleoidea from the Lower Jurassic of southwest Germany. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1982 (2): 91–97. Crossref
Riegraf, W. 1996. Belemniten-Fanghäkchen (Cephalopoda, Coleoidea) aus der Psilonotenbank (Unterer Jura, tiefstes Hettangium) von Südwestdeutschland. Stuttgarter Beiträge zur Naturkunde Serie B (Geologie und Paläontologie) 239: 1–38.
Riegraf, W. and Reitner, J. 1979. Die “Weichteilbelemniten” des Posidonienschiefers (Untertoarcium) von Holzmaden (Baden-Württemberg) sind Fälschungen. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1979 (5): 291–304.
Schweigert, G. 2018. Miscellanea aus dem Nusplinger Plattenkalk (Ober-Kimmeridgium, Schwäbische Alb). 19. Zwei Fallbeispiele besonderer Belemnitenerhaltung. Jahresberichte und Mitteilungen des Oberrheinischen. Geologischen Vereins 100: 509–516. Crossref
Seilacher, A. 1982a. Ammonite shells as habitats—floats or benthic islands? In: G. Einsele and A. Seilacher (eds.), Cyclic and Event Stratification, 504. Springer, Berlin. Crossref
Seilacher, A. 1982b. Ammonite shells as habitats in the Posidonia shales of Holzmaden—floats or benthic islands? Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1982 (2): 98–114. Crossref
Simms, M.J. 2004. The Wessex Basin (Dorset and central Somerset). In: M.J. Simms, N. Chidlaw, N. Morton, and K.N. Page (eds.), British Lower Jurassic Stratigraphy, 54–108. Geological Conservation Review Series, No. 30, Joint Nature Conservation Committee, Peterborough.
Stevens, K., Pohle, A., Hoffmann, R., and Immenhauser, A. 2023. Bayesian inference reveals a complex evolutionary history of belemnites. Palaeontologia Electronica 26: a13. Crossref
Tanner, A.R., Fuchs, D., Winkelmann, I.E., Gilbert, M.T.P., Pankey, M.S., Ribeiro, Â.M., Kocot, K.M., Halanych, K.M., Oakley, T.H., Da Fonseca, R.R., Pisani, D., and Vinther, J. 2017. Molecular clocks indicate turnover and diversification of modern coleoid cephalopods during the Mesozoic marine revolution. Proceedings of the Royal Society B: Biological Sciences 284: 20162818. Crossref
Warrington, G. and Ivimey-Cook, H.C. 1995. The Late Triassic and Early Jurassic of coastal sections in West Somerset and South and Mid-Glamorgan. In: P.D. Taylor (ed.), Field Geology of the British Jurassic, 9–30. Geological Society, London.
Weedon, G.P. 1986. Hemipelagic shelf sedimentation and climatic cycles: the basal Jurassic (Blue Lias) of South Britain. Earth and Planetary Science Letters 76: 321–335. Crossref
Weedon, G.P., Jenkyns, H.C., and Page, K.N. 2018. Combined sea-level and climate controls on limestone formation, hiatuses and ammonite preservation in the Blue Lias Formation, South Britain (uppermost Triassic–Lower Jurassic). Geological Magazine 155: 1117–1149. Crossref
Weedon, G.P., Page, K.N., and Jenkyns, H,C. 2017. Cyclostratigraphy, stratigraphic gaps and the duration of the Hettangian Stage (Jurassic): insights from the Blue Lias Formation of southern Britain. Geological Magazine 156: 1469–1509. Crossref
Weis, R. and Delsate, D. 2005. Presence de Belemnites precoces dans l’Hettangian de Belgique. Memoirs of the Geological Survey of Belgium 51: 27–31.
Weis, R. and Delsate, D. 2006. The earliest belemnites: new records from the Hettangian of Belgium and Luxembourg. Acta Universitatis Carolinae Geologica 24: 181–184.
Westermann, G.E.G. 1975. Architecture and buoyancy of simple cephalopod phragmocones and remarks on ammonites. Paläontologische Zeitschrift 49: 221–234. Crossref
Whitaker, A. and Green, G.W. 1983. Geology of the Country around Weston-super-Mare. Memoir for the 1:50 000 Geological Sheet 279, new series with parts of sheets 263 and 295. HMSO, London.
Zhu, K.-Y. and Bian, Z.-X. 1984. Sinobelemnitidae, a new family of Belemnitida from the Upper Triassic of Longmenshan, Sichuan. Acta Palaeontologica Sinica 23: 300–325.
Acta Palaeontol. Pol. 69 (3): 425–445, 2024
https://doi.org/10.4202/app.01172.2024