
Revision of the thylacocephalan biota from the Upper Triassic Polzberg Konservat-Lagerstätte, Austria
THOMAS LAVILLE, PETRA LUKENEDER, and ALEXANDER LUKENEDER
Laville, T., Lukeneder, P., and Lukeneder, A. 2024. Revision of the thylacocephalan biota from the Upper Triassic Polzberg Konservat-Lagerstätte, Austria. Acta Palaeontologica Polonica 69 (4): 713–730.
Known for over 140 years, the Upper Triassic Polzberg Konservat-Lagerstätte is an exceptional early Carnian marine assemblage discovered in the Northern Calcareous Alps of Lower Austria. The Polzberg biota is composed of a diverse marine fauna, including two species of thylacocephalans, an enigmatic group of fossil euarthropods that was highly diversified during the Triassic and thus representing a major component of many marine faunas throughout the Triassic. Recent excavations at the Polzberg lead to the discovery of new thylacocephalan specimens. In this study, we examine in detail this new thylacocephalan material as well as the type material. Four thylacocephalan taxa are now reported from the Polzberg Biota, making it one of the most diverse faunas from the Late Triassic. Paraostenia striata nov. comb., which was previously assigned to Austriocaris and later to Atropicaris, and Paraostenia cf. ambatolokobensis correspond to the first mention of Paraostenia in the Late Triassic. A new taxon, Atropicaris? sp, is reported for the first time from Polzberg. In addition, the presence of gills and muscles of the posterior trunk in Austriocaris carinata and Paraostenia cf. ambatolokobensis is the first trace of soft tissue preservation in Polzberg thylacocephalans. Additionally, we discuss the similarities of the Polzberg thylacocephalan fauna with other Triassic and Jurassic faunas. It shows great similarity with the Late Triassic Kozja dnina fauna, but also more surprisingly with the Middle Jurassic La Voulte fauna. This can be explained by the connection between the Neotethys and the Alpine Tethys during the Early Jurassic.
Key words: Arthropoda, Thylacocephala, Austria, Carnian Pluvial Episode, Palaeobiota, Polzberg Konservat-Lagerstätte, Triassic.
Thomas Laville [thomas.laville1@univ-tlse3.fr; ORCID: https://orcid.org/0000-0001-9081-7150 ], GET—Geosciences Environment Toulouse, Université Toulouse III—Paul Sabatier, 14 avenue Edouard Belin, 31400 Toulouse, France.
Petra Lukeneder [a0501032@unet.univie.ac.at; ORCID: https://orcid.org/0000-0003-4399-1214 ], Doctoral School of Ecology and Evolution, University of Vienna, Djerassiplatz 1, 1030 Vienna, Austria.
Alexander Lukeneder [alexander.lukeneder@nhm-wien.ac.at; ORCID: https://orcid.org/0000-0002-8384-3366 ], Department of Geology and Palaeontology, Natural History Museum Vienna, Burgring 7, 1010 Vienna, Austria.
Received 12 June 2024, accepted 15 October 2024, published online 27 December 2024.
Copyright © 2024 T. Laville et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Known for over 140 years (Stur 1874), the Upper Triassic Polzberg Konservat-Lagerstätte preserves an exceptional early Carnian marine assemblage discovered in the Northern Calcareous Alps of Lower Austria. Dated from the Carnian Julian 2 Ib, the Polzberg biota offers a unique insight into the evolution of marine ecosystems during one of the major geological Triassic events: the Carnian Pluvial Episode (CPE; Lukeneder et al. 2012, 2024; Ruffel et al. 2015; Dal Corso et al. 2020; Simms and Ruffel 2018). The CPE is characterised by a global warming and enhanced humidification during the early Carnian. This climate change led to an increase in siliciclastic influx towards the north-western branch of the Tethys, resulting in the deposition of the fossiliferous Reingraben formation at Polzberg.
The Polzberg biota is composed of a diverse marine fauna, with intermittent freshwater influences, as indicated by the presence of some freshwater inhabitants (Lukeneder and Lukeneder 2021). It yielded plant remains, bivalves, cephalopods, gastropods, polychaetes, arthropods, echinoids, vertebrate remains and coprolites, providing important new insights into Carnian food webs (Lukeneder and Lukeneder 2023).
Among arthropods, thylacocephalans are quite diverse and abundant in the Polzberg biota. Thylacocephalans are enigmatic fossil euarthropods characterised by a shield enveloping most of the body, hypertrophied compound eyes, three pairs of large raptorial appendages and a posterior trunk formed of eight to 22 elongated segments. Known at least from the Silurian (Haug et al. 2014) to the Cretaceous (e.g., Charbonnier et al. 2017), thylacocephalans were highly diversified during the Triassic, representing a major component of many marine faunas throughout the Triassic (e.g., Arduini and Brasca 1984; Dalla Vecchia 2012; Charbonnier et al. 2019; Ehiro et al. 2019; Ji et al. 2021; Laville et al. 2024). However, for the Carnian, the fossil record of thylacocephalans is very scarce, being restricted to the Kozja dnina Member, Slovenia, that yielded three taxa (Laville et al. 2024), and to the Polzberg Konservat-Lagerstätte (Glaessner 1931; Forchielli and Pervesler 2013). The Polzberg thylacocephalan fauna includes two species: Austriocaris carinata Glaessner, 1931, and Austriocaris striata Glaessner, 1931. The latter species has been the subject of many speculations. Since the 1980s, it has been successively reassigned to Ostenocaris (Arduini et al., 1984) by Briggs and Rolfe (1983), to Atropicaris Arduini & Brasca, 1984, by Schram (2014) and to Microcaris Pinna, 1974, by Ji et al. (2021). To date no definitive decision has been made regarding its systematic affinities and it is therefore necessary to revise this species.
The present study provides new insights into the Polzberg thylacocephalan fauna. We revised all known Polzberg thylacocephalans, including the type material and new findings made during recent excavations. Additionally, we discuss the similarities of the Polzberg thylacocephalan fauna with other Triassic and Jurassic faunas.
Institutional abbreviations.—GBA, Geological Survey Austria (GeoSphere Austria since 2023); MSNM, Museo di Storia Naturale di Milano, Italy; NHMW, Naturhistorisches Museum Wien, Austria.
Other abbreviations.—Aad, anterodorsal angle; Aav, anteroventral angle; Apd, posterodorsal angle; Apv, posteroventral angle; Ha, anterior height; Hmax, maximal height; Hp, posterior height; Lr, rostrum length; Ls, total shield length; Lw, shield length without the rostrum.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:0F3508F1-AB47-45F3-A62E-313F406814CE.
Geological setting
The Upper Triassic outcrops at Polzberg are located on the western slope of Mount Schindelberg (1066 m), north of the Ois River, 4 km northeast of Lunz am See in Lower Austria. The locality Schindelberg is synonymous with the locality Polzberg (= Pölzberg; Stur 1874, 1886; 1:50 000, geological map, sheet 71 Ybbsitz; Ruttner and Schnabel 1988, and sheet 72 Mariazell; Bauer and Schnabel 1997; Lukeneder and Lukeneder 2021; Fig. 1). The northernmost tectonic elements of the Northern Calcareous Alps (NCA, Bajuvaric Units) in Lower Austria are the Frankenfels Nappe, followed to the south by the Lunz Nappe. Within the Lunz Nappe, the Reifling Basin (Krystyn 1991; Lukeneder and Lukeneder 2021, 2024; Lukeneder et al. 2024)—a Late Triassic intraplatform basin—is located between Polzberg and Großreifling. The exact position of the fossiliferous localities in the southern area of the Lunz Nappe within the lower, fossiliferous part of the Reingraben formation was determined by GPS (global positioning system): N 47°53’4.98” and E 15°4’28.15”, town Gaming, federal district Scheibbs.
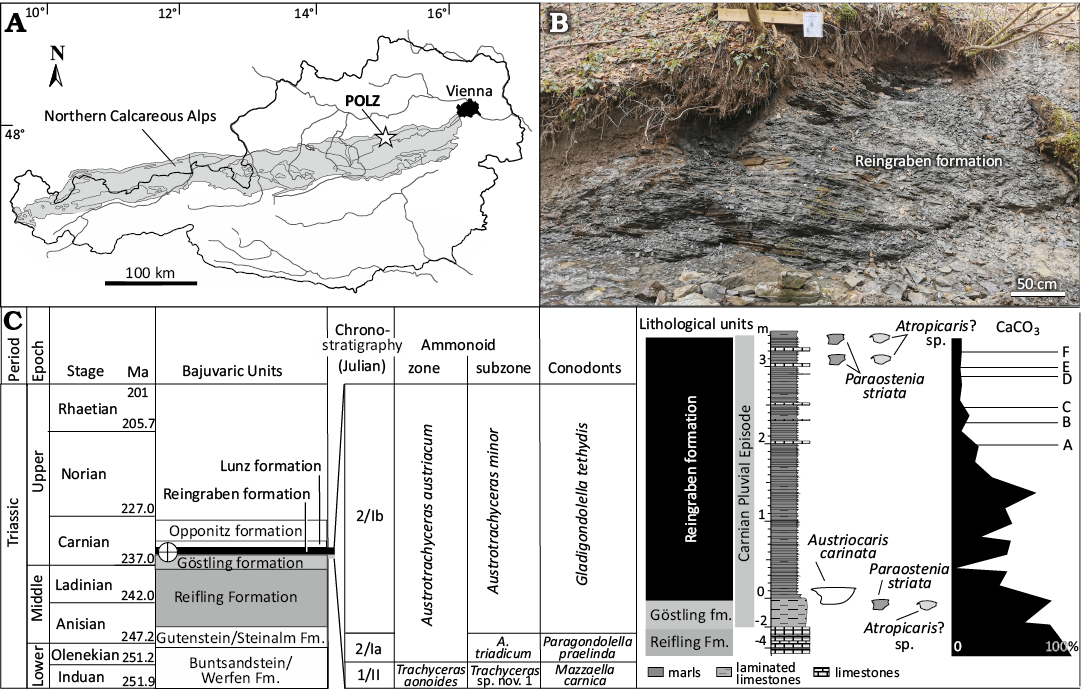
Fig. 1. Locality map of the Upper Triassic (lower Carnian) Polzberg area. A. Polzberg area (white star – POLZ) within the Northern Calcareous Alps (NCA), Lower Austria. B. Outcrop situation of the basal laminated Reingraben formation at Polzberg. C. Stratigraphic position of the lower Carnian Reingraben formation (cross-circle) at Polzberg with the layers comprising the Polzberg paleobiota within the Austrotrachyceras austriacum Zone. Log with indicated lithological units, thickness of the Polzberg section, CaCO3 content and calcareous dolostone-layers A–F; NCA area in gray. Adapted after Lukeneder and Lukeneder 2021 and Lukeneder et al. 2024. Abbreviations: A., Austrotrachyceras; Fm./fm., Formation/formation.
The deposits at Polzberg can be assigned to three units (Lukeneder et al. 2024), in stratigraphic order the Reifling Formation (formalized), the Göstling Formation (= Göstling limestones, Göstlinger Kalke; not formalized; frequently noted as Göstling Formation of the uppermost part of the Reifling Formation) and the Reingraben formation (= Reingraben Shales, Reingrabener Schiefer; not formalized). The thylacocephalans described herein derive from the basal three meters, calcareous to argillaceous of the Reingraben formation.
Material and methods
The present study is based on 72 thylacocephalan specimens housed in the Palaeontological collections of the Naturhistorisches Museum Wien, Austria. The material was collected over the last 140 years (field campaign GBA 1886 and NHMW 1909). Birgitt and Karl Aschauer (both Waidhofen an der Ybbs, Lower Austria) contributed with private collections from the last 20 years. Two of us (PL and AL) also contributed to these extensive collections with their own findings over the last 4 years. Rock matrix was removed and specimens were mechanically prepared using hammer, chisel and awls, subsequently using various types of pneumatic tools (e.g., air scribes).
Most specimens are compressed laterally. Ten specimens are preserved in butterfly configuration, suggesting they are possibly exuviae (see Laville et al. 2021). Five specimens show soft tissue preservation (GBA 2021/002/0010; NHMW 1901/0015/0005, 1901/0015/0009, 1901/0015/00010, 1901/0015/0045).
Specimens were photographed with a Nikon Digital Camera, D 5200 SLR, lens Micro SX SWM MICRO 1:1 Ø52 Nikon AF-S, processed by the free graphic software tool digiCamControl version V.2.1.2.0 at the NHMW. Digital high-quality photomicrographs were taken using a Discovery.V20 Stereo Zeiss microscope. The magnifications were ×10, ×20 and ×40 in incident light mode. Data from the AxioCam MRc5 Zeiss were processed and documented using the AxioVision SE64 Rel. 4.9 imaging system at the NHMW. In some photographs, specimens were whitened with ammonium chloride sublimate before photography to enhance details of ornamentation. Processing of pictures (histogram optimization, contrast, brightness) was made with GIMP 2.10 (GNU). Plates and line drawings were made with Inkscape 1.2 (GNU).
Measurements on the shield follow the scheme defined by Laville et al. (2021). They were taken directly from digital photographs using Fiji (GNU; Schindelin et al. 2012).
Systematic paleontology
Euarthropoda Lankester, 1904
Thylacocephala Pinna et al., 1982
Genus Atropicaris Arduini & Brasca, 1984
Type species: Atropicaris rostrata Arduini & Brasca, 1984, by original designation—Argilliti di Riva di Solto Formation, Sevatian, Norian, Upper Triassic; Ponte Giurino, Valle Imagna, Italy.
Included species: Atropicaris lintveri Laville et al., 2024, Anisian, Middle Triassic, Slovenia; Atropicaris aff. lintveri, Anisian, Middle Triassic, Slovenia (Laville et al. 2024); Atropicaris rostrata Arduini & Brasca, 1984, Upper Triassic, Italy; Atropicaris? sp., Anisian, Middle Triassic, China (Feldmann et al. 2015); Atropicaris? sp., Carnian, Upper Triassic, Austria (this study); Atropicaris sp., Ladinian, Middle Triassic, Switzerland (Bürgin et al. 1991); Atropicaris sp., Carnian, Upper Triassic, Slovenia (Laville et al. 2024).
Diagnosis (after Laville et al. 2024).—Thylacocephala with a triangular rostrum ending in a spatulate tip, a serrate dorsal midline, a concave posterior margin, a spiny posteroventral corner, and dorsal and ventral intercalary ridges.
Stratigraphic and geographic range.—Anisian to Carnian, Middle to Late Triassic; Asia (China) and Europe (Austria, Italy, Slovenia, Switzerland).
Atropicaris? sp.
Fig. 2.
Material.—24 specimens: NHMW 1910/0015/0033, 0035; NHMW 2021/0123/0059, 0767, 0769, 0776, 0777, 0779, 0780, 0782–0784, 0787, 0788, 0790, 0793, 0795; NHMW 2021/0123/0078, 0101–0106. All from Reingraben formation, Austrotrachyceras austriacum Zone, Austrotrachyceras minor Biohorizon, Julian 2 Ib, Carnian, Late Triassic; Polzberg near Lunz am See, Austria.
Dimensions.—Due to the deformation and distortion of the specimens, we only provide rough estimates for the length and maximal height of the shield: Ls >15.5 mm; Hmax ≈ 8.5 mm.
Description.—Shield morphology: Trapezoidal shield in lateral view, longer than high (Fig. 2). Maximal height reached around mid-length. Straight anterior margin slightly inclined posteroventrally (Fig. 2A, B). Anterodorsal corner not preserved. Convex dorsal midline, poorly preserved. Posterodorsal corner, posterior margin and posteroventral corner not preserved. V-shaped ventral margin consisting in an anterior part and a posterior one of equal length. Rounded anteroventral corner (Fig. 2B). Marginal fold delimited by a shallow groove at least near the ventral margin (Fig. 2B1).
Shield macro-ornamentation: Shield covered of at least 57 transverse sigmoid ridges, posteroventrally oriented with at least 49 primary ridges, three dorsal intercalary ridges and five ventral intercalary ridges (Fig. 2).
Other structures: Appendages, posterior trunk and soft parts not preserved.

Fig. 2. Thylacocephalan arthropod Atropicaris? sp. from the Upper Triassic Polzberg Konservat-Lagerstätte, Austria. A. NHMW 1910/0015/0035; A1, lateral view; A2, explanatory drawing. B. NHMW 2021/0124/0102; B1, left lateral view (ammonium chloride coating); B2, close-up of anteroventral corner (ammonium chloride coating). C. Left lateral view of NHMW 2021/0124/0101 (ammonium chloride coating). D. Right lateral NHMW 2021/0124/0103 (ammonium chloride coating).
Remarks.—Some of the specimens described here were previously assigned to Austriocaris striata (now Paraostenia striata comb. nov.; see below). However, their morphology differs from the one of the type material (and thus from Paraostenia). Even though all specimens possess transversal ridges, the specimens described above have a straight and anterior margin slightly inclined posteroventrally, without any optic notch, an anterior part of the ventral margin anterodorsally tilted and a straight posterior part of the ventral margin while the type material of P. striata has an anterior margin formed of a large optic notch, occupying the entire margin, a ventral margin with a horizontal anterior part and a slightly concave posterior part. Therefore, those specimens do not belong to P. striata.
One of the main features of our specimens is the presence of transverse ridges. This kind of ornamentation is well known in multiple thylacocephalans taxa such as species of Ankitokazocaris Arduini, 1990, Atropicaris, Concavicaris Rolfe, 1961, Ferrecaris Calzada and Mañé, 1993, Harrycaris Briggs and Rolfe, 1983, Kamnikaris Laville et al., 2024, Keelicaris Teruzzi & Charbonnier in Charbonnier et al., 2017, Microcaris Pinna, 1974, Paraconcavicaris Rak et al., 2018, and Thylacocephalus Lange et al., 2001.
The species described here differ from those of Ankitokazocaris, Concavicaris, Harrycaris, and Paraconcavicaris by the absence of an optic notch. Additionally, species of Paraconcavicaris has a ventral margin with a horizontal anterior part, longer than the posterior one. Our specimens have similarities with species of Kamnikaris, including a convex dorsal midline and a straight anterior margin, slightly tilted posteroventrally. However, they differ from this taxon by their pattern of macro-ornamentation: species of Kamnikaris have vertical arcuate ridges on its anterior part, and sigmoid ridges on its median and posterior parts while the studied specimens only has sigmoid ridges on the entire surface of its shield. Moreover, species of Kamnikaris have a ventral margin, which has an anterior part much shorter than the posterior one, while it is of equal length in our specimens. This is also the case for species of Thylacocephalus and Microcaris. Moreover, these of Microcaris are characterised by a straight dorsal midline and a horizontal anterior part of the ventral margin. The specimens described herein also differ from species of Thylacocephalus by the absence of spines on the anterior margin, of a dorsal carina and of numerous pores on the lateral surface of the shield. The morphology of species of Keelicaris is also quite different from the one of our specimen. In Keelicaris, the shield is keel-shaped with straight transverse ridges anteroventrally tilted, the anterior margin is made of an optic notch and the ventral margin is formed of a concave posterior part and a horizontal ventral part.
Species of Atropicaris and Ferrecaris display several similarities with our specimens: a straight anterior margin, slightly tilted posteroventrally, convex dorsal midline, v-shaped ventral margin with straight posterior and anterior parts of equal lengths. It is therefore most likely that our specimens belong to one of these taxa. Atropicaris and Ferrecaris differ by their rostrum morphology (spatulate tip for Atropicaris; sharp, triangular tip for Ferrecaris), the ornamentation of their dorsal midline (serrate vs. smooth), the morphology of their posterior margin (concave surrounded by a posterodorsal and a posteroventral spine vs. straight with rounded posterodorsal and posterodorsal corners). Unfortunately, the rostrum and the posterior part are not preserved in our specimens and the dorsal midline is only poorly preserved, not allowing the observation of the ornamentation. It is therefore almost impossible to assign to one or the other taxon. As it seems that only species of Atropicaris show ventral intercalary ridges, this feature could be useful to accurately identify our specimens. Thus, our specimens most probably belong to a species of Atropicaris. Nevertheless, in view of the poor preservation of the type material of Ferrecaris, the latter character should be taken with caution as it would be possible for such ridges to be observed if better preserved material was discovered. Based on those comparisons, we leave our specimens in open nomenclature, although probably belonging to a species of Atropicaris.
Genus Austriocaris Glaessner, 1931
Type species: Austriocaris carinata Glaessner, 1931, by original designation, Reingraben formation, Austrotrachyceras austriacum Zone, Austrotrachyceras minor Biohorizon, Julian 2 Ib, Carnian, Upper Triassic, Polzberg, Austria.
Included species: Austriocaris sp., Carnian, Upper Triassic, Slovenia (Laville et al. 2024); Austriocaris secretanae Laville et al., 2023, Callovian, Middle Jurassic, France.
Emended diagnosis.—Thylacocephala with an optic notch occupying only the ventral part of the anterior margin, presence of a straight dorsal midline and a sharp posterodorsal corner.
Remarks.—In their diagnosis of Austriocaris, Laville et al. (2023) erroneously mentioned the presence of a sharp posteroventral corner. In reality, it is the posterodorsal angle that is sharp and diagnostic for Austriocaris.
Stratigraphic and geographic range.—Carnian, Late Triassic of Austria.
Austriocaris carinata Glaessner, 1931
Figs. 3–5.
1931 Austriocaris carinata sp. nov.; Glaessner 1931: 483, text-figs. 5, 6.
1969 Austriocaris carinata Glaessner, 1931; Rolfe 1969: R317, fig. 140.2.
1983 Austriocaris carinata Glaessner, 1931; Briggs and Rolfe 1983: 270, text-fig. 6C.
1990 Austriocaris carinata Glaessner, 1931; Dalla Vecchia and Muscio 1990: 41.
2014 Austriocaris carinata Glaessner, 1931; Schram 2014: 349.
2017 Austriocaris carinata Glaessner, 1931; Ji et al. 2017: 175.
2018 Austriocaris carinata Glaessner, 1931; Rak et al. 2018: 278, table 1.
2019 Austriocaris carinata Glaessner, 1931; Charbonnier et al. 2019: table 1.
2021 Austriocaris carinata Glaessner, 1931; Ji et al. 2021: table 1.
2021 Austriocaris carinata Glaessner, 1931; Laville et al. 2021: table 1.
2021 Austriocaris carinata Glaessner, 1931; Lukeneder and Lukeneder 2021: fig. 3L, table 1.
2023 Austriocaris carinata Glaessner, 1931; Lukeneder and Lukeneder. 2023: fig. 3Q, table 1.
Holotype: NHMW 1910/0015/0041; sub-complete shield with well-preserved ornamentation.
Type locality: Polzberg near Lunz am See, Austria.
Type horizon: Reingraben formation, Austrotrachyceras austriacum Zone, Austrotrachyceras minor Biohorizon, Julian 2 Ib, Carnian, Upper Triassic.
Material.—Type material and 37 specimens: GBA 2021/ 002/0010, NHMW 1910/0015/0001–0017, 0037–0039, 0040, 0044–0050; NHMW 2021/0123/0002, 0203, 0764–0766; NHMW 2021/0124/0108–0110. All from type locality and horizon.
Dimensions.—See Table 1.
Table 1. Descriptive statistics of morphometric measurements. Abbreviations: N, number of specimens measured; Na, number of specimens for which a specific measurement has not been made; NA, non-applicable; SD, standard deviation.
|
Mesures |
Austriocaris
carinata |
Paraostenia
cf. ambatolokobensis |
Paraostenia
striata |
|
|
Total shield length (mm) |
mean (SD) |
86.45 (12.46) |
31.35 (NA) |
27.16 (1.31) |
|
minimum-maximum |
76.06–100.26 |
31.35–31.35 |
26.23–28.09 |
|
|
Na |
0 |
0 |
0 |
|
|
Shield length without the rostrum (mm) |
mean (SD) |
76.68 (11.04) |
NA (NA) |
NA (NA) |
|
minimum-maximum |
67.97–89.10 |
NA |
NA |
|
|
Na |
0 |
1 |
2 |
|
|
Rostrum length (mm) |
mean (SD) |
9.83 (1.53) |
NA (NA) |
NA (NA) |
|
minimum-maximum |
8.17–11.18 |
NA |
NA |
|
|
Na |
0 |
1 |
2 |
|
|
Anterior height (mm) |
mean (SD) |
29.17 (4.09) |
14.98 (NA) |
12.96 (0.59) |
|
minimum-maximum |
26.27–32.06 |
14.98–14.98 |
12.54–13.38 |
|
|
Na |
1 |
0 |
0 |
|
|
Maximal height (mm) |
mean (SD) |
NA (NA) |
19.36 (NA) |
15.96 (1.19) |
|
minimum-maximum |
NA |
19.36–19.36 |
15.12–16.80 |
|
|
Na |
3 |
0 |
0 |
|
|
Posterior height (mm) |
mean (SD) |
7.71 (2.37) |
4.56 (NA) |
6.23 (0.02) |
|
minimum-maximum |
6.04–9.39 |
4.56–4.56 |
6.22–6.25 |
|
|
Na |
1 |
0 |
0 |
|
|
Anterodorsal angle (°) |
mean (SD) |
61.15 (0.87) |
68.52 (NA) |
67.13 (1.94) |
|
minimum-maximum |
60.53–61.77 |
68.52–68.52 |
65.76–68.50 |
|
|
Na |
1 |
0 |
0 |
|
|
Anteroventral angle (°) |
mean (SD) |
82.28 (1.14) |
71.31 (NA) |
74.16 (4.02) |
|
minimum-maximum |
81.37–83.56 |
71.31–71.31 |
71.32–77.00 |
|
|
Na |
0 |
0 |
0 |
|
|
Posteroventral angle (°) |
mean (SD) |
145.76 (0.77) |
107.32 (NA) |
90.52 (0.67) |
|
minimum-maximum |
145.22–146.30 |
107.32–107.32 |
90.05–91.00 |
|
|
Na |
1 |
0 |
0 |
|
|
Posterodorsal angle (°) |
mean (SD) |
82.27 (1.61) |
97.70 (NA) |
76.20 (16.65) |
|
minimum-maximum |
80.56–83.76 |
97.70–97.70 |
64.43–87.98 |
|
|
Na |
0 |
0 |
0 |
|
|
Maximal height/total shield length ratio |
mean (SD) |
NA (NA) |
0.62 (NA) |
0.59 (0.02) |
|
minimum-maximum |
NA |
0.62–0.62 |
0.58–0.60 |
|
|
Na |
3 |
0 |
0 |
|
Description.—Shield morphology: Sub-trapezoidal shield in lateral view, much longer than high (Fig. 3A1, A3). Maximal height reached around mid-length. Anterior margin made of an optic notch in its ventral part, sinuous and posteroventrally oriented in its dorsal part (Fig. 3A–C). Anterodorsal corner expressed as a sharp, short triangular rostrum (Fig. 3B). Straight and horizontal dorsal midline, divided into two branches near the posterodorsal angle and thus delimiting this later (Fig. 4A1–A3). Posterodorsal corner expressed as a spine (Fig. 3D, E). Short, straight and vertical posterior margin. Ventral margin divided into a straight posterior part, anteroventrally oriented, and a straight horizontal anterior part (Fig. 3C). Anterior part longer than the posterior one. Both parts merging approximately at the posterior third of the length. Rounded anteroventral corner. Marginal fold delimited by a shallow groove near all free margins (Fig. 4D1, D2).

Fig. 3. Thylacocephalan arthropod Austriocaris carinata Glaessner, 1931, from the Upper Triassic Polzberg Lagerstätte, Austria. A. NHMW 1910/0015/0041 (holotype); A1, left lateral view; A2, left lateral view (ammonium chloride coating); A3, explanatory drawing. B. NHMW 1910/0015/0002; B1, left lateral view; B2, close-up of rostrum (ammonium chloride coating). C. NHMW 1910/0015/0014, left lateral view. D. NHMW 1910/0015/0001, left lateral view. E. NHMW 1910/0015/0039; E1, dorsal view; E2, dorsal view (ammonium chloride coating).

Fig. 4. Thylacocephalan arthropod Austriocaris carinata Glaessner, 1931, from the Upper Triassic Polzberg Lagerstätte, Austria. A. NHMW 1910/0015/0050; A1, dorsal view; A2, dorsal view (ammonium chloride coating); A3, close-up of posterodorsal corner; A4, close-up of ventrolateral carina. B. NHMW 2021/0123/0764a; B1, right lateral view; B2, close-up of dorsolateral part of the shield (ammonium chloride coating). C. NHMW 1910/0015/0044, right lateral view. D. NHMW 1910/0015/0045; D1, left lateral view; D2, left lateral view (ammonium chloride coating); D3, close-up of ventrolateral carina (ammonium chloride coating); D4, close-up of dorsolateral carina (ammonium chloride coating); D5, close-up of gills (ammonium chloride coating).
Shield macro-ornamentation: Shield with a straight dorsolateral carina, slightly tilted posteroventrally (Fig. 4D1, D2). Dorsolateral carina converging with the dorsal midline at about one fifth of the length. Dorsolateral carina adorned with at least 30 tubercles, which might be remains of spines. Mediolateral carina located in the posterior half of the shield (Fig. 3A). Mediolateral carina divided into a posterior half, anteroventrally tilted, and straight horizontal anterior half. C-shaped ventrolateral carina, located in the anterior half of the shield (Figs 3A, 4A1, A2, A4). Concavity of the carina directed posteriorly. In some specimens, ventrolateral carina associated with a large oval bulge near its dorsal tip (Fig. 4C). Short straight lateral carina dividing the rostrum into two parts (Fig. 4B). Fields of small punctuations visible between the anteroventral corner and the ventrolateral carina (Fig. 4D1, D3), and in the dorsal part of the shield, above the dorsolateral carina (Fig. 4B). High density of punctuations on the ventral branch of the ventrolateral carina, and on the part symmetrical to the ventral branch, in relation to the dorsal branch, giving the impression of having a third branch to the carina (Fig. 3A).
Posterior trunk: Posterior trunk made of at least five segments, visible by their lateral musculature in the posterior part of the shield (Fig. 5).
Gills: Set of four gills located in the central part of the shield. Gills with an elliptic shape, becoming arcuate near the dorsal midline (Figs. 4D5, 5A1–A3).
Other structures: Compound eyes and raptorial appendages not preserved.

Fig. 5. Thylacocephalan arthropod Austriocaris carinata Glaessner, 1931 from the Upper Triassic Polzberg Lagerstätte, Austria. A. NHMW 1910/0015/0009; A1, right lateral view; A2, right lateral view (ammonium chloride coating); A3, explanatory drawing; A4, close-up of posterior trunk (ammonium chloride coating). B. Right lateral view of NHMW 1910/0015/0005. C. Right lateral view of NHMW 1910/0015/0010.
Remarks.—In its original description of Austriocaris carinata, Glaessner (1931) had reversed the orientation of the specimens, the posterior part being at the front. This influenced his description, leading for example to describe the optical notch as a posterior notch, from which the abdomen came out. Rolfe (1969) and Forchielli and Pervesler (2013) later followed this orientation.
Since the original description, A. carinata has been recognised as a thylacocephalan, and new species of Austriocaris have been described (Laville et al. 2023, 2024). This allowed to clarify the orientation of Austriocaris.
Two other morphological characters have also been misidentified in the past. The first is the anterior part of the ventral margin. Indeed, this structure is actually not anteroventrally tilted, as depicted by Glaessner (1931) and Rolfe (1969), but it is horizontal, as shown in Fig. 3C. The second character is the presence of a rostral plate. The posterodorsal spine delimited by the dichotomy of the dorsal midline has been interpreted as a rostral plate based on the comparison with phyllocarids. However, this spine neither is a rostrum and nor is articulated (absence of hinge) and thus is not movable as in phyllocarids.
With this redescription, we also provide new details that were previously unknown such as the presence of tubercles on the dorsolateral carina. A tuberculate dorsolateral carina was already known in one species of Austriocaris, Austriocaris secretanae Laville et al., 2023. The most important character newly described is the soft-part-preservation. Indeed, several gills as well as the lateral musculature of some posterior trunk segments are reported here. This is the first record of soft part preservation in Polzberg thylacocephalans.
Stratigraphic and geographic range.—Carnian, Upper Triassic of Austria.
Genus Paraostenia Secrétan, 1985
Type species: Paraostenia voultensis Secrétan, 1985, by monotypy—gracilis Biozone, lower Callovian, Middle Jurassic; La Voulte-sur-Rhône, Ardèche, France.
Species included: Paraostenia ambatolokobensis (Arduini, 1990), Dienerian/Smithian, Lower Triassic, Madagascar; Paraostenia striata (Glaessner, 1931) comb. nov., Carnian, Upper Triassic, Austria; Paraostenia voultensis Secrétan, 1985, Callovian, Middle Jurassic, France; Paraostenia sp. Ehiro & Kato in Ehiro et al., 2015, Spathian, Lower Triassic, Japan.
Emended diagnosis (after Laville et al. 2023).—Thylacocephala with a symmetric optic notch, a sharp anterodorsal corner, a dorsal carina, a concave posterior margin, rounded posterodorsal and posteroventral corners, long and gracile appendages.
Stratigraphic and geographic range.—Dienerian to Callovian, Early Triassic to Middle Jurassic; Asia (Japan), Africa (Madagascar) and Europe (Austria, France).
Paraostenia striata (Glaessner, 1931) comb. nov.
Fig. 6.
1931 Austriocaris striata sp. nov.; Glaessner, 1931: 483, 484, text-figs. 7, 8.
1984 Ostenia striata (Glaessner, 1931); Briggs and Rolfe 1983: 270, text-fig. 6b.
1984 Austriocaris striata Glaessner, 1931; Arduini and Brasca 1984: 92.
1986 Austriocaris striata Glaessner, 1931; Tintori et al. 1986: 241.
1990 Austriocaris striata Glaessner, 1931; Dalla Vecchia and Muscio 1990: 41.
2014 Atropicaris striata (Glaessner, 1931); Schram 2014: 352.
2015 Ostenocaris striata (Glaessner, 1931); Ehiro et al. 2015: 279, table 1.
2019 Austriocaris striata Glaessner, 1931; Teruzzi and Muscio 2019: 54.
2019 Atropicaris striata (Glaessner, 1931); Charbonnier et al. 2019: table 1.
2021 Microcaris (Atropicaris) striata (Glaessner, 1931); Ji et al. 2021: 311, table 1.
2021 Atropicaris striata (Glaessner, 1931); Laville et al. 2021: table 1.
2021 Atropicaris striata (Glaessner, 1931); Lukeneder and Lukeneder 2021: fig. 3M, table 1.
2023 Atropicaris striata (Glaessner, 1931); Lukeneder and Lukeneder 2023: fig. 3P, table 1.
2024 Atropicaris striata (Glaessner, 1931); Lukeneder et al. 2024: 21.
Holotype: NHMW 1910/0015/0042, complete shield.
Type locality: Polzberg near Lunz am See, Austria.
Type horizon: Reingraben formation, Austrotrachyceras austriacum Zone, Austrotrachyceras minor Biohorizon, Julian 2 Ib, Carnian, Upper Triassic.
Diagnosis (this study).—Paraostenia with thin transversal and slightly sigmoid ridges on the shield.
Material.—Type material and eight specimens: NHMW 1910/0015/0034, 0036, 0043; NHMW 2021/0123/0770, 0772, 0791, 0796, 0797. All from the type locality and horizon.
Dimensions.—See Table 1.
Description.—Shield morphology: Curved pentagonal-shaped shield in lateral view, longer than large (Fig. 6A1–A3, B1–C2). Maximal height reached around mid-length. Anterior margin made of a well-developed, symmetric optic notch, occupying the entire margin (Fig. 6B). Sharp anterodorsal corner. Convex dorsal midline. Rounded posterodorsal and posteroventral corners. Short, concave posterior margin, posteroventrally tilted (Fig. 6A1–A3). Ventral margin made of a horizontal anterior part and a slightly concave posterior one, anteroventrally tilted; both parts having approximately the same length (Fig. 6A). Rounded anteroventral corner (Fig. 6C3). Marginal fold delimited by a shallow groove near all free margins (Fig. 6A4).
Shield macro-ornamentation: Shield with a comma-shaped ventrolateral furrow (Fig. 6A1–A3). Anterior part of the ventral margin adorned with at least 17 small tubercles near the anteroventral corner (Fig. 6C3). Shield covered of at least 140 thin slightly sigmoid transverse ridges, posteroventrally oriented in the first quarter of the shield, vertical in the second quarter; in the second half of the shield, ridges being vertical in the dorsal part and becoming anteroventrally tilted near the posterior part of the ventral margin (Fig. 6A1–A4).
Other structures: Appendages, posterior trunk and soft parts not preserved.

Fig. 6. Thylacocephalan arthropod Paraostenia striata (Glaessner, 1931) from the Upper Triassic Polzberg Lagerstätte, Austria. A. NHMW 1910/0015/0034; A1, left lateral view; A2, left lateral view, ammonium chloride coating; A3, explanatory drawing; A4, close-up of posteroventral corner, ammonium chloride coating. B. NHMW 1910/0015/0042 (holotype, counter-part); B1, dorsal view; B2, close-up of left side of the shield, ammonium chloride coating; B3, explanatory drawing. C. NHMW 1910/0015/0042 (holotype, part); C1, dorsal view; C2, close-up of right side of the shield, ammonium chloride coating; C3, close-up of anteroventral corner, ammonium chloride coating.
Remarks.—The species A. striata was first assigned by Glaesnner (1931) to Austriocaris. Many years later, Briggs and Rolfe (1983) noticed the differences between the specimens of A. striata and Austriocaris. Austriocaris has a rostrum, an optic notch restricted to the ventral part of the anterior margin, a straight dorsal midline, a spiny posterodorsal corner, a straight dorsal margin and a shield covered by three lateral carinae while A. striata does not have a rostrum and lateral carinae but have a convex dorsal midline, a rounded posterodorsal corner and a concave dorsal margin. Briggs and Rolfe (1983) thus decided to assign this species to Ostenocaris Arduini et al., 1984 (known as Ostenia at that time). However, A. striata differs from Ostenocaris species by the morphologies of the anterior margin (optic notch in A. striata, sinuous and proverse in Ostenocaris) and of the posterior margin (concave vs. straight). The confusion of Briggs and Rolfe (1983) might come from the fact that in the original description of Ostenocaris cypriformis Arduini et al., 1984, the type species of Ostenocaris, some specimens of Paraostenia have been assigned, provoking many misidentifications as explained by Laville et al. (2021).
A year after Briggs and Rolfe (1983), Arduini and Brasca (1984) reassigned A. striata to Atropicaris based mostly on the presence of ridges and of a rostrum. This choice was later followed by most authors (e.g., Dalla Vecchia and Muscio 1990; Schram 2014; Ehiro et al. 2015; Ji et al. 2021 as Microcaris (Atropicaris) striata; Laville et al. 2021). As explained above for Atropicaris? sp., several specimens assigned to A. striata actually have a morphology quite different from the holotype, and are most likely belonging to Atropicaris. However, the holotype and additional specimens mentioned here differ from Atropicaris species especially by the absence of a rostrum, by the presence of an optic notch and by the horizontal morphology of the anterior part of the ventral margin.
Actually, A. striata possesses the typical characters of Paraostenia Secrétan, 1985: a large optic notch, a sharp anterodorsal corner, a rounded anteroventral corner, a concave posterior margin, a rounded posterodorsal corner and a horizontal anterior part of the ventral margin. A. striata should be assigned to Paraostenia. We therefore propose a new combination Paraostenia striata comb. nov. for this species.
Paraostenia striata display a unique character among Paraostenia: the thin transversal ridges. Even though shield macro-ornamentation is well-known in other Paraostenia species (horseshoe-like ornamentation in Paraostenia voultensis Secrétan, 1985; longitudinal ridges in Paraostenia ambatolokobensis [Arduini, 1990]), thin transversal ridges are only known in P. striata.
In addition, P. striata does not carry any lateral or dorsal carina, unlike other species. Nevertheless, due to the poor preservation of the dorsal midline or to the butterfly configuration of the specimens, it is difficult to accurately determine the presence or absence of a dorsal carina.
Stratigraphic and geographic range.—Carnian, Upper Triassic of Austria.
Paraostenia cf. ambatolokobensis (Arduini, 1990)
Fig. 7.
Material.—NHMW 2024/0201/00012 from the Reingraben formation, Austrotrachyceras austriacum Zone, Austrotrachyceras minor Biohorizon, Julian 2 Ib, Carnian, Upper Triassic; Polzberg near Lunz am See, Austria.
Dimensions.—See Table 1.
Description.—Shield morphology: Curved hexagonal-shaped shield in lateral view, longer than large (Fig. 7A1, A2). Maximal height reached around mid-length. Anterior margin made of a well-developed, symmetric optic notch, occupying the entire margin. Anterodorsal corner not preserved. Convex dorsal midline with a large fin-like carina (Fig. 7A3). Rounded posterodorsal corner. Short, concave posterior margin. Rounded posteroventral corner. Ventral margin made of a horizontal anterior part and a slightly concave posterior one, anteroventrally tilted; both parts having approximately the same length (Fig. 7A1, A2). Rounded anteroventral corner. Marginal fold delimited by a shallow groove near anterior and ventral margin (Fig. 7A4).
Shield macro-ornamentation: Shield with a comma-shaped ventrolateral furrow, being most probably a muscle scar (Fig. 7A1, A2). Long dorsolateral carina running along almost the entire length of the shield. Shield covered of short, transversal ridges. Ridges having a comma-like shape below the dorsal carina and near the anterodorsal and anteroventral corners (Fig. 7A3), and a scale-like shape on the rest of the shield (Fig. 7A4).
Posterior trunk: Posterior trunk made of at least four segments (Fig. 7A1, A2).
Gills: Set of eight gills located in the central part of the shield. Gills with an elliptic shape, becoming arcuate near the dorsal midline (Fig. 7A1, A2).

Fig. 7. Comparison of thylacocephalan arthropods Paraostenia cf. ambatolokobensis (Arduini, 1990), from the Upper Triassic Polzberg Lagerstätte, Austria (A) and Paraostenia ambatolokobensis (Arduini, 1990) from the Upper Triassic, Madagascar (B). A. NHMW 2024/0201/0001; A1, left lateral view; A2, explanatory drawing; A3, close-up of dorsal carina; A4, close-up of ventral margin. B. MSNM i27655 a/b, right lateral view.
Remarks.—The specimen described above have key features of Paraostenia Secrétan, 1985: a large optic notch, a dorsal carina, a concave posterior margin, rounded posterodorsal and posteroventral corners and a horizontal anterior part of the ventral margin. It differs from the other Paraostenia specimens from Polzberg by the presence of a dorsal carina and by its pattern of macro-ornamentation (scale-like ridges vs long, transverse ridges). In terms of size, the specimen is a bit longer and taller than Paraostenia striata and the Hmax/Ls ratios are quite similar (see Table 1). The specimen resembles the specimens of Paraostenia ambatolokobensis from the Lower Triassic of Madagascar (Arduini 1990). Indeed, they both have a dorsal carina and a dorsolateral carina but most importantly they share a unique pattern of macro-ornamentation among species of Paraostenia: the presence of short transverse comma-like ridges on the dorsal area of the shield, and the presence of scale-like ridges on the rest of the shield (see Fig. 7B for P. ambatolokobensis). This pattern has not been described in any other Paraostenia species. It is therefore most probable that this specimen belongs to P. ambatolokobensis. However, due to the poor preservation of some morphological features (e.g., anterodorsal corner) in our specimens and to the fact that only one specimen of this taxon has been recovered from the Polzberg Lagerstätte, we refrain to assign this specimen to a particular species and let it in open nomenclature.
Discussion
Oval bulge in Austriocaris carinata: a trace of parasitism?—Glaessner (1931) mentioned the fact that the oval bulge associated with the mediolateral carina is not preserved in several specimens of A. carinata, as it disappeared under pressure. We do not concur with this explanation. Indeed, among the specimens that we studied, some are well-preserved and do not show any deformation or wrinkling. Moreover, in those specimens (e.g., NHMW 1910/0015/0041, Fig. 3A; NHWM 1910/0015/0045, Fig. 4D1–D4), some structures are preserved in 3D such as the carinae. Therefore, if present, this bulge should have been preserved in those specimens.
Another hypothesis could explain the presence/absence of the oval bulge: parasitism. Indeed, the presence of parasites in pancrustaceans is far from new, whether in extant (e.g., O’Brien and Van Wyk 1985; Williams and Boyko 2012) or fossil specimens (see Klompmarker and Boxshall 2015 for a review). In pancrustaceans, some parasites may cause swelling or cysts in the shield of some hosts (McDermott 1991; Klompmaker et al. 2014). These bulges are generally located near the branchial region, as is the case for some of our specimens. In fossil pancrustaceans (mainly decapods), the origin of these swellings has generally been attributed to bopyrid isopods, although no fossils from this group have been found so far. It is therefore possible that a similar phenomenon affected some of the individuals of A. carinata. In order to confirm or not the possible parasitism, it would be necessary to find the parasite itself. Pending such confirmation, this hypothesis cannot be ruled out for the time being.
Diversity and palaeogeographic distribution.—We report four thylacocephalan taxa from the Polzberg Konservat-Lagerstätte. Two were already known (Austriocaris carinata and Paraostenia striata comb. nov.; Glaessner 1931; Forchielli and Pervesler 2013) and two are here reported for the first time as part of the Polzberg biota (Atropicaris? sp.; Paraostenia cf. ambatolokobensis). The diversity of the Polzberg thylacocephalan biota is, with four taxa, relatively high in comparison to most other Late Triassic thylaccocephalan faunas. Indeed, other Late Triassic thylacocephalan-bearing formations have low diversities, with only one known taxon (Pinna 1974, 1976; Arduini and Brasca 1984; Dalla Vecchia 1993), with the exception of the Forni Dolomites Formation (Norian, Upper Triassic; Italy; Dalla Vecchia 2012) and the Martuljek Limestone Formation (Carnian, Upper Triassic; Slovenia; Laville et al. 2024), which have a taxonomic diversity close to the Polzberg biota, with three taxa.
Until now, Austriocaris was the only known taxon described from the Polzberg biota. Since then, it has been reported from two other deposits: Kozja dnina, Slovenia (Laville et al. 2024) and La Voulte-sur-Rhône Lagerstätte (Laville et al. 2023). Atropicaris is a well-known taxon from the Middle (China: Feldmann et al. 2015; Slovenia: Laville et al. 2024; Switzerland: Bürgin et al. 1991) and Upper Triassic (Italy: Arduini and Brasca 1984; Dalla Vecchia and Muscio 1990; Dalla Vecchia 2012; Slovenia: Laville et al. 2024) and it is therefore not surprising to find this taxon in Polzberg. Paraostenia is known from the Lower Triassic (Japan: Ehiro et al. 2015; Madagascar: Laville et al. 2021) and from the Lower and Middle Jurassic (France: Secrétan 1985; Italy: Arduini et al. 1980) but not from the Middle and Upper Triassic. Therefore, Paraostenia striata and Paraostenia cf. ambatolokobensis from the Polzberg Konservat-Lagerstätte corresponds to the first mention of this taxon from the Upper Triassic.
Interestingly, the Polzberg thylacocephalan fauna displays particular affinities with two other faunas: one from the Late Triassic, the Kozja dnina biota, another one from the Middle Jurassic, the La Voulte-sur-Rhône Lagerstätte. The Kozja dnina biota has a similar diversity as the Polzberg biota, with three thylacocephalan taxa reported, two of which are also known in the Polzberg: Austriocaris and Atropicaris (Laville et al. 2024). The La Voulte Lagerstätte has a much greater diversity than the Polzberg Konservat-Lagerstätte, with seven taxa described (Laville et al. 2023). Nevertheless, two of these taxa are also known in the Polzberg: Austriocaris and Paraostenia. The similarity of the Polzberg thylacocephalan fauna with the Kozja dnina biota is not surprising. Both deposits are of Carnian age (Polzberg: Julian; Kozja dnina: Tuvalian). They are both located in the Meliata-Maliac Ocean (Stampfli et al. 2002), even though they belong to two different branches of the ocean (Meliata for the Polzberg, Maliac for Kozja dnina). Finally, they were both formed in a similar environment (deep-water setting within an intraplatform basin; see Lukeneder and Lukeneder [2021] for the Polzberg and Celarc and Kolar-Jurkovšek [2008] for Kozja dnina). On the other hand, the similarity with the La Voulte Lagerstätte is much more surprising. The age difference is about 60 Ma and they were belonging to two different oceans (Meliata-Maliac Ocean for Polzberg, Alpine Tethys for the La Voulte Lagerstätte). However, these two oceans were probably connected during the Early Jurassic, through the Neotethys (Frizon de Lamotte et al. 2015). It probably allowed the dispersal of the fauna of the Neotethys and thus of the Meliata-Maliac ocean, towards the Alpine Tethys. Moreover, both faunas were located in deep environments, close to the slope-basin transition (Charbonnier et al. 2014). All these factors can explain the similarity between both faunas.
Conclusions
The revision of thylacocephalans from the Polzberg Konservat-Lagerstätte made it possible to study its diversity in detail. Four thylacocephalans taxa are now reported from Polzberg, making it one the most diverse Late Triassic faunas. Paraostenia striata nov. comb., which was previously assigned to Austriocaris or Atropicaris, and Paraostenia cf. ambatolokobensis correspond to the first mention of Paraostenia in the Late Triassic. A new taxon, Atropicaris? sp. is reported for the first time from Polzberg. In addition, the presence of gills and muscles of the posterior trunk in Austriocaris carinata and in Paraostenia cf. ambatolokobensis are the first traces of soft part preservation in Polzberg thylacocephalans. Finally, comparisons with other Jurassic but also Triassic thylacocephalan faunas show the similarity of the Polzberg thylacocephalan fauna with the Late Triassic Kozja dnina fauna, and more surprisingly with the Middle Jurassic La Voulte fauna.
Acknowledgments
We acknowledge the Federal Government of Lower Austria (project K3-F-964/001-2020 and K3-F-964/001-2023, Department Science and Research; headquarters in St. Pölten) and the association of the Freunde des Naturhistorischen Museums Wien (Project Trias Lukeneder) for the financial support. We thank the owners of the outcrop area, Franziska and Hermann Hofreiter, Eva and Karl Jagersberger (all Gaming, Austria), for sampling and digging permission. Manuel Andreas Staggl (University of Vienna, Austria) is thanked for providing the specimen of Paraostenia cf. ambatolokobensis. The manuscript greatly benefited from valuable comments of the in-house editoral team and the handling editor, Andrzej Kaim, and of two anonymous reviewers.
References
Arduini, P. 1990. Studies on Permo-Trias of Madagascar. 1. Thylacocephala from the Lower Trias of Madagascar. Atti della Società italiana di Scienze naturali e del Museo civico di Storia naturale di Milano 131: 197–204.
Arduini, P. and Brasca, A. 1984. Atropicaris: nuovo genere della classe Thylacocephala. Atti della Società italiana di Scienze naturali e del Museo civico di Storia naturale di Milano 125: 87–93.
Arduini, P., Pinna, G., and Teruzzi, G. 1980. A new and unusual Lower Jurassic cirriped from Osteno in Lombardy: Ostenia cypriformis n. g. n. sp. Atti della Società italiana di scienze naturali e del museo civico di storia naturale di Milano 121: 360–370.
Arduini, P., Pinna, G., and Teruzzi, G. 1984. Ostenocaris nom. nov. pro Ostenia Arduini, Pinna e Teruzzi, 1980. Atti della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano 125 (1–2): 48.
Bauer, F.K. and Schnabel, W. 1997. Geologische Karte der Republik Österreich. Sheet Mariazell 72, 1:50.000. Geologische Bundesanstalt.
Briggs, D.E.G. and Rolfe, W.D.I. 1983. New Concavicarida (new order:? Crustacea) from the Upper Devonian of Gogo, Western Australia, and the palaeoecology and affinities of the group. Special Papers in Palaeontology 30: 249–276.
Bürgin, T., Furrer, H., and Eichenberger, U. 1991. Die Prosanto-Formation: eine fischreiche Fossil-Lagerstätte in der Mitteltrias der Silvretta-Decke (Kanton Graubünden, Schweiz). Eclogae Geologicae Helvetiae 84: 921–990.
Calzada, S. and Mañé, R. 1993. Primera cita de un Tilacocéfalo (Crustacea) (Ferrecaris n. gen. magransi n. sp.) en el Ladiniense español. Trabajos Del Museo Geológico Del Seminario 246: 12–16.
Celarc, B. and Kolar-Jurkovšek, T. 2008. The Carnian–Norian basin-platform system of the Martuljek Mountain Group (Julian Alps, Slovenia): progradation of the Dachstein carbonate platform. Geologia Carpathica 59: 211–224.
Charbonnier, S., Audo, D., Caze, B., and Biot, V. 2014. The La Voulte-sur-Rhône Lagerstätte (Middle Jurassic, France). Comptes Rendus Palevol 13: 369–381. Crossref
Charbonnier, S., Brayard, A., and The Paris Biota Team. 2019. New thylacocephalans from the Early Triassic Paris Biota (Bear Lake County, Idaho, USA). Geobios 54: 37–43. Crossref
Charbonnier, S., Teruzzi, G., Audo, D., Lasseron, M., Haug, C., and Haug, J.T. 2017. New thylacocephalans from the Cretaceous Lagerstätten of Lebanon. Bulletin de la Société géologique de France 188: 19. Crossref
Dal Corso, J., Bernardi, M., Sun, Y., Song, H., Seyfullah, L.J., Preto, N., Gianolla, P., Ruffell, A., Kustatscher, E., Roghi, G., Merico, A., Hohn, S., Schmidt, A.R., Marzoli, A., Newton, R.J., Wignall, P.B., and Benton, M.J. 2020. Extinction and dawn of the modern world in the Carnian (Late Triassic). Science Advances 6: article eaba0099. Crossref
Dalla Vecchia, F.M. 1993. Segnalazione di crostacei nell’Unità Fonte Santa (Triassico Sup.) presso Filettino (Lazio, Italia). Gortania 14: 59–69.
Dalla Vecchia, F.M. 2012. Il Friuli 215 milioni di anni fa. Gli straordinari fossili di Preone: finestra su di un mondo scomparso. 224 pp. Comune di Preone, Preone.
Dalla Vecchia, F.M. and Muscio, G. 1990. Occurrence of Thylacocephala (Arthropoda, Crustacea) from the Upper Triassic of Carnic Prealps (NE Italy). Bollettino della Società-Paleontologica Italiana 29: 39–42.
Ehiro, M., Sasaki, O., Kano, H., and Nagase, T. 2019. Additional thylacocephalans (Arthropoda) from the Lower Triassic (upper Olenekian) Osawa Formation of the South Kitakami Belt, Northeast Japan. Palaeoworld 28: 320–333. Crossref
Ehiro, M., Sasaki, O., Kano, H., Nemoto, J., and Kato, H. 2015. Thylacocephala (Arthropoda) from the Lower Triassic of the South Kitakami Belt, Northeast Japan. Paleontological Research 19: 269–282. Crossref
Feldmann, R.M., Schweitzer, C.E., Hu, S., Huang, J., Zhou, C., Zhang, Q., Wen, W., Xie, T., and Maguire, E. 2015. Spatial distribution of Crustacea and associated organisms in the Luoping Biota (Anisian, Middle Triassic), Yunnan Province, China: evidence of periodic mass kills. Journal of Paleontology 89: 1022–1037. Crossref
Forchielli, A. and Pervesler, P. 2013. Phosphatic cuticle in thylacocephalans: a taphonomic case study of Austriocaris (Arthropoda, Thylacocephala) from the Fossil-Lagerstätte Polzberg (Reingraben shales, Carnian, Upper Triassic, Lower Austria). Austrian Journal of Earth Sciences 106: 46–61.
Frizon de Lamotte, D., Fourdan, B., Leleu, S., Leparmentier, F., and de Clarens, P. 2015. Style of rifting and the stages of Pangea breakup. Tectonics 34: 1009–1029. Crossref
Glaessner, M.F. 1931. Eine Crustaceenfauna aus den Lunzer Schichten Niederösterreichs. Jahrbuch der Geologischen Bundesanstalt 81: 467–486.
Haug, C., Briggs, D.E.G., Mikulic, D.G., Kluessendorf, J., and Haug, J.T. 2014. The implications of a Silurian and other thylacocephalan crustaceans for the functional morphology and systematic affinities of the group. BMC Evolutionary Biology 14: article 159. Crossref
Ji, C., Tintori, A., Jiang, D., and Motani, R. 2017. New species of Thylacocephala (Arthropoda) from the Spathian (Lower Triassic) of Chaohu, Anhui Province of China. PalZ 91 (2): 171–184. Crossref
Ji, C., Tintori, A., Jiang, D., Motani, R., and Confortini, F. 2021. New Thylacocephala (Crustacea) assemblage from the Spathian (Lower Triassic) of Majiashan (Chaohu, Anhui Province, South China). Journal of Paleontology 95: 305–319. Crossref
Klompmaker, A.A. and Boxshall, G.A. 2015. Fossil crustaceans as parasites and hosts. In: K. De Baets and D.T.J. Littlewood (eds.), Advances in Parasitology, Vol. 90, 233–289. Academic Press, Amsterdam. Crossref
Klompmaker, A.A., Artal, P., Bakel, B.W.M. van, Fraaije, R.H.B., and Jagt, J.W.M. 2014. Parasites in the fossil record: A Cretaceous fauna with isopod-infested decapod crustaceans, infestation patterns through time, and a new ichnotaxon. PLOS ONE 9: article e92551. Crossref
Krystyn, L. 1991. Die Fossillagerstätten der alpinen Trias. In: D. Nagel and G. Rabeder (eds.), Exkursionen im Jungpaläozoikum und Mesozoikum Österreichs, 24–78. Österreichische Paläontologische Gesellschaft Wien, Wien.
Lange, S., Hof, C.H.J., Schram, F.R., and Steeman, F.A. 2001. New genus and species from the Cretaceous of Lebanon links the Thylacocephala to the Crustacea. Palaeontology 44: 905–912. Crossref
Lankester, E. 1904. The structure and classification of the Arthropoda. Journal of Cell Science 2: 523–582. Crossref
Laville, T., Forel, M.-B., and Charbonnier, S. 2023. Re-appraisal of thylacocephalans (Euarthropoda, Thylacocephala) from the Jurassic La Voulte-sur-Rhône Lagerstätte. European Journal of Taxonomy 898: 1–61. Crossref
Laville, T., Hitij, T., Forel, M.-B., Gašparič, R., Žalohar, J., and Charbonnier, S. 2024. Middle and Late Triassic thylacocephalans from Slovenia: implications for their diversification. Bulletin of Geosciences 99: 219–253. Crossref
Laville, T., Smith, C.P.A., Forel, M.-B., Brayard, A., and Charbonnier, S. 2021. Review of Early Triassic Thylacocephala. Rivista Italiana di Paleontologia e Stratigrafia 127: 73–101.
Lukeneder, A. and Lukeneder, P. 2021. The Upper Triassic Polzberg palaeobiota from a marine Konservat-Lagerstätte deposited during the Carnian Pluvial Episode in Austria. Scientific Reports 11: article 16644. Crossref
Lukeneder, A. and Lukeneder, P. 2023. New data on the marine Upper Triassic palaeobiota from the Polzberg Konservat-Lagerstätte in Austria. Swiss Journal of Palaeontology 142: article 9. Crossref
Lukeneder, A. and Lukeneder, P. 2024. Ammonoid taxonomy of the Carnian Polzberg Konservat-Lagerstätte in Austria. Jahrbuch der Geologischen Bundesanstalt 162: 27–55.
Lukeneder, S., Lukeneder, A., Harzhauser, M., İslamoğlu, Y., Krystyn, L.,, and Lein, R. 2012. A delayed carbonate factory breakdown during the Tethyan-wide Carnian Pluvial Episode along the Cimmerian terranes (Taurus, Turkey). Facies 58: 279–296. Crossref
Lukeneder, A, Lukeneder P., Sachsenhofer, R., Roghi, G., and Rigo. M. 2024. Multi-proxy record of the Austrian Upper Triassic Polzberg Konservat-Lagerstätte in light of the Carnian Pluvial Episode. Scientific Reports 14: article 11194. Crossref
McDermott, J.J. 1991. Incidence and host-parasite relationship of Leidya bimini (Crustacea, Isopoda, Bopyridae) in the brachyuran crab Pachygrapsus transversus from Bermuda. Ophelia 33: 71–95. Crossref
O’Brien, J. and Van Wyk, P. 1985. Effects of crustacean parasitic castrators (epicaridean isopods and rhizocephalan barnacles) on growth of crustacean hosts. In: A.M. Wenner (ed.), Crustacean Issues 3: Factors in Adult Growth, 191–218. Balkema, Rotterdam.
Pinna, G. 1974. l crostacei della fauna triassica di Cene in Val Seriana (Bergamo). Memorie della Società italiana di scienze naturali in Milano 21: 5–34.
Pinna, G. 1976. I crostacei Triassici dell’alta Valvestino (Brescia). Natura Bresciana 13: 33–42.
Pinna, G., Arduini, P., Pesarini, C., and Teruzzi, G. 1982. Thylacocephala: una nuova classe di crostacei fossili. Atti della Società italiana di Scienze naturali e del Museo civico di Storia naturale di Milano 123: 469–482.
Rak, Š., Broda, K., and Kumpan, T. 2018. First Carboniferous thylacocephalan from Europe and its significance for the understanding of functional morphology of Concavicarididae Schram, 2014. Crustaceana 91: 265–285. Crossref
Rolfe, W.D.I. 1961. Concavicaris and Quasicaris, substitute names for Colpocaris Meek, 1872, and Pterocaris Barrande, 1872. Journal of Paleontology 35: 1243–1243.
Rolfe, W.D.I. 1969. Phyllocarida. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology, Part R, Arthropoda 4, Vol. 1, R296–R331, Geological Society of America and University of Kansas Press, Lawrence.
Ruffell, A., Simms, M.J., and Wignall, P.B. 2015. The Carnian Humid Episode of the late Triassic: A review. Geological Magazine 153: 271–284. Crossref
Ruttner, A. and Schnabel, W. 1988. Geologische Karte der Republik Österreich. Sheet Ybbsitz 71, 1:50.000. Geologische Bundesanstalt.
Schindelin, J., Arganda-Carreras, I., Frise, E., Kaynig, V., Longair, M., Pietzsch, T., Preibisch, S., Rueden, C., Saalfeld, S., Schmid, B., Tinevez, J.-Y., White, D.J., Hartenstein, V., Eliceiri, K., Tomancak, P., and Cardona, A. 2012. Fiji: an open-source platform for biological-image analysis. Nature Methods 9: 676–682. Crossref
Schram, F.R. 2014. Family level classification within Thylacocephala, with comments on their evolution and possible relationships. Crustaceana 87: 340–363. Crossref
Secrétan, S. 1985. Conchyliocarida, a class of fossil crustaceans: relationships to Malacostraca and postulated behaviour. Transactions of the Royal Society of Edinburgh: Earth Sciences 76: 381–389. Crossref
Simms, M.J. and Ruffell, A.H. 2018. The Carnian Pluvial Episode: from discovery, through obscurity, to acceptance. Journal of the Geological Society 175: 989–992. Crossref
Stampfli, G.M., Borel, G.D., Marchant, R., and Mosar, J. 2002. Western Alps geological constraints on western Tethyan reconstructions. Journal of the Virtual Explorer 8: 77–106. Crossref
Stur, D. 1874. Neue Aufschlüsse im Lunzer Sandsteine bei Lunz und ein neuer Fundort von Wengerschiefer im Pölzberg zwischen Lunzersee und Gaming. Verhandlungen der kaiserlich königlichen Geologischen Reichsanstalt 1: 271–273.
Stur, D. 1886. Vorlage des ersten fossilen Schädels von Ceratodus aus den ober triadischen Reingrabner Schiefern von Pölzberg nördlich bei Lunz. Verhandlungen der kaiserlich königlichen Geologischen Reichsanstalt 15: 381–383.
Teruzzi, G. and Muscio, G. 2019. Thylacocephalans from the Anisian (Middle Triassic) of the Carnic Alps. Gortania 40: 49–55.
Tintori, A., Bigi, E., Crugnola, G., and Danini, G. 1986. A new Jurassic Thylacocephala Rugocaris indunensis gen. n. sp. n. and its paleoecological significance. Rivista Italiana di Paleontologia e Stratigrafia 92: 239–250.
Williams, J.D. and Boyko, C.B. 2012. The global diversity of parasitic isopods associated with crustacean hosts (Isopoda: Bopyroidea and Cryptoniscoidea). PLoS One 7: e35350. Crossref
Acta Palaeontol. Pol. 69 (4): 713–730, 2024
https://doi.org/10.4202/app.1180.2024
Appendix 1
List of studied specimens.
|
Repository number |
Specimen number |
Identification |
Layer |
Observations |
|
GBA 2021/002/0010 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0001 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0002 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0003 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0004 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0005 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0006 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0007 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0008 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0009 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0010 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0011 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0012 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0013 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0014 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0015 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0016 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0017 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0033 |
|
Atropicaris? sp. |
|
|
|
NHMW 1910/0015/0034 |
|
Paraostenia striata |
|
|
|
NHMW 1910/0015/0035 |
|
Atropicaris? sp. |
|
|
|
NHMW 1910/0015/0036 |
|
Paraostenia striata |
|
|
|
NHMW 1910/0015/0037 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0038 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0039 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0040 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0041 |
|
Austriocaris carinata |
|
holotype |
|
NHMW 1910/0015/0042 |
|
Paraostenia striata |
|
holotype |
|
NHMW 1910/0015/0043 |
|
Paraostenia striata |
|
|
|
NHMW 1910/0015/0044 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0045 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0046 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0047 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0048 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0049 |
|
Austriocaris carinata |
|
|
|
NHMW 1910/0015/0050 |
|
Austriocaris carinata |
|
|
|
NHMW 2021/0123/0002 |
1501 |
Austriocaris carinata |
-50 cm to 0 cm |
|
|
NHMW 2021/0123/0059 |
|
Atropicaris? sp. |
|
|
|
NHMW 2021/0123/0203 |
2077 |
Austriocaris carinata |
-50 cm to 0 cm |
|
|
NHMW 2021/0123/0764 |
|
Austriocaris carinata |
|
|
|
NHMW 2021/0123/0765 |
39 |
Austriocaris carinata |
-50 cm to 0 cm |
|
|
NHMW 2021/0123/0766 |
52 |
Austriocaris carinata |
-50 cm to 0 cm |
|
|
NHMW 2021/0123/0767 |
194 |
Atropicaris? sp. |
-50 cm to 0 cm |
|
|
NHMW 2021/0123/0769 |
273 |
Atropicaris? sp. |
-50 cm to 0 cm |
|
|
NHMW 2021/0123/0770 |
|
Paraostenia striata |
|
|
|
NHMW 2021/0123/0772 |
1193 |
Paraostenia striata |
-50 cm to 0 cm |
|
|
NHMW 2021/0123/0776 |
4320 |
Atropicaris? sp. |
300 cm to 320 cm |
|
|
NHMW 2021/0123/0777 |
4323 |
Atropicaris? sp. |
300 cm to 320 cm |
|
|
NHMW 2021/0123/0779 |
4331 |
Atropicaris? sp. |
300 cm to 320 cm |
|
|
NHMW 2021/0123/0780 |
4334 |
Atropicaris? sp. |
300 cm to 320 cm |
|
|
NHMW 2021/0123/0782 |
4335 |
Atropicaris? sp. |
300 cm to 320 cm |
|
|
NHMW 2021/0123/0783 |
4412 |
Atropicaris? sp. |
300 cm to 320 cm |
|
|
NHMW 2021/0123/0784 |
4456 |
Atropicaris? sp. |
300 cm to 320 cm |
|
|
NHMW 2021/0123/0787 |
4464 |
Atropicaris? sp. |
300 cm to 320 cm |
|
|
NHMW 2021/0123/0788 |
4466 |
Atropicaris? sp. |
300 cm to 320 cm |
|
|
NHMW 2021/0123/0790 |
4540 |
Atropicaris? sp. |
300 cm to 320 cm |
|
|
NHMW 2021/0123/0791 |
4445 |
Paraostenia striata |
300 cm to 320 cm |
|
|
NHMW 2021/0123/0793 |
4739 |
Atropicaris? sp. |
300 cm to 320 cm |
|
|
NHMW 2021/0123/0795 |
4864 |
Atropicaris? sp. |
320 cm to 340 cm |
|
|
NHMW 2021/0123/0796 |
4894 |
Paraostenia striata |
320 cm to 340 cm |
|
|
NHMW 2021/0123/0797 |
|
Paraostenia striata |
|
|
|
NHMW 2021/0124/0078 |
|
Atropicaris? sp. |
|
|
|
NHMW 2021/0124/0101 |
|
Atropicaris? sp. |
|
|
|
NHMW 2021/0124/0102 |
|
Atropicaris? sp. |
|
|
|
NHMW 2021/0124/0103 |
|
Atropicaris? sp. |
|
|
|
NHMW 2021/0124/0104 |
|
Atropicaris? sp. |
|
|
|
NHMW 2021/0124/0105 |
|
Atropicaris? sp. |
|
|
|
NHMW 2021/0124/0106 |
|
Atropicaris? sp. |
|
|
|
NHMW 2021/0124/0108 |
|
Austriocaris carinata |
|
|
|
NHMW 2021/0124/0109 |
|
Austriocaris carinata |
|
|
|
NHMW 2021/0124/0110 |
|
Austriocaris carinata |
|
|
|
NHMW 2024/0201/0001 |
|
Paraostenia cf. ambatolokobensis |
300 cm to 340 cm |
|
Appendix 2
Measurements (in mm) made on the specimens. Abbreviations: Aad, anterodorsal angle; Aav, anteroventral angle; Apd, posterodorsal angle; Apv, posteroventral angle; Ha, anterior height; Hmax, maximal height; Hp, posterior height; Lr, rostrum length; Ls, total shield length; Lw, shield length without the rostrum; NA, non-applicable.
|
Specimen |
Species |
Ls |
Lw |
Lr |
Ha |
Hmax |
Hp |
Aad |
Aav |
Apv |
Apd |
|
NHMW 1910/0015/0002 |
Austriocaris carinata |
83.023 |
72.972 |
10.126 |
NA |
NA |
6.036 |
NA |
81.919 |
145.215 |
83.763 |
|
NHMW 1910/0015/0041 |
Austriocaris carinata |
100.259 |
89.099 |
11.179 |
32.062 |
NA |
9.388 |
60.532 |
83.555 |
146.304 |
82.497 |
|
NHMW 1910/0015/0050 |
Austriocaris carinata |
76.059 |
67.965 |
8.17 |
26.271 |
NA |
NA |
61.767 |
81.369 |
NA |
80.562 |
|
NHMW 1910/0015/0034 |
Paraostenia striata |
28.086 |
NA |
NA |
12.539 |
16.796 |
6.215 |
68.502 |
77.001 |
90.051 |
87.975 |
|
NHMW 1910/0015/0042_part |
Paraostenia striata |
26.061 |
NA |
NA |
13.678 |
15.127 |
6.206 |
68.189 |
73.36 |
92.131 |
64.568 |
|
NHMW 1910/0015/0042_counter-part |
Paraostenia striata |
26.394 |
NA |
NA |
13.078 |
15.104 |
6.291 |
63.339 |
69.272 |
89.86 |
64.301 |
|
NHMW 2024/0201/0001 |
Paraostenia
cf. |
31.354 |
NA |
NA |
14.979 |
19.362 |
4.560 |
68.516 |
71.310 |
107.324 |
97.696 |