
Late Ordovician (Sandbian–Hirnantian) marine gastropods from the Argentine Precordillera: their biogeographical significance in a middle to high latitudinal setting
MARIEL FERRARI, VERÓNICA BERTERO, and MARCELO G. CARRERA
Ferrari, M., Bertero, V., and Carrera, M.G. 2024. Late Ordovician (Sandbian–Hirnantian) marine gastropods from the Argentine Precordillera: their biogeographical significance in a middle to high latitudinal setting. Acta Palaeontologica Polonica 69 (4): 747–767.
Gastropods from the Upper Ordovician of the Argentine Precordillera received less attention than other coeval marine invertebrates in this region. The present contribution supplies accurate taxonomic information recovering 10 gastropod genera which are represented by 10 species from the La Pola and Don Braulio formations (Sandbian and Hirnantian units, respectively) at the San Juan Province, Argentina: two species, namely Tetranota argentina sp. nov. and Clathrospira gondwanica sp. nov., are new to science. The gastropod association shows a remarkable diversity, and in contrast to other invertebrate groups reported in the Argentine Precordillera (e.g., trilobites, ostracodes, brachiopods, sponges, and bivalves) which had Gondwanan affinities during the Late Ordovician, the gastropod assemblage from the La Pola and Don Braulio formations had major palaeobiogeographical similarities with their Northern Hemisphere (Laurentia, Avalonia, and Baltica) counterparts. The occurrence of the genus Clathrospira suggests that it could have been the first precursor of the order Pleurotomariida in South America during the Sandbian–Hirnantian, and can help to shed light on the origin of this clade in the southernmost Gondwana continent as early as Palaeozoic times.
Key words: Gastropoda, systematics, palaeobiogeography, Argentine Precordillera, Late Ordovician.
Mariel Ferrari [mferrari@cenpat-conicet.gob.ar; ORCID: https://orcid.org/0000-0001-6422-9806 ], Instituto Patagónico de Geología y Paleontología, IPGP (CCT CONICET-CENPAT), Boulevard Alte. Brown 2915, (9120), Puerto Madryn, Chubut, Argentina.
Verónica Bertero [vbertero@apn.gob.ar; ORCID: https: //orcid.org/0009-0007-8377-7411 ], Dirección Nacional de Conservación, Administración Parques Nacionales (Dir. Regional Centro)-Córdoba, Argentina.
Marcelo G. Carrera [mcarrera@unc.edu.ar; ORCID: https://orcid.org/0000-0003-0013-9195 ], CICTERRA (CONICET-Universidad Nacional de Córdoba), Facultad de Ciencias Exactas, Físicas y Naturales, Av. Vélez Sarsfield 1699, X5016GCA, Córdoba, Argentina
Received 4 September 2024, accepted 16 October 2024, published online 30 December 2024.
Copyright © 2024 M. Ferrari et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Ordovician of the Argentine Precordillera is well known for its exquisite outcrops yielding abundance of taxa. Most of them have been already mentioned or described in detail (Benedetto 1986, 2003, 2004; Benedetto et al. 2009; Carrera 1994; Carrera and Ernst 2010; Halpern and Carrera 2014; Ernst and Carrera 2012, 2022; Fernández Martínez et al. 2004; Sánchez et al. 2002; Vaccari 1995; Waisfeld et al. 1999; 2003, and references therein). Conversely gastropods were not treated and only a few references and descriptions are available in literature.
The first descriptions of Ordovician Precordilleran gastropods were given by Kayser (1876) who reported representatives of the genera Ophileta Vanuxem, 1842, Maclurites Le Sueur, 1818, and Murchisonia d’Archaic, 1841, and subsequently, some taxa were mentioned by Kobayashi (1935, 1937). Rohr et al. (2001) provided a list of gastropods from the San Juan Formation, including 14 genera that were not well preserved and, thus, not formally described. Later, Bertero (2009) described the genus Malayaspira Kobayashi, 1958, from the middle interval of the San Juan Formation and recently, Bertero et al. (2023) provided valuable information about the opercula and associated shell in life position of two Teiichispira species from the same formation. Dzik (2019) figured protoconchs of some gastropod genera (e.g., Raphistoma, Holopea, Mimospira, Tropidodiscus, Sinuites) from the Darriwilian, Middle Ordovician of the San Juan Formation. These gastropod reports from the Lower/Middle Ordovician carbonates of the Argentine Precordillera contrast with the absence of descriptions up to now of taxa from the Upper Ordovician siliciclastic units of the Precordillera.
In the present contribution we described 10 gastropod genera which are represented by 10 species from the La Pola and Don Braulio formations of a Sandbian–Hirnantian age; these are Sphenosphaera sp., Bucania sp., Sinuites aff. S. reticulatus Perner, 1903, Tropidodiscus sp., Trochonemella? sp., Scalites peracutum (Ulrich & Scofield 1897), Holopea sp., Cyclonema aff. C. bilix (Conrad, 1842). Two of them, namely Tetranota argentina sp. nov. and Clathrospira gondwanica sp. nov., are new to science. We interpret these new species as endemic to Argentina, and Clathrospira gondwanica sp. nov. as the first precursor of the order Pleurotomariida in South America during the Late Ordovician.
The gastropod association here described shows a remarkable diversity almost reaching the diversity known from other conspicuous fossil groups, such as brachiopods, which were described from the same formations by Benedetto (1986, 1990, 2015).
We analyze in this work the temporal and palaeogeographic distribution of the Argentinean gastropods and their biogeographic affinities during the Late Ordovician.
Institutional abbreviations.—CEGH-UNC, Centro de Investigaciones en Ciencias de la Tierra (CICTERRA), Universidad Nacional de Córdoba, Argentina.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:43849C9B-D0EB-406F-AA9B-D791604D8167.
Geological setting
The Argentine Precordillera consists of a thick Palaeozoic sedimentary succession mainly of Cambro-Ordovician rocks, showing a transition from a nearshore carbonate bank through mixed carbonate-siliciclastic slope deposits to basinal clastics (Fig. 1).
Lower–Middle Ordovician rocks consist of Tremadocian to Darriwilian platform carbonates (the La Silla and San Juan formations, Keller et al. 1994; Cañas 1999). In general, carbonate sedimentation ceased during the early Darriwilian due to regional drowning, and the deposition of graptolitic black shales and mudstones (the Los Azules and Gualcamayo formations). After this change in the sedimentary regime, and following a late Darriwilian hiatus, mixed calcareous-siliciclastic sedimentation prevailed during the Sandbian (the Las Plantas and Las Aguaditas formations).
Ordovician deposition culminates with Hirnantian glacial diamictites followed by fossiliferous transgressive mudstones (Sánchez et al. 1991; Buggish and Astini 1993).
In the Villicum range, easternmost Precordillera basin (Fig. 1), the La Cantera Formation served as the base of the Sandbian units. This unit is partially a lateral equivalent to the mixed siliciclastic carbonate Las Aguaditas Formation with its diverse bryozoan fauna studied by Ernst and Carrera (2012).
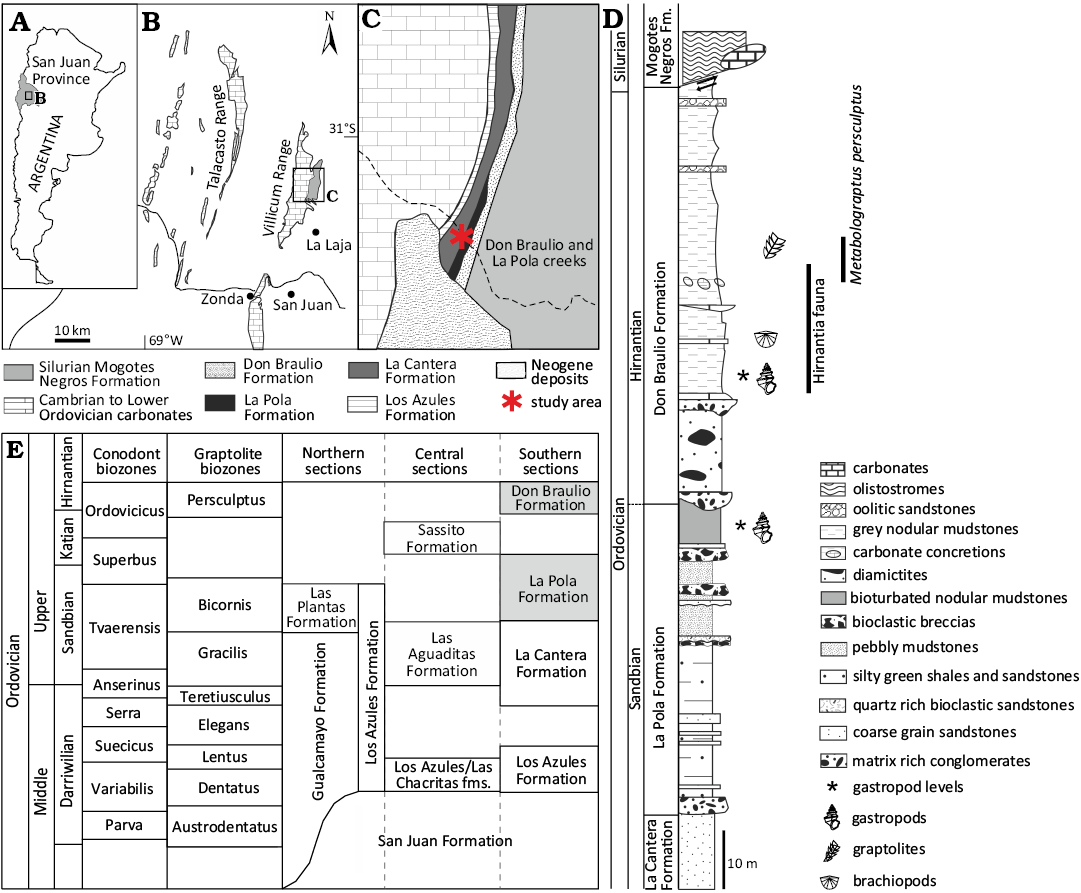
Fig. 1. Geographical, stratigraphical and biostratigraphical setting of the Argentine Precordillera during the Late Ordovician. A. Map showing location of the study area in the San Juan Province, Argentina. B, C. Geological maps of the study area including the outcrops of Upper Ordovician marine deposits of the Don Braulio and La Pola formations. D. Stratigraphic column of the La Pola and Don Braulio formations at Sierra Villicum, and detail of sampled interval showing the gastropod fauna associations described in the text. E. Biostratigraphical scheme of La Pola and Don Braulio formations based on conodonts and graptolites.
The La Cantera Formation is a sand-dominated unit that yields massive polymictic conglomerates at its base and fines upwards. Biostratigraphical constrains at its top suggests a Sandbian age based on the presence of graptolite Nemagraptus gracilis recorded by Peralta (1986) within the silty shales near its top (Fig. 1B).
The La Pola Formation and coeval or partially coeval units (the Las Plantas, Las Aguaditas and Sassito formations) are similar in that their faunas are all dominated by bryozoans (Carrera 2003; Ernst and Carrera 2008, 2012, 2022) accompanied by abundant brachiopods. The Hirnantian Don Braulio Formation has lower bryozoan diversity (Halpern and Carrera 2014).
The La Pola Formation represents a succession of thick-bedded coarse-grained debris flows (ranging from mud to clast supported) with interbedded pebbly mudstones, amalgamated lenticular quartz-bioclastic-rich sandstones, as well as a few turbidites and silty shales. Facies associations suggest a depositional environment related to a proximal deep-marine trough. Sedimentary provenance points to a quartz-rich, high-energy shelf as the source and a coeval or intermixed carbonate interval (Astini 2001). Among the known fauna from the La Pola Formation, mainly composed of diverse brachiopods, bryozoans, and crinoid remains, gastropods have so far been poorly known.
Stratigraphically older graptolites and conodonts have been recovered from various clasts, with the youngest assigned to the Nemagraptus gracilis/Dicranograptus clingani biozones. Brussa (2000) identified the graptolites Dicranograptus ramosus ramosus, Dicranograptus nicholsoni, and other taxa which indicate a Sandbian age (Gisbornian 1, 2). More recently, a conodont association from the La Pola Formation allowed definition of the Amorphognathus tvaerensis Zone (upper Sandbian–lower Katian), and within this zone the association can be assigned to the Baltoniodus variabilis Subzone (Heredia and Milana 2010) (Fig. 1B).
The Don Braulio Formation unconformably overlies the La Cantera and La Pola formations (Darriwilian–Katian) and underlies the Silurian Mogotes Negros Formation (Ludlovian–Pridolian) (Fig. 1C). The Hirnantian age of this unit has been established on the basis of brachiopods belonging to the cosmopolitan Hirnantia fauna (Benedetto 1986) and the record of the Metabolograptus persculptus Zone (Brussa et al. 2003). The Don Braulio Formation diamictite has been interpreted as glacial-marine in origin based on its strong erosional base, striated pavements, and the presence of dropstones (Fig. 1C; Peralta and Carter 1990; Buggisch and Astini 1993). It is spatially restricted in both lateral and vertical extension.
Material and methods
The material here described occurred in two Upper Ordovician units (the La Pola and Don Braulio formations corresponding to a Sandbian–Hirnantian age). In the La Pola Formation (upper Sandbian–lower Katian), gastropods are found in a bioturbated nodular mudstone at the top of the formation near the upper boundary with the unconformably base of the Don Braulio Formation (Hirnantian) (Fig. 1C). Some of the Sandbian gastropods occur as allochthonous remains in the conglomerate at the base of the Don Braulio Formation (some forms are found in both units).
Gastropods of the Don Braulio Formation occur in the grey nodular mudstone just above the Hirnantian diamictites in the Metabolograptus persculptus Zone (Brussa et al. 2003) (Fig. 1C).
The biogeographical analysis based on the Precordilleran gastropod genera here described was performed using PAST (Hammer et al. 2001). Cluster analysis was performed applying the Dice similarity coefficients, which emphasize matches in dataset (Hammer and Harper 2006).
Systematic palaeontology
Class Gastropoda Cuvier, 1797
Subclass Amphigastropoda Simroth, 1906
Order Bellerophontida Ulrich & Scofield 1897
Superfamily Bellorophontoidea McCoy, 1852
Family Bellerophontidae McCoy, 1852
Subfamily Bucanopsinae Wahlman, 1992
Genus Sphenosphaera Knight, 1945
Type species: Bellerophon clausus Ulrich in Ulrich & Scofield, 1897, by original designation; from the Bigby-Cannon Formation of the Upper Ordovician, North America.
Remarks.—Wahlman (1992: 165) proposed a diagnosis for the genus as “shell moderately rapidly expanding, aperture flaring posteriorly and laterally, whorls floor with distinct median ridge, ridge no present on parietal shelf, ornament collabral”. The author also stated that representatives of Sphenosphaera have small to moderate-sized shells, rounded but generally not globose and narrow or closed umbilicus. The aperture is broader than long, and the shell margins flared laterally and posteriorly, producing flat to slightly excavated posterolateral shelves. The slit is shallow to moderately deep, housed in a broad sinus. The selenizone is distinct, either developing a rounded to flat-topped ridge or a concave channel, in many cases with distinct lunulae and a thin revolving thread bordering on each side. The shell surface is marked by fine and closely spaced growth lines (Wahlman 1992: 165).
The present research provides the first occurrence of Sphenosphaera in the lower Palaeozoic (Upper Ordovician) of Argentina and South America.
Stratigraphic and geographic range.—Ordovician to Silurian; Europe, North and South America (Wahlman 1992; this paper).
Sphenosphaera sp.
Fig. 2A, B.
2020 Sphenosphaera? sp.; Bertero 2020: 37, pl. 1: 1–4.
Material.—CEGH-UNC 24795–24800, six recrystallized teleoconchs; from the Sandbian, Upper Ordovician, La Pola Formation, Sierra de Villicum, San Juan Province, Argentina.
Description.—Anomphalous, rounded but not globose, small-sized shell with a height and length of 8–15 mm. Flanks rapidly expanding; median-dorsal shell slightly elevated with a distinct selenizone. The aperture is 1.98 mm wider than long, and the margins flared laterally and posteriorly forming flat to slightly excavated posterolateral shelves. The spire is sub-rhomboidal in apical view and growth lines weakly visibly on the shell surface. As wide and deep and a V-shaped sinus (Fig. 2A3).

Fig. 2. Sandbian–Hirnantian (Late Ordovician) gastropods from La Pola and Don Braulio formations at the Sierra Villicum locality, Argentine Precordillera. A, B. Bucanopsin bellerophontoid Sphenosphaera sp. A. CEGH-UNC 24795-2, in lateral (A1), dorsal (A2, A3), and apical (A4, A5) views. B. CEGH-UNC 24796-6, in ventral view. C. Bucaniin bellerophontoid Bucania sp., CEGH-UNC 24733-1, in lateral (C1) and dorsal (C2) views. D. Plectonotin bellerophontoid Tetranota argentina sp. nov., CEGH-UNC 24713, holotype, in dorsal (D1, D5, D6) and dorsal-lateral (D2, D3) views, cross section (D4) and ornament detail (D7). E−G. Sinuitin bellerophontoid Sinuites aff. S. reticulatus Perner, 1903. E. CEGH-UNC 24694-1, in apertural and basal view. F. CEGH-UNC 24696, in basal and apertural (F1) and lateral (F2−F4) views. G. CEGH-UNC 24697-3, in dorsal-lateral view (G1, G2) and ornament detail (G3, G4). H. CEGH-UNC 24694, in lateral view.
Remarks.—The type species of Sphenosphaera was originally ascribed to the genus Bellerophon by Ulrich in Ulrich and Scofield (1897). Wahlman (1992) suggested that Sphenosphaera differs from Bellerophon in being less globose, in having more flared apertural margins which form prominent posterolateral shelves and in having a distinct rounded median ridge on the whorl floor. The Argentinean species is comparable to Sphenosphaera clausus (Ulrich in Ulrich and Scofield, 1897) (in Wahlman 1992: 167, pl. 30: 1–23, pl. 31: 1–13, pl. 32: 1–15), from the Katian, Upper Ordovician of North America. The Laurentian species, however, has a selenizone on an elevated flat-topped median carina bordered by fine revolving threads and distinct lunulae, and closely spaced and regular growth lines which are broadly curved over the dorsal slopes in maturity. Sphenosphaera troosti (Férussac and d’Orbigny, 1840) (in Wahlman 1992: 168, pl. 29: 1–22), from the Katian, Upper Ordovician of North America, differs from Sphenosphaera sp. in having markedly broader and inflated whorls, a more prominent selenizone located on a rounded dorsomedian ridge, and closely spaced and fine growth lines. Sphenosphaera burginensis (Ulrich in Ulrich and Scofield, 1897) (in Wahlman 1992: 170, pl. 28: 10–23), also from the Katian, Upper Ordovician of North America, has narrower whorls that are expanding rather slowly, a selenizone elevated on a thin dorsomedian carina, and closely spaced and thin growth lines. Sphenosphaera recurvus (Ulrich in Ulrich & Scofield, 1897) (in Wahlman 1992: 171, pl. 33: 1–6), from the Katian, Upper Ordovician of North America, is much narrower in dorsal view than Sphenosphaera sp., has the aperture more subtriangularly elongated and not expanding laterally, has strongly curved growth lines, a deep sinus, and the selenizone is markedly elevated and truncated. Finally, Sphenosphaera capax (Ulrich in Ulrich & Scofield, 1897) (in Wahlman 1992: 172, pl. 34: 1–13), also from the Katian, Upper Ordovician of North America, is very similar to the Argentinean species in shell shape, although the Laurentian form is more globose than Sphenosphaera sp. and much larger with a length and height of 38–26 mm, respectively.
Family Bucaniidae Ulrich & Scofield, 1897
Subfamily Bucaniinae Ulrich & Scofield, 1897
[= Grandostomatinae Horný, 1962]
Genus Bucania Hall, 1847
Type species: Bellerophon sulcatinus Emmons, 1842, by subsequent designation of Waagen (1880); from the Chazyan Stage of the Middle Ordovician, North America.
Remarks.—According to Wahlman (1992: 127), representatives of Bucania are characterized by having a small to large-sized shell, with rounded or depressed whorls gradually expanding, and with a medium or large umbilicus. The aperture has a thin lip with a broad V-shaped sinus and a central slit; the slit is shallow or quite deep. The selenizone is generally narrow, being slightly elevated, flat or channel-like. The shell surface is ornamented by transverse growth lines or lamellae and spiral threads; spiral threads run normal and are generally interrupted by growth lines. Growth lines may become lamellose in mature growth stages and accompanied by a laterally thickening. Considering these characters, the specimens analyzed here are assigned to Bucania.
This is the first occurrence of Bucania in the lower Palaeozoic (Upper Ordovician) of Argentina and South America.
Stratigraphic and geographic range.—Lower–Middle Ordovician to middle Silurian; North America, Europe, Asia and South America.
Bucania sp.
Fig. 2C.
2020 Bucania sp.; Bertero 2020: 39, pl. 1: 5, 6.
Material.—CEGH-UNC 24733–24736, four recrystallized teleoconchs; from the Sandbian, Upper Ordovician, La Pola Formation, Sierra de Villicum, San Juan Province, Argentina.
Description.—Shell relatively small, narrow with three or four rounded whorls. The height is 20 mm and the width 18 mm. The aperture has a thin lip and a broad V-shaped sinus. Slit not clearly visible. The selenizone is central, narrow and slightly elevated; the umbilicus is widely open forming an umbilical angle of 85° (Fig. 2C1). The shell is dorsally convex, ornamented by regularly spaced, crowded and transversal growth lines that are intercepted by more or less separated spiral threads.
Remarks.—Taking into consideration the characters such as a widely open umbilicus, the presence of a V-shaped sinus, a narrow selenizone and a shell surface ornamented by spiral threads, the material herein described certainly represents a member of Bucania. Bucania emmonsi Ulrich in Ulrich and Scofield, 1897 (in Wahlman 1992: 128, pl. 12: 1–4), from the Sandbian–Katian, Upper Ordovician of North America, is comparable to Bucania sp., although the Laurentian form is much smaller, has stronger and more deeply incised spiral threads and a slightly concave selenizone. Bucania lindsleyi (Safford, 1869) (in Wahlman 1992: 131, pl. 18: 1–9), from the Katian, Upper Ordovician of North America, differs from the Argentinean species in having more rapidly expanding whorls, a wide and broadly concave selenizone which is marked by strong lunulae. Bucania frankfortensis Ulrich in Ulrich & Scofield, 1897 (in Wahlman 1992: 133, pl. 13: 22–44, pl. 16: 1–4), also from the Katian, Upper Ordovician of North America, has a distinctly triangular outline, narrower whorls, regularly spaced and wrinkled growth lines and very weak spiral threads. Bucania rugatina Ulrich in Ulrich and Scofield, 1897 (in Wahlman 1992: 136, pl. 16: 9–15; pl. 17: 1–12), from the Katian, Upper Ordovician of North America, differs from the Argentinean species in having a narrower and deeper umbilicus, a wider selenizone bordered by fine ridges, closely spaced growth lines and wrinkled and sublamellose spiral threads. Bucania gracillima Koken, 1896 (in Ebbestad 1998: 27–28, figs. 2, 3), from the Sandbian–Katian, Upper Ordovician of Sweden, is much larger than Bucania sp. (length and height 88–72 mm, respectively), has a more developed ornament pattern consisting on crenulated growth lines, and a clearly visible dorsal selenizone with lunulae. Bucania erratica Frisk & Ebbestad, 2007 (89, fig. 4I–T; Ebbestad et al. 2013: fig. 20: 8O–W), from the Sandbian, Upper Ordovician of Sweden, differs from the Argentinean form in having four depressed lenticular whorls, and ornamentation consisting on dense, crenulated and non-lamellose growth lines. Bucania czekanowskii (Schmidt, 1858) (in Isakar and Ebbestad 2000: 57, figs. 6a–h, 7a–i), from the Sandbian, Upper Ordovician of Estonia, differs from Bucania sp. in having an ornament pattern consisting of flaring growth lines on regular intervals that create a geometrical pattern resembling triangles in lateral view. Bucania cornu Koken, 1897 (in Isakar and Ebbestad 2000: 66, fig. 12a–g), from the Katian, Upper Ordovician of Estonia, has a slit reaching about 1/2 whorl back, but close to form a trema when the widely expanded aperture is developed in shells of maturity. Finally, Bucania radiata (d’Eichwald, 1856) (in Isakar and Ebbestad 2000: 61, fig. 11a–k), from the Katian, Upper Ordovician of Estonia, differs from the species here described in having a marked expansion of the whorl during ontogeny and an increased angulation of the dorsal surface at the median carina giving a distinctly acute median part of the whorl.
Subfamily Plectonotinae Boucot & Yochelson, 1966
Genus Tetranota Ulrich & Scofield, 1897
Type species: Bucania bidorsata Hall, 1847, by original designation; from the Watertown Formation (Trenton Group) of the Turinian, Upper Ordovician, North America.
Remarks.—Wahlman (1992: 144) proposed the diagnosis of the genus Tetratona to include “shells with selenizone lying on broad, elevated dorsomedian band and usually bordered by a pair of revolving threads, with one or two pairs of dorsolateral revolving angular ridges; the apertural lip chiefly expanded laterally”. According to Wahlman (1992), species of Tetranota also have shells variable in size with the umbilicus more or less large and deep. The whorls are broad and depressed and the aperture wider than long throughout growth stages. The selenizone is located on a broad, elevated, gently convex to nearly flat topped dorsomedian band, which generally has revolving threads along its edges; lunulae faint in most cases. Outward from the dorsal band, the shell is briefly concave and has a dorsolateral pair of angular revolving ridges. The growth lines are fine and closely spaced.
The present research records the first occurrence of Tetranota in the lower Palaeozoic (Upper Ordovician) of Argentina and South America.
Stratigraphic and geographic range.—Ordovician; North America, England, and South America (Argentina).
Tetranota argentina sp. nov.
Fig. 2D.
2020 Tetranota bidorsata (Hall, 1847); Bertero 2020: 42, pl. 1: 12, 13, pl. 2: 1–3.
ZooBank LSID: urn:lsid:zoobank.org:act: 06C088B3-73F8-4451-9C49 -E1A04F4AEA24.
Etymology: Referred to the first occurrence of the genus in the lower Palaeozoic (Upper Ordovician) of Argentina.
Type material: Holotype CEGH-UNC 24713 (Fig. 2D), partially preserved teleoconch without protoconch and aperture. Paratypes: CEGH-UNC 24714, 24715, 24716, from the type horizon and locality.
Type locality: Sierra de Villicum locality, San Juan Province, Argentina.
Type horizon: La Pola Formation, Sandbian, Upper Ordovician.
Material.—Type material and 38 specimens composed of internal moulds with patches of external shell with fragmentary ornamentation (CEGH-UNC 24700–24712, 24717–24732, and 24774–24782) from the type locality and horizon.
Diagnosis.—Shell involute and subreiniform with a moderately wide umbilicus (umbilical angle of 70º). A dorsal and broad median band carries a concave selenizone. Two acute revolving threads border the selenizone, and two central-lateral grooves are present outward the dorsomedian band. The strong reticulate ornament pattern consists of distinct and fine growth lines which are intercepted by spiral ribs.
Description.—Involute, depressed, whorl profile subreiniform, small to medium-sized shell with a width of 15 mm and a length of 13 mm. Umbilicus deep and moderately wide with an angle of 70º, (Fig. 2D1, D2). In cross section the whorls are lenticular, with well rounded periphery and without thickening of the shell (Fig. 2D4). Dorsum with prominently elevated, broad median band of 2.3 mm (6 times the width of the shell) that carries a slightly concave selenizone (Fig. 2D1, D5, D6). The selenizone is bordered by two angular and acute revolving threads. Outward from the dorsomedian band, a pair of central-lateral concave grooves are visible. Dorsal slopes are also visible near the umbilical shoulder. Ornament consists of raised and distinct, regularly and densly spaced fine growth lines which are intercepted by spiral lirae forming a reticulate pattern. Spiral lirae run into an anteroposterior direction.
Remarks.—According to the characterization of Wahlman (1992) the new species here described represents a true member of Tetranota. Shells identified here as Tetranota argentina sp. nov. were originally ascribed by Bertero (2020) to the type species Tetranota bidorsata (Hall, 1847) (in Wahlman 1992: 145, pl. 21: 1–14, pl. 22: 1–10), from the Katian, Upper Ordovician of North America, considering that both are very similar in shell shape ornamentation. However, the type species is slightly larger, has a more elevated dorsal median band and a more concave selenizone bordered by more sharper revolving threads, has stronger growth lines and weaker spiral lirae, and thus, shows a weaker reticulate ornament pattern than T. argentina sp. nov. Wahlman (1992) also described other species of Tetranota from the Katian, Upper Ordovician of North America. One of these Laurentian species, Tetranota obsoleta Ulrich in Ulrich & Scofield, 1897 (in Wahlman 1992: 146, pl. 20: 13–15, pl. 22: 16–21), differs, however, from Tetranota argentina sp. nov. in having a smaller shell, with a narrower and deeper umbilicus, a dorsomedian band slightly elevated and concave, and lunulae and growth lines poorly preserved. Tetranota sexcarinata Ulrich & Scofield, 1897 (in Wahlman 1992: 147, pl. 22: 11–15; in Bertero 2020: 44, pl. 2: 4–8), from the Sandbian–Katian, Upper Ordovician of Argentina and North America, differs from T. argentina sp. nov. in having two pairs of acute dorsolateral ridges on both sides of the elevated dorsomedian selenizone, more widely spaced growth lines, and lacks spiral lirae and a reticulate ornament pattern. Tetranota wisconsinensis (Whitfield, 1878) (in Wahlman 1992: 148, pl. 38: 13–16), from Sandbian–Katian, Upper Ordovician of North America, has a larger and more tightly coiled shell than the new species, with the dorsolateral ridges gradually disappearing during ontogeny. Tetranota scanica Ebbestad et al., 2013 (fig. 8L–N), from the Katian, Upper Ordovician of Sweden, differs from T. argentina sp. nov. in having finer and sharper growth lines and denser and more irregularly spaced lunulae.
The genus Tetranota (with the species Tetranota sexcarinata Ulrich & Scofield, 1897), has been previously reported in the Middle Ordovician of Argentina by Bertero (2020: 44, see above); thus, T. argentina sp. nov. is the first report of the genus in the Sandbian, Upper Ordovician, marine deposit of the Precordillera Argentina, which extends the palaeobigeographical distribution of the genus to South America during the early Palaeozoic.
Stratigraphic and geographic range.—Sandbian, Upper Ordovician, La Pola Formation, Sierra de Villicum, San Juan Province, Argentina.
Family Sinuitidae Dall, 1913
Subfamily Sinuitinae Dall, 1913
Genus Sinuites Koken, 1896
Type species: Bellerophon bilobatus Sowerby, 1839, by subsequent designation of Bassler (1915); from the Soudleyan–Longvillian, Upper Ordovician of England.
Remarks.—Wahlman (1992: 110) characterized representatives of Sinuites as having tightly coiled and convolute shells, with a closed or very small umbilicus. The dorsum is generally rounded, somewhat flattened with a dorsomedian ridge. The aperture is large and narrow, with a deep, rounded, median sinus and lateral margins projecting downward as rounded or subangular lobes. Parietal deposits are thin. Shell generally ornamented with closely spaced and fine growth lines which may be intersected by faint revolving lines. Following the characterization of Wahlman (1992), the specimens here analyzed are assigned to Sinuites.
Ebbestad and Yochelson (2000) highlighted that there have been much controversy related to this genus and regarded the discussion on preserved muscle scars of torted versus untorted molluscs. Horný (1996) also addressed a number of the problems related to morphology, systematic delimitation and ecology of Sinuites, proposing a semi-infaunal, predatory mode of life for this genus.
Here we supply the first occurrence of Sinuites in the Upper Ordovician, lower Palaeozoic of Argentina and South America.
Stratigraphic and geographic range.—Ordovician; North America, South America, Europe, Baltic region, and North Africa.
Sinuites aff. S. reticulatus Perner, 1903
Fig. 2E–H.
2020 Sinuites aff. S. reticulatus Perner, 1903; Bertero 2020: 45, pl. 1: 7, 8; pl. 2: 9–12; pl. 3: 1.
Material.—CEGH-UNC 24694–24699, six incomplete teleoconchs preserved with fragmentary ornament pattern; from the Sandbian, Upper Ordovician, La Pola Formation, Sierra de Villicum, San Juan Province, Argentina
Description.—Involute, rounded and elongated to subglobose, small to medium-sized shell with 16 mm large and 9 mm width, with closed umbilicus (or cryptomphalous). The teleoconch is slightly compressed laterally. The aperture is oval, and the lateral and anterior-dorsal margins of the shell are covered by parietal inductura visible in specimen CEGH-UNC 24696 (Fig. 2F). The sinus is moderately shallow. The sculpture consists of a reticulate pattern of spiral almost concentric irregular wavy revolving lirae near the labro-umbilical depression (Fig. 2G).
Remarks.—The material here described strongly resembles Sinuites reticulatus Perner, 1903 (Horný 1996: 94), from the Darriwilian, Middle Ordovician of Czech Republic in shell shape and ornamentation; however, in S. reticulatus two visible layers of inductural deposits are distinguished, the lower, carrying lirae and corrugation which is located directly over the uppermost shell layer. Another similar species to Sinuites aff. S. reticulatus is Sinuites sowerbyi Perner, 1903 (in Horný 1997: 48, pl. 5: 1–2), from the Darriwilian, Middle Ordovician of Morocco, but it has a more feather-like ornament pattern covering the ribs within the dorsal sinus, and 5–6 regularly spaced collabral lines per mm. Sinuites destombesi Horný, 1997 (48, pl. 5: 3–10, pl. 6: 1–11, pl. 7: 1–3), from the Katian–Hirnantian, Upper Ordovician of Morocco, is slightly smaller than the Argentinean species, has a well-developed subinductura around the umbilicus and a dense network of collabral and spiral elements. Sinuites cancellatus Hall, 1847 (Wahlman 1992: 111, pl. 11: 6–12), from the Katian, Upper Ordovician of North America, is very similar to S. aff. S. reticulatus; although, S. cancellatus has a moderately shallow and V-shaped sinus, an inductura ornamented by fine and wavy spiral lines, and fine growth lines and even finer spiral elements on the shell surface. Another species comparable to S. aff. S. reticulatus is Sinuites globularis (Miller & Faber, 1894) (Wahlman 1992: 114, pl. 10: 1–13), also from the Katian, Upper Ordovician of North America; but the Laurentian form has broader and more inflated whorls, a dorsum with distinct and round dorsomedian ridges bounded by a shallow and deep sinus, and fine and closely spaced growth lines. Sinuites granistriatus (Ulrich in Ulrich and Scofield, 1897) (Wahlman 1992: 113, pl. 11: 1–5, pl. 21: 1; pl. 45: 1–9), also from the Katian, Upper Ordovician of North America, resembles Sinuites aff. S. reticulatus, even though the Laurentian species has a clearly visible dorsomedian ridge in juvenile growth stages which tends to disappear toward mature whorls. Sinuites cf. niger Koken, 1897 (in Ebbestad et al. 2013: 317, fig. 9D–J), from the Sandbian–Katian, Upper Ordovician of Sweden, has the subinductura in the near part of the last whorl consisting of irregular wrinkled wavy lines that may bifurcate. Sinuites usitatus Isakar, 1991 (49, pl. 4–5; in Ebbestad and Yochelson 2000: 192, fig. 5A–I), from the Katian–Hirnantian, Upper Ordovician of the Baltic region, is slightly larger than the Argentinean form, has a wider and open umbilicus, strong wrinkles on dorsal part of the shell, and a post-apertural constriction on the last whorl. Finally, Sinuites sp. (in Ebbestad 2016: 89, fig. 3A–D), from the Floian, Lower Ordovician of Morocco, is less globose and more compressed laterally than Sinuites aff. S. reticulatus, has a weakly open umbilicus, and lacks the wavy revolving lirae near the periumbilical area.
Family Tropidodiscidae Knight, 1956
[= Temnodiscinae Horný, 1963]
Genus Tropidodiscus Meek & Worthen, 1866
Type species: Bellerophon curvilineatus Conrad, 1842, by original designation of Meek and Worthen (1866); from the Eifelian, Middle Devonian of North America.
Remarks.—Members of this genus have a laterally compressed and lenticular shell, consisting on several whorls, whorls expanding gradually, with a large open umbilicus and deep sutures. Tropidodiscus also is characterized by the presence of a V-shaped sinus that leads into deep slit, a very narrow selenizone on a narrow keel, with or without lunulae, and ornament consisting on prosocyrt growth lines curving backward (Wahlman 1992: 118; Ebbestad and Yochelson 2000: 194). The species of Tropidodiscus have also small to large sized-shells which may run from few to more than 30 mm, the slender shell with the elongated aperture has been interpreted as well suited for high mobility (Linsley 1978; Wahlman 1992; Ebbestad and Yochelson 2000) but small members of the genus could have fed on algal fronds (Ebbestad and Yochelson 2000).
Ebbestad and Yochelson (2000) addressed that Tropidodiscus has a wide palaeobiogeographical distribution and occurs in Early Ordovician to Devonian in Laurentia, Baltica, Siberia, Kazakhstan and Gondwana including more than 50 species, but Ebbestad et al. (2008) suggested that the genus is most widely distributed along the western Tethyan sea (Avalonia).
Here we supply the first occurrence of Tropidodiscus in the Upper Ordovician of Argentina and South America.
Stratigraphic and geographic range.—Lower Ordovician (Canadian) to Upper Devonian; North and South America, Europe, North Africa and Asia.
Tropidodiscus sp.
Fig. 3A, B.
2020 Tropidodiscus sp. Bertero 2020: 47, pl. 1: 9, 10.
Material.—CEGH-UNC 24677–24693; seventeen incomplete specimens preserved as inner and external moulds; from the Sandbian, Upper Ordovician, La Pola Formation, Sierra de Villicum and Quebrada de Don Braulio, San Juan Province, Argentina.
Description.—Lenticular, planispiral and strongly compressed shell, with a wide and open umbilicus. The typical acute keel is not clearly visible. The teleoconch consists of about 3 whorls. The profile is triangular in transversal section (Fig. 3A). The length of the shell 20−70 mm and the umbilical angle 80−90º. The median selenizone is slightly acute, and growth lines are not clearly visible in the available specimens.

Fig. 3. Sandbian–Hirnantian (Late Ordovician) gastropods from La Pola and Don Braulio formations at the Sierra Villicum and Quebrada Don Braulio localities, Argentine Precordillera. A, B. Tropidodiscid bellerophontoid Tropidodiscus sp. A. CEGH-UNC 24679-5 in transversal view. B. CEGH-UNC 24678 in lateral view. C. Trochonematoid Trochonemella? sp., CEGH-UNC 24752, in lateral and apertural (C1), apical and lateral (C2), and lateral (C3, C4) views. D−G. Raphistomatid Scalites peracutum (Ulrich & Scofield 1897). D. CEGH-UNC 24772, in lateral view. E. CEGH-UNC 24783, in apical view. F. CEGH-UNC 24773, in apical view. G. CEGH-UNC 24786-9, in basal and umbilical views. H, I. Holopeid vetigastropod Holopea sp. H. CEGH-UNC 25451, associated specimens in lateral view. I. CEGH-UNC 25448, in lateral view. J−N. Eotomariid Clathrospira gondwanica sp. nov. J. CEGH-UNC 24737, paratype, in lateral view. K. CEGH-UNC 24738-11, holotype, in lateral view (K1, K2). L. CEGH-UNC 24740, in apical (L1) and lateral (L2) views. M. CEGH-UNC 24738-11, holotype, in lateral (M1) and basal (M2) views. N. CEGH-UNC 24739, in apical (N1) and basal and apertural (N2) views. O–Q. Platyceratid Cyclonema cf. bilix (Conrad, 1842). O. CEGH-UNC 24765, in apical view (O1), ornament detail (O2). P. CEGH-UNC 24766-1, in lateral views (P1, P2). Q. CEGH-UNC 24767-2, in lateral view.
Remarks.—The specimens analyzed show the typical character of Tropidodiscus. Tropidodiscus subacutus (Ulrich & Scofield, 1897) (in Wahlman 1992: 118, pl. 41: 9–13), from the Katian, Upper Ordovician of North America, differs from the Argentinean form in being slightly smaller, has the dorsum acutely carinate with a narrow truncation at the crest of the selenizone, a wider umbilicus, a very narrow and V-shaped sinus, and fine growth lines. Another similar species to Tropidodiscus sp. is Tropidodiscus cristatus (Safford, 1869) (in Wahlman 1992: 119, pl. 41: 6–8), from the Katian, Upper Ordovician of North America; although the former has a strongly acute whorl floor keel. Tropidodiscus magnus (Miller, 1878) (in Wahlman 1992: 119, pl. 42: 1, 2), from the Katian, Upper Ordovician of North America, has a slightly more compressed shell, a narrower umbilicus and less expanded last teleoconch whorl. Tropidodicus (Peruniscus) sp. (in Rohr 1980: 150, pl. 1: 15–16), from the Middle/Upper Ordovician of North America, has a slightly more acute and subangular keel. Tropidodiscus sp. (in Ebbestad et al. 2008: 610, fig. 3.18–20), from the Upper Ordovician of Iran, is much smaller than the Argentinean species. Tropidodiscus laticonchus Ebbestad & Yochelson, 2000 (194, fig. 8A–G), from the Upper Ordovician of Norway, has a more inflated and lenticular shell, with more whorls of uniform expansion rate, a distinct raised median selenizone, a wider umbilicus and ornament with clearly visible prosocyrt growth lines. Topidodiscus maximus Ebbestad & Yochelson, 2000 (196, fig. 9A–H), also from the Katian, Upper Ordovician of Norway, is much larger that the Argentinean species with a length of 380 mm, the teleoconch consists of 5 to 6 whorls, and it has a distinct raised and narrow selenizone and weak prosocyrt growth lines. Tropidodiscus pusillus (Barrande in Perner, 1903) (in Horný 1997: 52, pl. 9: 10, 11, pl. 10: 1–5), from the Darriwilian, Middle Ordovician , of Morocco, has a more laterally compressed shell, a more acute and conspicuous median selenizone, and the collabral lines are clearly visible in the shell surface.
Order uncertain (Palaeozoic Basal Taxa that are Certainly Gastropoda sensu Bouchet et al. 2017)
Superfamily Trochonematoidea Zittel, 1895
Family Trochonematidae Zittel, 1895
Genus Trochonemella Okulitch, 1935
Type species: Trochonemella notablis (Ulrich & Scofield, 1897), by original designation; from the Black River Group of lower Silurian, Kentucky, North America.
Remarks.—Members of Trochonemella are characterized by the presence of a narrowly phaneromphalous shell; sinus relatively shallow culminating in a wide notch that gives rise to a selenizone (Knight et al. 1960).
Stratigraphic and geographic range.—Ordovician of North America and South America.
Trochonemella? sp.
Fig. 3C.
2020 Trochonemella sp.; Bertero 2020: 81, pl. 7: 5, 7, 10.
Material.—CEGH-UNC 24752; one recrystallized teleoconch; from the Sandbian, Upper Ordovician, La Pola Formation, Sierra de Villicum and Quebrada de Don Braulio, San Juan Province, Argentina.
Description.—Trochiform, gradate, step-like, low-spired and medium-sized shell, with a height of 70 mm and a width of 90 mm; pleural angle of 110º. Sutures impressed. The teleoconch consists of about five whorls, last whorl more expanded than the spire; juvenile whorls are convex (Fig. 3C). On mature whorls the ramp is slightly concave and becomes convex to the lower portion of whorls (Fig. 3C2). A deep spiral furrow is located above mid-whorl and delimiting the sutural ramp and the lower portion of the whorl. The spiral furrow is bordered by two conspicuous spiral ribs. Collabral elements are not visible. The base is flat to slightly angular. Umbilical characters are not preserved. The aperture is subangular.
Remarks.—The present material does not show some of the diagnostic features of Trochonemella, such as the shallow sinus and the wide notch that give rise to the selenizone; thus, it is tentatively assigned to the genus. Trochonemella reusingi Rohr, 1988 (in Blodgett and Rohr 2012: 3), from the Upper Ordovician of North America was interpreted as one of the most common and conspicuous fossils from this region. It differs from Trochonemella? sp. in being larger and having strongly spinose peripheral spiral cords. Trochonemella churkini Rohr & Blodgett, 1985 (670, figs. 2.7–2.10), from the Katian, Upper Ordovician of west-central Alaska, differs from the Argentinean species in having a more turbinate and gradate outline shell, a selenizone at mid-whorl with clearly visible lunulae, an upper-outer angulation of whorls with scoop-like protrusions and collabral threads above sutures. Another species comparable to Trochonemella? sp. is Trochonemella? mikulici Rohr, 1980 (176, pl. 8: 21–24), from the Katian, Late Ordovician of North America, although T.? milkulici has strong orthocline growth lines, a strong raised selenizone above mid-whorl, lunulae at selenizone clearly visible in mature whorls, and orthocline growth lines on the shell surface. Trochonemella antelopensis Rohr, 1996 (59, figs. 3.14–3.17), from the Middle and Upper Ordovician of North America, differs from the Argentinean species in having a raised selenizone located above mid-whorl, a slightly concave whorl surface below selenizone, labral edge with strongly prosocline to opisthocyrt growth lines, lower–upper angulation with short frill, and a circumumbilical spiral cord. Trochonemella sp. (Ebbestad and Stott 2008: 235), from the Upper Ordovician of Canada, is smaller than the Argentinean species with a maximum height of 27 mm.
Superfamily uncertain (unassigned to superfamily in Bouchet et al. 2017)
Family Raphistomatidae Koken, 1896
[= Ceratopeidae Yochelson & Bridge, 1957]
Genus Scalites Emmons, 1842
Type species: Scalites angulatus Emmons 1842, by monotypy; from the Chazy Group of the Middle Ordovician, North America.
Remarks.—Based on phylogenetic analyses, Wagner (2002) suggested including within Scalites all species previously classified as Raphistoma Hall, 1847. According to that author, the type species S. angulatus shares several synapomorphies with Raphistoma, and thus, he pointed out that it would be more consistent with the phylogeny to use Scalites for the species of Raphistoma. Members assigned to Scalites are diagnosed by flattened ramps that (owing to the overall orientation of the aperture) are nearly perpendicular to the coiling axis, strong adapically hooked peripheral bands, and strong channeled basal carina (Wagner 2002).
Stratigraphic and geographic range.—Lower Ordovician–Silurian of North America, Europe and South America.
Scalites peracutum (Ulrich & Scofield, 1897)
Fig. 3D–G.
1897 Raphistoma peracuta Ulrich & Scofield, 1897.
1928 Raphistoma peracuta Ulrich & Scofield, 1897; Twenhofel 1928: 236.
1987 Rhaphistoma peracuta Ulrich & Scofield, 1897; Sloan and Webers 1987: 185.
2002 Scalites peracutum (Ulrich & Scofield, 1897); Wagner 2002: 74.
2020 Scalites peracutum (Ulrich & Scofield, 1897); Bertero 2020: 59, pl. 4: 3–7.
Material.—CEGH-UNC 24783–24794, CEGH-UNC 24771–24773; twelve recrystallized teleoconchs and three moulds from Sierra Villicum and Quebrada Don Braulio localities, Argentina, Sandbian, Upper Ordovician.
Description.—Lenticular, phaneromphalous, medium to small sized and low-spired shell with the upper surface nearly flat to convex. The height is 70 mm and the width is 20–35 mm. The teleoconch has a subtriangular shape in transversal view and consists of four whorls (Fig. 3D). Sutures are deeply incised in a spiral furrow. The peripheral selenizone is slightly wide and flat and bears prosocline lunulae. The ornament on the upper whorl consists of fine and strongly prosocline growth lines (Fig. 3E, fragmentary ornament pattern on the periphery of last whorl). The base is flat and angular ornamented by prosocline growth lines. The umbilicus is wide, deep and funnel shaped, ornamented by acute axial ribs with a prosocline and orthocline orientation at regular intervals followed by a slightly flared increment (Fig. 3G). The axial ribs are intercepted by two acute spiral ribs. A sharp basal carina with rounded nodes borders the umbilical area (Fig. 3G).
Remarks.—Representatives of Scalites shares several characters of Raphistoma Ulrich & Scofield, 1897. Raphistoma is diagnosed by having a flat to convex shell with a triangular shape in transversal view, a flat spire, and a sharp periphery bearing the selenizone. Rohr et al. (1992) also pointed out that Scalites and Raphistoma are very similar raphistomatids, with Scalites known from three species and Raphistoma with over twenty species assigned.
The species herein described differs from the type species Scalites angulatus Emmons, 1842 (Rohr et al. 2004), from the Middle Ordovician of North America, in having a more flattened spire, a subtriangular shell in transversal view, in lacking a thickening of the columella and strong growth lines on base, and in having a widely open umbilicus; S. angulatus is anomphalous. The specimens here described as Scalites peracutum strongly resembles the specimens of S. peracutum (Ulrich & Scofield, 1897) from the Ordovician of North America, in general shell morphology, although the North American form is much larger. Scalites? alaskensis (Rohr et al. 1992: 505: 3.1–7, 5), from the Middle Ordovician of Alaska, North America, has a narrowly open umbilicus, a more gradate shell, and ornament with fine spiral threads. Scalites amanoi Kobayashi, 1958, S. keizanensis (Kobayashi, 1934), S. katoi Kobayashi, 1934, and S. irregular Kobayashi, 1934, from the Lower/Middle Ordovician of South Korea, are based on poorly preserved specimens.
Stratigraphic and geographic range.—Sandbian, Upper Ordovician, La Pola Formation, Sierra de Villicum and Quebrada de Don Braulio, San Juan Province, Argentina.
Subclass Vetigastropoda Salvini-Pläwen, 1980
Superfamily uncertain (Palaeozoic taxa of uncertain position sensu Bouchet et al. 2017)
Family Holopeidae Cossmann, 1908
Genus Holopea Hall, 1847
Type species: Holopea symmetrica Hall, 1847, by subsequent designation of Bassler (1915); from the Chatfieldian, Upper Ordovician of North America.
Remarks.—Wagner (2002) recognized Holopea as a derived genus with reduced shell structures and suggested that it was restricted to the Ordovician. Tremadocian–Floian, Lower Ordovician species have been described from Canada (Billings 1865) and North Korea (Kobayashi 1931) but the majority of species range through the Sandbian and Katian, Upper Ordovician; the youngest species occur in the Hirnantian, uppermost Ordovician of Estonia (Koken and Perner 1925) and Canada (Twenhofel 1928) (see Ebbestad et al. 2013).
Stratigraphic and geographic range.—Upper Ordovician; North America, Europe, North Korea, Baltic region, and South America.
Holopea sp.
Fig. 3H, I.
2020 Holopea sp.; Bertero 2020: p. 54, pl. 3: 14, pl. 6: 3.
Material.—CEGH-UNC 25445–25452, eight recrystallized teleoconch; from the Hirnantian, Upper Ordovician, Don Braulio Formation (level 2B), Quebrada de la Vinchuca, Sierra de Villicum, San Juan Province, Argentina.
Description.—Globose, turbiniform, small sized-shell with a height of 30−57 mm and a width of 27−58 mm. Pleural angle of 70°−80°. The teleoconch consists of 2 or 3 1/2 rapidly expanding and strongly convex whorls. Sutures incised. The ornament consists of fine orthocline growth lines. Basal, umbilical and apertural characters not visible in the available material.
Remarks.—Holopea is a quite problematic genus considering that it lacks well defined and precise diagnostic characters for its taxonomic assignment (Yochelson 1963; Wagner 2002). Rohr (1980) ascribed the pleural angle as a character to distinguish different species within the genus. Following Rohr’s (1980) criteria, Holopea sp. is similar to Holopea glindmeyeri Rohr, 1980 (181, pl. 9: 4–8), from the Middle Ordovician of North America, which shows a pleural angle of 75–90°; however, the North America species is much smaller, and each whorls embrace previous whorl at about midline. Holopea elizabethi Rohr, 1980 (180: pl. 9: 28–37), from the Middle/Upper Ordovician of North America, also shows a similar pleural angle of 72°, but differs from the Argentinean species in having a higher spire and closely spaced and distinct prosocline growth lines. Holopea brucei Rohr, 1980 (181, pl. 9: 25–27), from the Middle Ordovician of North America, has a wider pleural angle of 105°, a narrow and deep umbilicus, closely spaced prosocline growth lines. Holopea mobergi Troedsson, 1918 (in Ebbestad et al. 2013: 318, fig. 10), from the Katian–Hirnantian, Upper Ordovician of Sweden, differs from the Argentinean species in having a much smaller shell and a slightly more pointed spire. Radvanospira antiquata (Perner, 1903) (in Horný 1997, as Holopea? antiquata: 60, pl. 15, 16; Ebbestad et al. 2019: 456, figs. 13a–g, 14a–h) from the Sandbian–Hirnantian, Upper Ordovician of Spain, Morocco, Bohemia and Iberia, has a more globose or naticiform shell, with more distinctly and dense growth lines and an auriform aperture. Holopea sp. (in Frýda et al. 2001: 125, fig. 3A, B), from the Katian–Hirnantian, Upper Ordovician of Spain, has fine spiral and collabral elements on the shell surface forming a reticulate ornament pattern, and strongly prosocline growth lines on last whorl. Considering that Holopea? antiquata was removed to the genus Radvanospira (see Ebbestad et al. 2019), the species here described is the first certain report of Holopea in the Hirnantian, Late Ordovician environments of Gondwana.
Order Pleurotomariida Cox & Knight, 1960
Superfamily Eotomarioidea Wenz, 1938
Family Eotomariidae Wenz, 1938
Subfamily Eotomariini Wenz, 1938
Genus Clathrospira Ulrich & Scofield, 1897
Type species: Pleurotomaria subconica Hall, 1847, by original designation; from the Middle Ordovician of North America.
Remarks.—Members of Clathrospira have a turbiniform and narrowly phaneromphalous shell, with a conical spire and shallow sutures, growth lines that are strongly prosocline above selenizone and strongly opisthocline below it, but rounding to gently prosocline on base, outlining a moderately deep labral sinus that culminates in a short slit; selenizone bordered by fine threads; growth lines and lunulae periodically strengthened, very fine and numerous spiral threads (Knight et al. 1960).
According to Horný (1997), the genus is imperfectly known and defined, lacks modern revision and its interpretation is rather wide (Ulrich and Scofield 1897; Longstaff 1924; Koken and Perner 1925; Vostokova 1955; Wagner 2002). Horný (1997) pointed out that the single modern detailed paper concerning one European species, Clathrospira elliptica (Hisinger, 1831) from the Oslo region, is that of Yochelson (1963). However, C. elliptica has later been revised by Wager (2002) and Frisk and Ebbestad (2007).
The very fine revolving lira, a character which is present in the type species, has not been largely accepted as diagnostic of Clathrospira (see Horný, 1997: 57). The genus is also diagnosed by having a very strong bilineate peripheral band that is located very low on the whorl owing to a very short left ramp relative to the right ramp (Wagner 2002).
Karapunar et al. (2022) argued that Pleurotomariida have the most extended fossil record among the living gastropods with one of their oldest occurrences in the Ordovician with the genus Clathrospira (Horný 1997; Wagner 2002; this paper). According to Karapunar et al. (2022), Pleurotomariida was the most diversified gastropod group in the middle and late Palaeozoic and dominant in shallow water communities at that time.
The report of the genus Clathrospira, and the new species Clathrospira gondwanica sp. nov. (see below), in the Upper Ordovician of the Argentine Precordillera, provides new and updated taxonomic information enabling to interpret the origin of the order Pleurotomariida in South America at about 458.4–445.2 Mya.
Stratigraphic and geographic range.—Middle Ordovician–Silurian; North America, South America and Europe.
Clathrospira gondwanica sp. nov.
Fig. 3J–N.
2020 Clathrospira subconica (Hall, 1847); Bertero 2020: 72, pl. 6: 7, 11, 12, 14; pl. 7: 1–5.
ZooBank LSID: urn:lsid:zoobank.org:act: 04F60BE6-63A4-43D4-B65 A-02932BFFBC7A.
Etymology: Referred to the first occurrence of Clathrospira in Gondwana (South America).
Type material: Holotype CEGH-UNC 24738 (11), complete and well preserved teleoconch without protoconch and juvenile whorls. Paratype CEGH-UNC 24737, complete and well preserved teleoconch with juvenile whorls; from the type locality and horizon.
Type locality: Sierra de Villicum locality, San Juan Province, Argentina.
Type horizon: La Pola and Don Braulio formations, Sandbian–Hirnantian, Upper Ordovician, lower Palaeozoic.
Material.— Type material and 25 recrystallized teleoconchs (CEGH-UNC 24739–24751, 24753–24764) from type locality and horizon.
Diagnosis.—Shell conical, trochiform, anomphalous and coeloconical to cyrtoconical. The pleural angle is variable and ranges 60–100°. The peripheral selenizone is situated above mid-whorl. The selenizone is cord-like shaped on juvenile whorls and concave on mature whorls, and lunulae are opisthocyrt. The prosocline growth lines on the flank of whorls are visible above selenizone. The growth lines are prosocyrt to orthocline on base. The aperture is subquadrangular.
Description.—Trochiform, conical, anomphalous, medium-sized shell with a height of 40−95 mm and a width of 40−85 mm. The pleural angle is variable and ranges 60−100°. The teleoconch consists of six whorls; spire whorls are coeloconoidal and last whorl becomes slightly cyrtoconoidal (Fig. 3K2). The selenizone is peripheral and located slightly above mid-whorl; opisthocyrt lunulae are clearly visible (Fig. 3J). The selenizone has a cord-like shape on spire whorls and turns into slightly deep, concave and U-shaped on last whorl (Fig. 3J) and it is bordered by two acute spiral lirae. The ornament consists of strongly prosocline and fine growth lines on the flank of whorls above the selenizone; the growth lines are intercepted by weak and irregularly spaced spiral cords. On last whorl, below the selenizone and towards the base the growth lines become sligthly prosocyrt to orthocline. The base is convex and the aperture subquadrangular.
Remarks.—The species here described is very similar in shell shape and ornamentation to the type species Clathrospira subconica (Hall, 1847), but the Argentinean form shows neither the narrow umbilicus nor the fine spiral threads. Clathrospira glindmeyeri Rohr, 1996 (59, pl. 2: 11–17, 23; assigned by Ebbestad and Peel 2001 to the genus Semizona), from the Whiterochian, Middle Ordovician of North America, differs from Clathrospira gondwanica sp. nov in having a more convex spire whorls with an cyrtoconoid shape, in having a narrow umbilicus, more acute prosocline growth lines, and a peripheral cord-shaped selenizone bordering the upper edge of last whorl. Deaechospira amouguerana (Horný, 1997) (57, pl. 12: 7–13; pl. 13: 1–3, originally assigned to Clathrospira amouguerana; Ebbestad et al. 2019: 261, figs. 17a–h, 18a–k, 19a–e), from the Sandbian–Katian, Upper Ordovician of Morocco, is very similar to Clathrospira gondwanica sp. nov., but differs in having a more arched whorls below the selenizone.
Stratigraphic and geographic range.—Sandbian–Hirnantian, Upper Ordovician, La Pola and Don Braulio formations, Sierra de Villicum and Quebrada de Don Braulio, San Juan Province, Argentina.
Subclass Neritimorpha Golikov & Starobogatov, 1975
Superfamily Platyceratoidea Hall, 1859
Family Platyceratidae Hall, 1859
Genus Cyclonema Hall, 1852
Type species: Pleurotomaria bilix Conrad, 1842, by original designation; from the lower Silurian of North America.
Remarks.—Knight et al. (1960) characterized members of Cyclonema as turbiniform to trochiform shells, with sharp spiral ribs and fine collabral threads. The diagnosis proposed by Thompson (1970) also characterized Cyclomena species as anomphalous shells, with an polygonal to auriform aperture, columellar lip lunate and excavates, ornament consisting on three orders of spiral lines cancellated by collabral lirae commonly thicken into growth wrinkles.
Stratigraphic and geographic range.—Middle Ordovician–Silurian; North America, South America, Europe.
Cyclonema cf. bilix (Conrad, 1842)
Fig. 3O–Q.
2020 Cyclonema bilix (Conrad, 1842); Bertero 2020: 69, pl. 8: 8–11.
Material.—CEGH-UNC 24765–24770, six recrystallized teleoconchs; from the Sandbian, Upper Ordovician, La Pola Formation, Quebrada La Pola (Level LP1), Sierra de Villicum and Quebrada de Don Braulio, San Juan Province, Argentina.
Description.—Turbiniform to trochiform, medium-sized shell with a height of 15 mm and a width of 10–18 mm; pleural angle of 55−85º. Teleoconch consists of four whorls in the best preserved specimen (Fig. 3O1). The ramp of whorls is flattened to slightly concave and excavated; the periphery of whorls is strongly convex. Sutures are impressed in a spiral channel. The ornament consists of three orders of regularly spaced spiral lirae. The spiral lirae are intercepted by fine and acute prosocline growth lines forming a reticulate pattern with small and rounded nodes at the intersection points (Fig. 3O2). The base is flattened to slightly convex and the aperture is not completely visible; the inner lip probably concave.
Remarks.—The species herein described from the Upper Ordovician of Argentina strongly resembles Cyclonema bilix (Conrad, 1842) from the Katian, Upper Ordovician of North America. According to Thompson (1970) C. bilix has a trochiform to conical shell with straight to concave whorl profile and rounded periphery, and is characterized by the presence of three orders of spiral ribs which form a reticulate pattern with strongly prosocline collabral lirae; these features are clearly visible in the material from the Argentine Precordillera. Cyclonema cf. bilix, however, has less conical and more convex whorl profile, the typical polygonal to rounded aperture, the straight to concave inner lip and the outer lip reflected obliquely on adults is not completely visible in our material and, thus, it is tentatively assigned to the species. Cyclonema humerosum Ulrich & Scofield, 1897 (in Thompson 1970: 244, pl. 32: 10–13), from the Katian, Upper Ordovician of North America, differs from C. cf. bilix in having a more deeply channeled suture, a broader and horizontal ramp, a more angular periphery and stronger and more acute spiral lirae. Cyclonema inflatum Ulrich & Scofield, 1897 (in Thompson 1970: 245, pl. 32: 14–18), also from the Katian, Upper Ordovician of North America, has a more slender and higher spire than the Argentinean form, a more globose last whorl, and the ornament consists in two orders of acute spiral cords more separated on the upper part of the whorl. Cyclonema (C.) hiiumaa Teichert, 1928 (in Isakar 1995: 185, figs. 1–6), from the Silurian of Estonia, has a broadly D-shaped aperture with the inner lip thickened and reflected.
Palaeobiogeographical inferences
During the Furongian and Middle Ordovician, the Precordillera or Cuyania terrane would have drifted away from Laurentia and located within the southern Iapetus Ocean until its final docking with western Gondwana during the Middle–Late Ordovician (Astini et al. 1995; Benedetto 2004; Keller 2012; Ramos 2004). According to this scenario, the Argentine Precordillera was situated as an isolated terrane in the middle of the Iapetus Ocean during the Sandbian (Fig. 4), still with a biogeographic North American signature in their faunas, but receiving some faunal influence from Gondwana and Baltica (see Benedetto 2003; Benedetto et al. 1999, 2009).
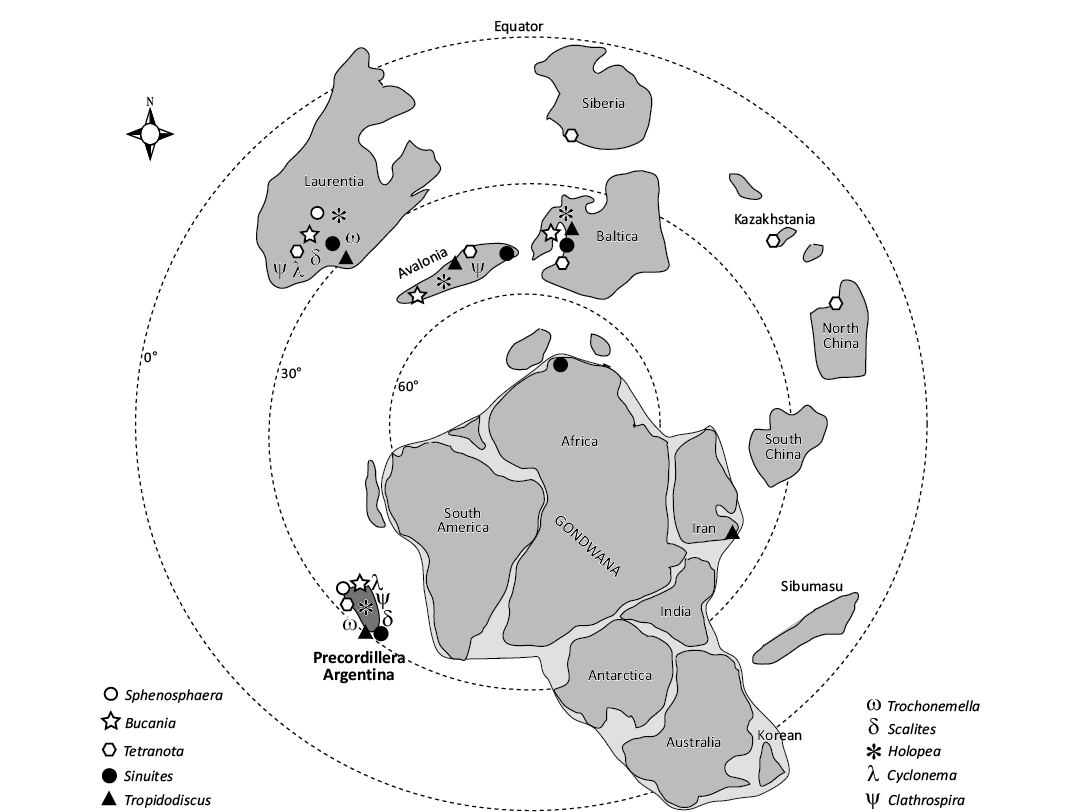
Fig. 4. Palaeobiogeographical map showing the distribution patterns of the gastropod genera here reported during the Late Ordovician worldwide (modified from Bertero et al. 2023). Note that the genus Clathrospira could be the first precursor of the order Pleurotomariida in South America during the Sandbian–Hirnantian intervals.
The palaeogeographic history of the Precordillera terrane shows that it shifted from equatorial to higher latitudes (Astini 1999; Keller et al. 1998). Warm-water carbonates were deposited during the Cambrian and Early Ordovician, whereas Middle Ordovician units set at mid-latitude (30–45°) locations, including the Katian Sassito limestones (Ernst and Carrera 2008). The Hirnantian glacigenic rocks of the Don Braulio Formation represent the last step in the shifting trajectory of the Precordillera terrane (Astini 1999; Benedetto et al. 2009, 2011). Biogeographic analyses of trilobites, ostracodes, brachiopods, sponges, and bivalves that occur in the Argentine Precordillera were carried out to assess the changes in palaeogeographic position of this region (Benedetto et al. 1999, 2009; Sánchez et al. 2002 and references therein). These studies showed that faunal affinities of the Argentine Precordillera were essentially Laurentian during Cambrian and Early Ordovician times. Later, the degree of endemicity increased during the isolation of the Argentine Precordillera and finally, affinities with Gondwana arose after the Darriwilian. From the Late Ordovician to the early Silurian the faunal similarities exhibited a clear Gondwana pattern. These palaeobiogeographical interpretations were based on many invertebrate taxa from the Argentine Precordillera except gastropods. Here, we attempt to assess the gastropod palaeogeographic affinities during the Sandbian–Hirnantian, Late Ordovician worldwide.
The gastropod assemblage from the Argentine Precordillera showed a cosmopolitan distribution during the Late Ordovician. These genera were typical components of Laurentia (North America), Avalonia (Europe), Baltica (Norway, Sweden and Estonia), Siberia (Russia), North China, Kazakhstan, Iran and Gondwana (Morocco and South America) (Fig. 4). The first occurrences of these genera in South America extend their palaeobiogeographical distribution into the southernmost Gondwana continent at a Sandbian–Hirnantian age.
The genera Sphenosphaera, Trochonemella, Scalites, and Cyclonema are known from the Ordovician (Sphenosphaera and Trochonemella) reaching up to the Silurian (Scalites and Cyclonema), and show a restricted palaeobiogeographical distribution during the Katian, Late Ordovician, when they occurred in Laurentia only (see Table 1) (Rollins 1967; Thompson 1970; Rohr 1980, 1996; Rohr and Blodgett 1985; Rohr et al. 1992, 2004; Wahlman 1992; Isakar 1995; Blodgett and Rohr 2012). Here we supply the first report of these genera in the Late Ordovician (Sandbian) of the Argentine Precordillera, which was an isolated terrain near southern Gondwana by that time (Fig. 4, Table 1).
The genera Sinuites, Bucania, Tetranota, Tropidodiscus, Clathrospira, and Holopea are reported from the Ordovician to the Silurian/Devonian (Clathrospira and Holopea reached the Silurian–Devonian) and, in contrast with the genera mentioned above, they show a wide palaeobiogeographical distribution during the Late Ordovician occurring in Laurentia, Avalonia, Baltica, Siberia, North China, Kazakhstan, Iran and Morocco, except Clathrospira that was considered to be restricted to Laurentia thus far (see Fig. 4 and Table 1) (Twenhofel 1928; Kobayashi 1931, 1934; Vostokova 1955; Knight et al. 1960; Thompson 1970; Pickerill and Brenchley 1979; Rohr 1980, 1996; Wahlman 1992; Horný 1996, 1997; Ebbestad, 1998, 2016; Ebbestad and Yochelson 2000; Isakar and Ebbestad 2000; Frýda et al. 2001; Frisk and Ebbestad 2007; Ebbestad et al. 2013; Isakar 1991, 1995; Wagner, 2002). Here we supply the first report of these genera in the Sandbian–Hirnantian of the Argentine Precordillera and the first report of Clathrospira in the Gondwana continent (Fig. 4; Table 1). Particularly, Bucania has been mentioned in the lower Tremadocian, Lower Ordovician of NW Argentina with the species Bucania sp. (in Aceñolaza 1968) but neither description nor illustrations of the species have been provided. The present research provides the first certain occurrence of Bucania in the Sandbian of the Argentine Precordillera. Sinuites and Holopea exhibited a Gondwana distribution being reported in Morocco (Sinuites) and in the Argentine Precordillera (Holopea) during the Hirnantian (see Table 1); on the other hand, Tetranota was one of the most widespread gastropod genera during the Late Ordovician. Ebbestad and Yochelson (2000) and Ebbestad et al. (2008) pointed out that Tropidodiscus was most widely distributed along the western Tethyan sea (Mediterranean Province) by that time. Wagner (2002) restricted the genus Clathrospira to Laurentia, and this occurrence was also supported by Ebbestad et al. (2013, 2019). The new report of Clathrospira in the Sandbian–Hirnanatian of the Argentine Precordillera is the first certain occurrence of the genus in Gondwana (Table 1).
Table 1. Table showing the palaeogeographical and stratigraphical distribution of the gastropod genera here described during the Late Ordovician. Note that the genera from the Argentine Precordillera sharing taxonomic affinities exclusively with Laurentia are highlighted.
|
Late Ordovician |
Late Ordovician Palaeobiogeographical Provinces |
|||||
|
Laurentia |
Avalonia |
Baltica |
Siberia |
Northern Gondwana (Morocco) |
Southern Gondwana |
|
|
Hirnantian |
|
Holopea |
Bucania Sinuites Holopea |
|
Sinuites |
Holopea Clathrospira |
|
Katian |
Sinuites Sphenosphaera Bucania Tetranota Holopea Trochonemella Clathrospira Cyclonema Tropidodiscus Scalites |
Holopea Sinuites Tropidodiscus Tetranota Clathrospira |
Bucania Sinuites Tropidodiscus Holopea Tetranota |
|
Sinuites |
|
|
Sandbian |
Bucania Tetranota |
Bucania |
Bucania Sinuites |
Tetranota |
|
Sinuites |
|
Sphenosphaera |
||||||
|
Bucania |
||||||
|
Tetranota |
||||||
|
Cyclonema |
||||||
|
Clathrospira |
||||||
|
Tropidodiscus |
||||||
|
Trochonemella |
||||||
|
Scalites |
||||||
During the Sandbian all the invertebrate groups in the Argentine Precordillera, including trilobites, ostracodes, brachiopods, bryozoans, bivalves, and gastropods show some degree of a cosmopolitan distribution, important endemism, and some genera with Baltic, Avalonian and even Laurentian affinities. In the Hirnantian, particularly brachiopods and trilobites were typical representatives of Hirnantia Fauna (Benedetto et al. 2011 and references therein) developed after the glacial deposits.
It is clear that the new gastropod association from the Argentine Precordillera shows a remarkable diversity almost reaching those of other fossils groups in the same interval. There are many genera represented in the Sandbian (the “Sandbian Group” see Fig. 4; Table 1), and these are: Sinuites, Tetranota, Sphenosphaera, Bucania, Cyclonema, Clathrospira, Tropidodiscus, Trochonemella, and Scalites. This group displays strong taxonomic similarities with the Laurentian, and also with Avalonian and Baltic faunas (mainly recorded in the Sandbian and Katian intervals in these regions). The palaeobiogeographical provinces showed in Table 1, including the Argentine Precordillera, share at least seven gastropod genera: Sinuites, Bucania, Tretranota, Tropidodiscus, Clathrospira, and Holopea (see Table 1). Four genera occur exclusively in Laurentia and Precordillera, and these are: Sphenosphera, Cyclonema, Trochonemella, and Scalites (Table 1, highlighted in grey).
The genera of the “Hirnantian Group” are scarce and distributed in Avalonia (Holopea), Baltica (Bucania, Sinuites and Holopea), Morocco (Sinuites) and in the Argentine Precordillera (Holopea and Clathrospira) (Table 1), and are not reported in Laurentia during the Hirnantian (Table 1). Holopea from the Hirnantian interval of the Argentine Precordillera occur in two main palaeogeographical regions including Avalonia and Baltica. This genus is found in the Katian of Laurentia, Avalonia and Baltica. In Morocco, Sinuites is recorded in the Katian and Hirnantian (Ashgill, according to Horný 1997), and Clathrospira and Holopea are absent in this palaeobiogeographical region. In contrast, in the Precordillera, Sinuites and Clathrospira first appeared in the Sandbian while Holopea occurred later in the Hirnantian. Other Sandbian forms of the Precordillera such as Bucania and Tetranota are typical of Avalonia, Baltica, and Siberia regions but also appear in Laurentia. This biogeographic pattern is consistent with the palaeogeographic trajectory noted for the Precordillera Terrain from the equator in the Cambrian and Early Ordovician moving to intermediate latitudes in the Sandbian, Late Ordovician, when still maintained the record of some Laurentian forms and with a biotic interchange between Avalonia and Baltica, sharing some genera such as Bucania, Sinuites, and Tetranota. Ebbestad et al. (2013) recognized Tetranota as a biostratigraphically important genus and summarized its palaeobiogeographical occurrence to Laurentia, Avalonia, Russia, Kazakhstan, China, Blatoscandia, and Gondwana.
Sinuites is present in the Katian–Hirnantian of Northern Gondwana (Morocco), being absent in Precordillera at that time (Table 1). On the other hand, Clathrospira and Holopea occur in the Hirnantian of the Precordillera Argentina associated with the Hirnantia fauna of high latitudes, while these genera are absent in Morocco in this time interval (Table 1). None of the genera here described are reported in Morocco during the Sandbian. Ebbestad et al. (2019) transferred the widely recognized Holopea? antiquata to the genus Radvanospira; here we follow this assignation and, thus, we do not consider Holopea as present among Late Ordovician Moroccoan taxa. According to Ebbestad et al. (2019) the genus Sinuites also showed affinities with the Bohemian faunas during the Late Ordovician.
Clathrospira and the cosmopolitan genus Tetranota are represented in the Argentine Precordillera by two new species such as Clathrospira gondwanica sp. nov. and Tetranota argentina sp. nov. Here we consider these taxa as endemic for the Sandbian–Hirnantian intervals in Argentina. Similar pattern was described from bryozoan faunas (Ernst and Carrera 2012, 2022; Halpern and Carrera 2014).
A quantitative biogeographical analysis is here applied to understand the interchange between the faunas of Precordillera Argentina with the other palaeobiogeographical regions mentioned in the text, and to testify the affinities of the Precordillera gastropod faunal assemblage during the Late Ordovician. Previous palaeobiogeographical analyses including Ordovician Precordilleran gastropods were carried out by Ebbestad et al. (2013). The results obtained by these authors show that the Precordilleran gastropods cluster with Tarim and the peri-Gondwana region (Perunica, Morocco, Ibero-Armorica, Alps and east and west Gondwana) in the Sandbian–Katian interval. The analysis of Ebbestad et al. (2013) also showed that the Precordilleran gastropods exhibited close affinities with their Laurentian counterparts until the Sandbian, and that the Precordillera shared with Avalonia the genus Tetranota and Tropidodiscus during the Sandbian–Katian. This pattern coincides with the data obtained in the present research (see Table 1). The results of the analysis show that the Argentine Precordillera clusters next to Laurentia in the Sandbian–Katian interval and this is consistent with the observations suggested for the “Sandbian Group” (see above) and by Ebbestad et al. (2013) (Fig. 5A). On the other hand, Precordillera clusters next to Avalonia for the Katian–Hirnantian intervals; this pattern may be explained in part by the presence of the genus Holopea in Avalonia during the Hirnantian (Fig. 5B; Table 1). Pecordillera is next to Morocco for the Sandian–Hirnantian intervals. The clustering between Precordillera and Morocco is consistent with the interpretations suggested above that from the Late Ordovician to the early Silurian the faunal similarities of Precordillera exhibited a clear Gondwana pattern (Fig. 5).
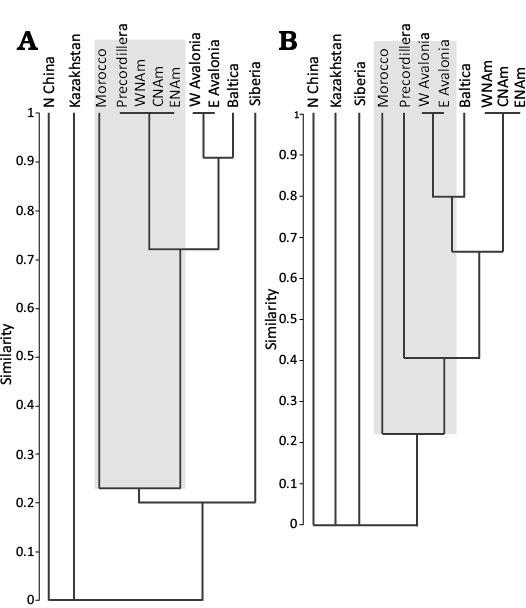
Fig. 5. Cluster analysis of Sandbian–Hirnantian gastropod genera described herein. A. Sandbian–Katian intervals; note that Precordillera clusters with Laurentia (central and western North America) and with Morocco. B. Katian–Hirnantian intervals; note that Precordillera clusters with Avalonia and also with Morocco. Abbreviations: CNAm, Central North America; ENAm, Eastern North America; WNAm, Western North America.
Finally, focusing our attention to Clathrospira, we interpret this genus as the first true pleurotomariid reported in the Late Ordovician of South America and in the southernmost Gondwana continent. Karapunar et al. (2024) suggested that the order Pleurotomariida (Subclass Vetigastropoda) has the longest fossil record among extant gastropod clades and represents one of the most diverse Palaeozoic and early Mesozoic gastropod groups (Erwin 1990; Karapunar and Nützel 2021; Karapunar et al. 2022). According to Karapunar et al. (2024), Pleurotomariida is the single extant gastropod clade with the fossil record extending back to the Ordovician and Cambrian periods and which reached its peak of diversity during the middle to late Palaeozoic (e.g., Knight et al. 1960; Hickman 1984; Wagner 2002; Karapunar and Nützel 2021; Karapunar et al. 2024). Based on a Bayesian phylogenetic analysis (Fossilized Birth-Death model), these authors set the origin time estimation for the clade Pleurotomariida between 460–509 Mya [slightly earlier than the appearance the first observed taxon Clathrospira trochiformis (Portlock 1843) from the Sandbian of Ireland and Scotland] (see Karapunar et al. 2024: fig. 2); thus, the genus Clathrospira (Horný 1997; Wagner 2002) is considered as one of the oldest fossil gastropods belonging to Pleurotomariida recovered in the Late Ordovician. The first occurrence of this genus in the Late Ordovician (Sandbian–Hirnantian) of the Argentine Precordillera supplies additional information enabling to interpret the origin, first appearance and evolution of the entire Pleurotomariida clade in South America during the early Palaeozoic (Fig. 4). We suggest that Clathrospira gondwanica sp. nov. could have been the first precursor of the order Pleurotomariida in Argentina and South America as earlier as Palaeozoic times given rise to the pleurotomariid lineage composed by several genera typical of the late Palaeozoic (Carboniferous–Permian) marine deposits in this region; some of them are Glabrocingulum, Peruvispira, Mourlonia, Ananias, and Borestus (see Pinilla and Taboada 2023). Thus, the report of Clathrospira in the Sandbian–Hirnantian of South America can help to shed light on the origin of the order Pleurotomariida in the southernmost Gondwana continent at that time.
Conclusions
The taxonomic knowledge of most gastropod taxa described so far from the Upper Ordovician of the Argentine Precordillera is outdated in comparison to other coeval invertebrate groups in this region. A more detailed research—including the collection of new gastropod material and the revision of all described gastropod groups of Ordovician age—is currently in progress. However, the new gastropod association of the La Pola and Don Braulio formations provides new data on its taxonomic composition in the Upper Ordovician of Argentina and South America. It clearly shows that the biogeographic pattern of the gastropod genera is consistent with the palaeogeographic trajectory noted for the Precordillera Terrain from the equator in the Cambrian and Early Ordovician moving to intermediate latitudes in the Sandbian, when still maintained the record of some Laurentian forms and with a biotic interchange between Avalonia and Baltica at mid-latitude (30–45°) locations. Then, the Argentine Precordillera exhibited palaeobiogeographical affinities with Morocco (Northern Gondwana) and Avalonia during the Hirnantian, latest Ordovician.
Similar to the conclusions based on other invertebrate groups, gastropods from the Argentine Precordillera still had a Laurentian or low latitude palaeobiogeographical distribution in the Sandbian coincident with a relatively warm paleoclimate, with Clathrospira and Holopea being the single genera occurring in intermediate/high southern latitudes during the Hirnantian. Precordillera also tended to display a Gondwana pattern during the Hirnantian.
The genera Clathrospira and the cosmopolitan Tetranota are represented in the Argentine Precordillera by two new species such as Clathrospira gondwanica sp. nov. and Tetranota argentina sp. nov.; these taxa are here considered as endemic for the Sandbian–Hirnantian intervals. Clathrospira gondwanica sp. nov. is also interpreted as the first precursor of the order Pleurotomariida in Argentina and South America during the Late Ordovician given rise to the pleurotomariid lineage and helping to shed light on the origin of the order Pleurotomariida in the Gondwana continent at that time.
Acknowledgments
We acknowledge support from CONICET (PIP 2021–2023 DD787 Cod F81643) to MGC. We are deeply grateful to Jan-Ove Ebbestad (Museum of Evolution, Uppsala University, Uppsala, Sweden) and David M. Rohr (Department of Earth and Physical Sciences, Sul Ross State University, Alpine, USA for their valuable comments.
References
Aceñolaza, F.G. 1968. Geología estratigráfica de la Region de la Sierra de Cajas. Depto. Humahuaca (Provincia de Jujuy). Revista de la Asociación Geológica Argentina 23: 207–222.
Astini, R.A. 1999. The Late Ordovician glaciation in the Proto-Andean margin of Gondwana revisited: geodynamic implications. In: P. Kraft, O. Fatka (eds.), Quo Vadis Ordovician? Acta Universitatis Carolinae, Geologica 43: 171–174.
Astini, R.A. 2001. Pavimentos estriados en la Formacion Don Braulio y naturaleza de la glaciacion hirnantiana (Ordovícico Tardío) en la region Andina. Revista de la Asociación Argentina de Sedimentología 8: 1–25.
Astini, R.A., Benedetto, J.L., and Vaccari, N.E. 1995. The early Paleozoic evolution of the Argentine Precordillera as a Laurentian rifted, drifted, and collided terrane: A Geodynamic model. Geological Society of America Bulletin 107: 253–273. Crossref
Bassler, R.S. 1915. Bibliographic index of American Ordovician and Silurian Fossils. Volume 2. Bulletin of the United States National Museum 92: 719–1521. Crossref
Benedetto, J.L. 1986. The first typical Hirnantia fauna from South America (San Juan Province, Argentine Precordillera). Biostratigraphie Du Paleozoïque 4: 439–447.
Benedetto, J.L. 1990. Los generos Cliftonia y Paromalomena (Brachiopoda) en el Ashgilliano tardío de la Sierra de Villicum, Precordillera de San Juan, Argentina. Ameghiniana 27: 151–159.
Benedetto, J.L. 2003. Ordovician Fossils of Argentina. Secretaría de Ciencia Y Tecnología. 665 pp. Universidad Nacional de Córdoba, Córdoba.
Benedetto, J.L. 2004. The allochthony of the Precordillera ten years later (1993–2003). A new paleobiogeoraphic test of the microcontinental model. Gondwana Research 7: 1027–1039. Crossref
Benedetto, J.L. 2015. A Middle Ordovician (Darriwilian) dysaerobic brachiopod assemblage from the Precordillera terrane of Argentina: implications for early colonization of deep waters. Ameghiniana 52: 69–106. Crossref
Benedetto J.L., Sánchez, T., Carrera M.G., Brussa, E., and Salas M.J. 1999. Paleontological constraints on successive paleogeographic positions of Precordillera terrane during the early Paleozoic. In: V. Ramos and D. Keppie (eds.), Gondwana-Laurentia Connections Before Pangea. Geological Society of America Special Paper 336: 21–42. Crossref
Benedetto, J.L., Sánchez, T.M., Carrera, M.G., Halpern, K., and Bertero, V. 2011. Faunal shifts and climatic changes in the Upper Ordovician of South America (W Gondwana). In: J.C. Gutiérrez-Marco, I. Rábano, and D. García-Bellido (eds.), Ordovician of the World. Cuadernos del Museo Geominero 14: 55–60.
Benedetto, J.L., Vaccari, N.E, Waisfeld, B.G., Sánchez, T.M., and Foglia, R.D. 2009. Cambrian and Ordovician biogeography of the South American margin of Gondwana and accreted terranes. Geological Society, London, Special Publications 325: 201–232. Crossref
Bertero, V. 2009. First record of Malayaspira Kobayashi, 1958 (Mollusca, Gastropoda) in the lower Ordovician San Juan Formation, Argentine Precordillera. Ameghiniana 46: 567–571.
Bertero,V. 2020. Los gastrópodos de la Precordillera Argentina en el marco de la radiación Ordovícica. 153 pp. Unpublished Ph.D. Thesis, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Córdoba, Argentina.
Bertero, V., Ferrari, M., and Carrera, M.G. 2023. Shell and associated operculum in Teiichispira (Macluritidae: Gastropoda) from the Early/Middle Ordovician of the Argentine Precordillera. Journal of Paleontology 97: 549–557. Crossref
Billings, E. 1865. Paleozoic Fossils, Volume 1, Containing Descriptions and Figures of New or Little Known Species of Organic Remains from the Silurian Rocks 1861–1865. 426 pp. Dawson Brothers, Montreal. Crossref
Blodgett, R.B. and Rohr, D. 2012. Trochonemella, Siskiyouspira, Spinycharydis, Beraunia, Kirkospira, Chulpacispira, Spinulrichospira, Cheeneetnukia, and Odontomaria a potpourri of Alaska’s fancy Paleozoic tropical gastropods. Newsletter of the Alaska Geological Survey 43 (3): 1–7.
Bouchet, P., Rocroi, J.-P., Hausdorf, B., Kaim, A., Kano, Y., Nützel, A., Parkhaev, P., Schrödl, M., and Strong, E.E. 2017. Revised classification, nomenclator and typification of gastropod and monoplacophoran families. Malacologia 61 (1–2): 1–526. Crossref
Boucot, A.J. and Yochelson, E.L. 1966. Paleozoic Gastropoda from the Moose River synclinorium, northern Maine. United States Geological Survey Professional Paper 503 (A): 1–18. Crossref
Brussa, E. 2000. Una nueva asociación de graptolitos caradocianos de la Formación La Cantera, sierra de Villicum, Precordillera Argentina. Ameghiniana 37 (4): 7R.
Brussa, E.D., Toro, B.D., and Benedetto, J.L. 2003. Biostratigraphy. In: J.L. Benedetto (ed.), Ordovician Fossils of Argentina, 75–90. Universidad Nacional de Córdoba, Córdoba, Argentina.
Buggisch, W. and Astini, R. 1993. The Late Ordovician Ice Age: New evidence from the Argentine Precordillera. In: R.H. Findlay, R. Unrug, M.R. Banks, and J.J. Veevers (eds.), Gondwana Eight, Assembly, Evolution, and Dispersal, 439–447. Balkema Editions, Rotterdam.
Cañas, F.L. 1999. Facies and sequences of the Late Cambrian–Early Ordovician carbonates of the Argentine Precordillera: A stratigraphic comparison with Laurentian platforms. In: V. Ramos and D. Keppie (eds.), Gondwana-Laurentia Connections Before Pangea. Geological Society of America, Special Paper 336: 43–62. Crossref
Carrera, M.G. 1994. An Ordovician sponge fauna from the San Juan Formation, Precordillera Basin, western Argentina. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 191: 201–220. Crossref
Carrera, M.G. 2003. The genus Prasopora (Bryozoa) from the Middle Ordovician of the Argentine Precordillera. Ameghiniana 40: 197–203.
Carrera, M.G. and Ernst, A. 2010. Darriwilian bryozoans from the San Juan Formation (Ordovician), Argentine Precordillera. Ameghiniana 47: 343–354. Crossref
Conrad, T.A.1842. Observations on the Silurian and Devonian Systems of the United States, with descriptions of new organic remains. Journal of Academy of Natural Sciences, Philadelphia 8: 228–280.
Cossmann, M. 1908. Revue critique de paléozoologie. Douzième année 1. 288 pp. Auteur et F.R. de Rudeval éd, Paris.
Cox, L.R. and Knight, J.B. 1960. Suborders of the Archaeogastropoda. Proceedings of the Malacological Society of London 33: 262–264.
Cuvier, G. 1797. Table Élémentaire de l’Historie Naturelle de Animaux. 710 pp. Baudouin, Paris. Crossref
Dall, W.H. 1913. Class 4. Gastropoda. Snails. In: C.R. Eastmann (ed.), Textbook of Paleontology, Vol. 1, Second Edition, 514–583. MacMillan & Co., London.
d’Archaic, E.J.A. 1841. Note sur le genre Murchisonia. Bulletin de la Société Géologique de France 12: 154–169.
d’Eichwald, E. 1856. Beitrag zur geographischen Verbreitung der fossilen Tiere Russlands. Alte Periode. Bulletin de la Société Impériale des Naturalistes de Moscou 29: 555–608.
Dzik, J. 2019. Decline in diversity of early Palaeozoic loosely coiled gastropod protoconchs. Lethaia: 32–46. Crossref
Ebbestad, J.O.R. 1998. Multiple attempted predation in the Middle Ordovician gastropod Bucania gracillima. GFF 120: 27–33. Crossref
Ebbestad, J.O.R. 2016. Gastropoda, Tergomya and Paragastropoda (Mollusca) from the Lower Ordovician Fezouata Formation, Morocco. Palaeogeography, Palaeoclimatology, Palaeoecology 460: 87–96. Crossref
Ebbestad, J.O.R. and Peel, J.S. 2001. An unusual trochiform gastropod from the Upper Ordovician of Sweden. Palaeontology 44: 375–387. Crossref
Ebbestad, J.O.R. and Stott, C.A. 2008. Failed predation in Late Ordovician gastropods (Mollusca) from Manitoulin Island, Ontario, Canada. Canadian Journal of Earth Sciences 45 (2): 231–241. Crossref
Ebbestad, J.O.R. and Yochelson, E.L. 2000. Isostrophic molluscs (Tergomya and Gastropoda) from the Upper Ordovician of Norway. Norsk Geologisk Tidsskrift 80: 187–202. Crossref
Ebbestad, J.O.R., Bassett, M.G., Dastanpour, M., and Popov. L.E. 2008. Ordovician (Caradoc) Gastropoda of the Katkoyeh Formation, Kerman Province, Iran. Geobios 41: 605–624. Crossref
Ebbestad, J.O.R., Högström, A.E.S., and Frisk, Å.M. 2013. Gastropods and tergomyans from the Upper Ordovician (Viru-Harju) of the Fågelsång area, Scania, southern Sweden. Journal of Systematic Palaeontology 11: 295–336. Crossref
Ebbestad, J.O.R., Polechová, M., Kröger, B., and Gutiérrez-Marco, J.C. 2019. Late Ordovician molluscs of the central and eastern Anti-Atlas, Morocco. In: A.W. Hunter, J.J. Álvaro, B. Lefebvre, P. Van Roy, and S. Zamora (eds.), The Great Ordovician Biodiversification Event: Insights from the Tafilalt Biota, Morocco. Geological Society, London, Special Publications 485: 237–296. Crossref
Emmons, E. 1842. Geology of New York, Part 2. 437 pp. New York State Geological Survey, Albany.
Ernst, A. and Carrera, M.G. 2008. Cryptostomid bryozoans from the Sassito Formation, Upper Ordovician cool-water carbonates of the Argentinean Precordillera. Palaeontology 51: 1117–1127. Crossref
Ernst, A. and Carrera, M.G. 2012. Upper Ordovician (Sandbian) bryozoan fauna from Argentine Precordillera. Journal of Paleontology 86: 721–752. Crossref
Ernst, A. and Carrera, M.G. 2022. A cool-water bryozoan association from the La Pola Formation (Sandbian, Ordovician) of Argentine Precordillera. Geodiversitas (Bulletin du Muséum National d’histoire Naturelle) 44: 563–600. Crossref
Erwin, D.H. 1990. Carboniferous–Triassic gastropod diversity patterns and the Permo-Triassic mass extinction. Paleobiology 16: 187–203. Crossref
Fernádez-Martínez, E., Carrera, M.G., and Rodríguez, S. 2004. Corales tabulados del Ordovícico (Caradoc) de la Precordillera Argentina. Revista Española de Paleontología 19: 47–59. Crossref
Férussac, A.E.J.P.F. d’Audebard de and d’Orbigny, A. 1840. Histoire naturelle générale et particulière des Céphalopodes acétabulifères vivants et fossiles. 136 pp. Baillière, Paris.
Frisk, Å.M. and Ebbestad, J.O.R. 2007. Paragastropoda, Tergomya, and Gastropoda (Mollusca) from the Upper Ordovician Dalby Limestone, Sweden. GFF 129: 83–99. Crossref
Frýda, J., Rohr, D.M., Robardet, M., and Guitiérrez-Marco, J.C. 2001. A new Late Ordovician microdomatid gastropod genus from Seville, south west Spain, with a revision of Ordovician Microdomatoidea. Alcheringa 25: 117–127. Crossref
Golikov, A.N. and Starobogatov, Y.I. 1975. Systematics of prosobranch gastropods. Malacologia 15: 185–232.
Hall, J. 1847. Paleontology of New York. Vol. 1: Organic Remains of the Lower Division of the New York System (Equivalent of the Lower Silurian Rocks of Europe). 338 pp. C. Van Benthuysen, Albany.
Hall, J. 1852. Paleontology of New York. Vol. 2: Organic Remains of the Lower Middle Division of the New York System. 362 pp. New York State Geological Survey, Albany.
Hall, J. 1859. Contributions to the palaeontology of New York: being some of the results of investigations during the years 1855, 1856, 1857 and 1858. Annual Report of the Regents of the University of the State of New York on the Conditions of the State Cabinet of Natural History and the Historical and Antiquarian Collection Annexed thereto 12: 8–10.
Halpern, K. and Carrera, M. 2014. Post-Glacial Hirnantian (Upper Ordovician) Bryozoans from Western Argentina: Implications for Survival and Extinction Patterns. Ameghiniana 51: 243–253. Crossref
Hammer, Ø. and Harper D.A.T. 2006. Paleontological Data Analysis. 352 pp. Blackwell Publishing, Oxford.
Hammer, Ø., Harper D.A.T., and Ryan P.D. 2001. PAST: Paleontological statistics software package for education and data analysis. Palaeontolgica Electronica 4: 1–9. Crossref
Heredia S. and Milana J.P. 2010. Conodontes sandbianos (Ordovícico Superior) en la Quebrada La Pola, Sierra de Villicum, Precordillera de San Juan (Argentina). Ameghiniana 47: 515–525. Crossref
Hickman, C.S. 1984. Pleurotomaria: Pedigreed perseverance? In: N. Eldredge and S.M. Stanley (eds.), Living Fossils, 225–231. Springer, New York. Crossref
Hisinger, W. 1831. Anteckningar i physik och geognosi under resor uti Sverige och Norrige. Femte Häfte. 174 pp. B.M. Bredberg, Stockholm.
Horný, R.J. 1962. New genera of Bohemian Lower Paleozoic Bellerophontina. Vestnik Ceského Geologického Ústavu 37: 473–476.
Horný, R.J. 1963. Lower Paleozoic Bellerophontina (Gastropoda) of Bohemia. Ustredni ustav geologicky, Sbornik geologickych ved, Paleontologie 2: 57–164.
Horný, R.J. 1996. Secondary shell deposits and presumed mode of life in Sinuites (Mollusca, Gastropoda). Acta Musei Nationalis Pragae, Series B, Historia Naturalis 51: 89–103.
Horný, R.J. 1997. Ordovician Tergomya and Gastropoda (Mollusca) of the Anti-Atlas (Morocco). Acta Musei Nationalis Pragae, Series B (Historia Naturalis) 53: 37–78.
Isakar, M. 1991. Harjuan (Upper Ordovician) new Bivalves and a new Gastropod from North Estonia. Acta et Commentationes Universitatis Tartuensis 12: 43–55.
Isakar, M. 1995. The Platyceratacean gastropod Cyclonema (Cyclonema) hiiumaa Teichert, 1928 from the Llandovery (Silurian) of Estonia. Proceedings of the Estonian Academy of Sciences Geology 44 (3):184–187. Crossref
Isakar, M. and Ebbestad, J.O.R. 2000. Bucania (Gastropoda) from the Ordovician of Estonia. Paläiontologische Zeitschrift 74: 51–68. Crossref
Karapunar, B. and Nützel, A. 2021. Slit-band gastropods (Pleurotomariida) from the Upper Triassic St. Cassian Formation and their diversity dynamics in the Triassic. Zootaxa 5042: 1–165. Crossref
Karapunar B., Höhna, S., and Nützel, A. 2024. Phylogeny of the longest existing gastropod clade (Pleurotomariida) reconstructed with Bayesian and Parsimony methods and its implications on gastropod shell characters. Journal of Systematic Palaeontology 22 (1): 2384141. Crossref
Karapunar, B., Nützel, A., Seuss, B., and Mapes, R.H. 2022. Taxonomy and diversity of slit-band gastropods (order Pleurotomariida) and some slit bearing Caenogastropoda from the Pennsylvanian of the USA. Papers in Palaeontology 8 (2): e1417. Crossref
Kayser, E. 1876. Primordiale und Untersilurische fossilien aus der Argentinischen Republik. Palaeontographica Supplementbände Band SIII: 1–33.
Keller, M. 2012. The Argentine Precordillera: a little American carbonate bank. In: J.R. Derby, R.D. Fritz, S.A. Longacre, W.A. Morgan, and C.A. Sternbach (eds.), The Great American Carbonate Bank: The Geology and Economic Resources of the Cambrian–Ordovician Sauk Megasequence of Laurentia. AAPG Memoir 98: 985–1000.
Keller, M., Buggisch, W., and Lehnert, O. 1998. The stratigraphical record of the Argentine Precordillera and its plate-tectonic background. In: R. Pankhurst and W. Rapella (eds.), The Proto-Andean Margin of Gondwana. Geological Society, Special Publication 142: 35–56. Crossref
Keller, M., Cañas, F., Lehnert, O., and Vaccari, N. 1994. The Upper Cambrian and Lower Ordovician of the Precordillera (Westem Argentina): Some stratigraphic reconsiderations. Newsletters on Stratigraphy 31: 115–132. Crossref
Knight, J.B. 1945. Some new genera of the Bellerophontacea. Journal of Paleontology 19: 333–340.
Knight, J.B. 1956. New families of Gastropoda. Journal of the Washington Academy of Sciences 46: 41–41.
Knight, J.B., Cox, L.R., Keen, A.M., Batten, R.L., Yochelson, E.L., and Robertson, R. 1960. Systematic descriptions. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology, Part 1, Mollusca 1: I169–I339. Geological Society of America and University of Kansas Press, Lawrence, Kansas.
Kobayashi, T. 1931. Studies on the Ordovician stratigraphy and palaeontology of North Korea with notes on the Ordovician fossils of Shantung and Liautung. Bulletin of the Geological Survey of Chosen 11: 1–60.
Kobayashi, T. 1934. The Cambro-Ordovician formations and faunas of south Chosen. Paleontology. Part I. Middle Ordovician faunas. Journal of the Faculty of Science, the Imperial University of Tokyo, Section 2 3 (8): 335–519.
Kobayashi, T. 1935. On the Kainella Fauna of the basal Ordovician age found in Argentina. Japanese Journal of Geology and Geography 12: 59–67.
Kobayashi, T. 1937. The Cambro-Ordovician shelly faunas of South America. Journal Faculty of Sciences, Imperial University of Tokyo 2: 309–522.
Kobayashi, T. 1958. Some Ordovician Fossils from the Thailand-Malayan Borderland. Japanese Journal of Geology and Geography 29: 223–231.
Koken, E. 1896. Die Leitfossilien: ein Handbuch für den Unterricht und für das Bestimmen von Versteinerungen. 848 pp. Chr. Herm. Tauchnitz, Leipzig. Crossref
Koken, E. 1897. Die Gastropoden des baltischen Untersilurs. Bulletin de l’Academie Impériale des Sciences de St.-Pétersbourg Série 7 5: 1–214.
Koken, E. and Perner, J. 1925. Die Gastropoden des Baltischen Untersilurs. Mémoires de l’Académie des Sciences de Russie Série 8, Classe Physico-Mathématique 37: 1–326.
Le Sueur, C.A. 1818. Observations on a new genus of fossil shells. Journal of the Academy of Natural Sciences of Philadelphia 1: 310–313.
Linsley, R.M. 1978. Shell form and evolution of the gastropods. American Science 66: 432–441.
Longstaff, J. 1924. Descriptions of Gastropoda, chiefly in Mrs. Robert Gray’s collection, from the Ordovician and Lower Silurian of Girvan. Quarterly Journal of the Geological Society of London 80: 408–446. Crossref
McCoy, F. 1852. A Synopsis of the Classification of the British Palaeozoic Rocks [by the Rev. Adam Sedgwick] with a Systematic Description of the British Palaeozoic Fossils in the Geological Museum of the University of Cambridge [by Frederick McCoy] with Figures of the New and Imperfectly Known Species. 661 pp. J.W. Parker and Son, London.
Meek, F.B. and Worthen, A.H.1866. Descriptions of invertebrates from the Carboniferous System. Geological Survey of Illinois, Palaeontology 2: 143–411.
Miller, S.A. 1878. Description of a new genus and eleven new species. Journal of Cincinnati Society of Natural History 1: 100–108.
Miller, S.A. and Faber, C.L. 1894. New species of fossils from the Hudson River Group, and remarks upon others. Journal of Cincinnati Society of Natural History 17: 22–33.
Okulitch, V.J. 1935. Fauna of the Black River Group in the vicinity of Montreal. Canadian Field Naturalist 44: 96–107. Crossref
Peralta, S.H. 1986. La graptofauna Ordovícica de la Formación La Cantera, Precordillera Oriental de San Juan. Actas 4 Congreso Argentino de Paleontología y Bioestratigrafía 1: 61–71.
Peralta, S.H. and Carter, C. 1990. La glaciación gondwánica del Ordovícico tardío: Evidencias en fangolitas guijarrosas de la Precordillera de San Juan. Actas 10 Congreso Geológico Argentino (San Juan) 2: 181–185.
Perner, J. 1903. Gastéropodes. Texte (Patellidae et Bellerophontidae) et planches. In: J. Barrande (ed.), Systême silurien du centre de la Bohême Vol. 4, 1–164. Musée Bohême, Prague.
Pickerill, R.K. and Brenchley, P.J. 1979. Caradoc marine benthic communities of the south Berwyn Hills, North Wales. Palaeontology 22: 229–264.
Pinilla, M.K. and Taboada, A.C. 2023. Gastrópodos eopérmicos de la Formación Pampa de Tepuel en el Valle de Tres Lagunas, Chubut, Argentina. Revista del Museo de La Plata 8 (2): 73–86. Crossref
Portlock, J.E. 1843. Report on the Geology of the County of Londonderry, and of Parts of Tyrone and Fermanagh. 784 pp. A. Milliken, Dublin.
Ramos V.A. 2004. Cuyania, an exotic block to Gondwana: review of a historical success and the present problems. Gondwana Research 7: 1–16. Crossref
Rohr, D.M. 1980. Ordovician–Devonian Gastropoda from the Klamath Mountains, California. Palaeontographica A 171: 141–199.
Rohr, D.M. 1988. Upper Ordovician gastropods from the Seward Peninsula, Alaska. Journal of Paleontology 62: 551–566. Crossref
Rohr, D.M. 1996. Ordovician (Whiterockian) Gastropods of Nevada, Part 2. Journal of Paleontology 70: 56–63. Crossref
Rohr, D.M. and Blodgett, R.B. 1985. Upper Ordovician Gastropoda from west-central Alaska. Journal of Paleontology 59: 667–673.
Rohr, D.M., Beresi, M.S., and Yochelson, E.L. 2001. Ordovician gastropods from Argentina. Journal of the Czech Geological Society 46: 3–4.
Rohr, D., Dutro, T., and Blodgett, R.B. 1992. Gastropods and brachiopods from the Ordovician Telsitna Formation, northern Kuskokwim Mountains, west-central Alaska. In: B.D. Webby and J.R. Laurie (eds.), Global Perspective on Ordovician Geology, Proceedings of the Sixth International Symposium on the Ordovician System, University of Sydney, Australia, 499–512. Balkema, Rotterdam.
Rohr, D., Measures, E.A., and Boyce, W.D. 2004. Middle Ordovician (Whiterockian) gastropods from the Table Point Formation, Western Newfoundland. Newfoundland Department of Mines and Energy Geological Survey Report 04-1: 225–234.
Rollins, H.D. 1967. The Phylogeny and Functional Morphology of the Knightitinae, Carinaropsinae, and Praematuratropidae (Gastropoda, Bellerophontacea). 187 pp. Unpublished Ph.D. Thesis, Columbia University, New York.
Safford, J.M. 1869. Geology of Tennessee: Nashville, Term. 550 pp. S.C. Mercer, printer of the State, New York.
Salvini-Plawen, L.V. 1980. A reconsideration of systematics in the Mollusca (phylogeny and higher classification). Malacologia 19: 249–278.
Sánchez, T.M., Benedetto, J.L, and Brussa, E. 1991. Late Ordovician stratigraphy, paleoecology, and sea level changes in the Argentine Precordillera. Advances in Ordovician Geology. Geological Survey of Canada Paper 90 (9): 245–258. Crossref
Sánchez, T.M., Carrera, M., and Waisfeld, B. 2002. Hierarchical factors controlling faunal distribution: A case study from the Ordovician of the Argentine Precordillera. Palaios 17: 309–326. Crossref
Schmidt, F. 1858. Untersuchungen üiber die Silurische Formation von Ehstland, Nord-Livland und Oesel. Archiv für die Naturkunde Liv-, Ehst- und Kurlands, 1st Serie 2: 1–248.
Simroth, H. 1906. Versuch einer neuen Deutung der Bellerophontidae. Sitzungsberichte der Natuforschenden Gesellschaft zu Leipzig 32 (for 1905): 3–8.
Sloan, R.E. and Webers, G.F. 1987. Stratigraphic ranges of Middle and Late Ordovician Gastropoda and Monoplacophora of Minnesota. In: R.E. Sloan (ed.), Middle and Late Ordovician Lithostratigraphy and Biostratigraphy of the Upper Mississippi Valley. Minnesota Geological Survey Report of Investigations 35: 183–186.
Sowerby, J. de C. 1839. Fossil shells in the lowest beds of the Old Red Sandstone and fossil shells of the Upper Ludlow Rock. In: R.I. Murchison (ed.), The Silurian System, 602–604, 608–613. John Murray, London.
Teichert, C. 1928. Stratigraphische und paleontologische Untersuchungen im unteren Gotlandium (Tamsal-Stufe) des westlichen Estlands und der Insel Dagd. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Beilageband 60, Abteilung B: 1–112.
Thompson, E.H. 1970. Morphology and taxonomy of Cyclonema Hall (Gastropoda), Upper Ordovician Cincinnatian Province. Bulletins of American Paleontology 58: 219–284.
Troedsson, G.T. 1918. Om Skånes brachiopodskiffer. Lunds Universitets Årsskrift Ny Följd 2: 1–104.
Twenhofel, W.H. 1928. Geology of Anticosti Island. Geological Survey of Canada, Memoir 154: 1–481. Crossref
Ulrich, E.O. and Scofield, W.H. 1897. The Lower Silurian Gastropoda of Minnesota. The Paleontology of Minnesota 39 (2): 813–1081.
Vaccari, N.E. 1995. Early Ordovician trilobite biogeography of Precordillera and Famatina, western Argentina: Preliminary results. In: J.D, Cooper, M.L. Droser, and S.C. Finney (eds.), Ordovician Odyssey. Short Papers for the Seventh International Symposium on the Ordovician System, 193–196. Pacific Section Society for Sedimentary Geology, Fullerton.
Vanuxem, L. 1842. Geology of New York, Part 3 Comprising Survey of the Third Geological District. 306 pp. W. & A. White and J. Vissche, Albany.
Vostokova, V.A. 1955. Gastropods of the Ordovician of the Leningrad region and the Baltic states [in Russian]. Voprosy Paleontologii 2: 82–124.
Waagen, W.H. 1880. Productus-limestone fossils. Memoir of the Geological Survey of India, Palaeontologica Indica 13: 73–183.
Wagner, P.J. 2002. Phylogenetic relationships of the earliest anisostrophically coiled gastropods. Smithsonian Contributions to Paleobiology 88: 1–152. Crossref
Wahlman, G.P. 1992. Middle and Upper Ordovician symmetrical univalved mollusks (Monoplacophora and Bellerophontina) of the Cincinnati Arch Region, US. Geological Survey Professional Paper 1066: 1–213. Crossref
Waisfeld, B.G., Sanchez, T.M., and Carrera, M.G. 1999. Biodiversification patterns in the Early Ordovician of Argentina. Palaios 14: 198–214 Crossref
Waisfeld, B.G., Sanchez, T.M., Benedetto, J.L., and Carrera, M.G. 2003. Early Ordovician faunal assemblages from western Argentina. Biodiversification trends in different geodynamic and palaeogeographic settings. Palaeogeography, Palaeoclimatology, Palaeoecology 196: 343–373. Crossref
Wenz, W. 1938. Handbuch der Paläozoologie. Band 6: Gastropoda. viii + 240 pp. Borntraeger, Berlin.
Whitfield, R.P. 1878. Preliminary descriptions of new species of fossils from the lower geological formations of Wisconsin. Wisconsin Geological Survey Annual Report 1877: 50–89.
Yochelson, E.L. 1963. The Middle Ordovician of the Oslo Region, Norway. Norsk Geologisk Tidsskrift 43: 133–213.
Yochelson, E.L. and Bridge, J. 1957. The Lower Ordovician gastropod Ceratopea. Geological Survey Professional Paper 294: 281–304. Crossref
Zittel, K.A., von 1895. Grundzüge der Palaeontologie (Palaeozoologie). 971 pp. Oldenbourg, Münich.
Acta Palaeontol. Pol. 69 (4): 747–767, 2024
https://doi.org/10.4202/app.01208.2024