
Cranial anatomy and stratigraphy of a new specimen of the tyrannosaurine dinosaur Daspletosaurus from the Judith River Formation of Central Montana, USA
ETHAN WARNER-COWGILL, GLENN W. STORRS, RAYMOND R. ROGERS, and ANTHONY E. MALTESE
Warner-Cowgill, E., Storrs, G.W., Rogers, R.R., and Maltese, A.E. 2025. Cranial anatomy and stratigraphy of a new specimen of the tyrannosaurine dinosaur Daspletosaurus from the Judith River Formation of Central Montana, USA. Acta Palaeontologica Polonica 70 (1): 159–174.
The tyrannosaurine Daspletosaurus contains three recognized species from the Campanian of Montana and Alberta: Daspletosaurus torosus, Daspletosaurus wilsoni, and Daspletosaurus horneri. The recently named D. wilsoni has been proposed to represent a transitional anagenetic form between D. torosus and D. horneri, a hypothesis contingent on both the stratigraphic succession of these three taxa and the presence of an intermediate morphology in D. wilsoni. Adequate testing of this hypothesis is hampered by limited knowledge of the morphological variation and stratigraphic ranges of both D. wilsoni and D. torosus. We introduce a new, ontogenetically mature specimen of Daspletosaurus from the upper Campanian Coal Ridge Member of the Judith River Formation of central Montana that is well constrained to ~76.3–75.8 Ma. This specimen has a combination of features not yet reported in Daspletosaurus, increasing the known range of morphological disparity within this genus. The cranial morphology and stratigraphic position of this specimen precludes its referral to D. horneri. Although stratigraphically equivalent to D. wilsoni, this specimen lacks one of the three characters purported to distinguish that taxon from D. torosus (dorsal quadrate process of quadratojugal broadly visible laterally). We propose that this character is intraspecifically variable within Daspletosaurus and therefore not diagnostic, thus weakening the case that D. wilsoni is distinct from D. torosus. Additional specimens with stratigraphic controls are necessary to determine if D. wilsoni is a valid taxon.
Key words: Dinosauria, Theropoda, Tyrannosauridae, Daspletosaurus, anagenesis, Late Cretaceous, Campanian, Laramidia, Judith River Formation.
Ethan Warner-Cowgill [plateaupaleo@gmail.com; ORCID: https://orcid.org/0009-0004-9935-274X ] and Glenn W. Storrs [gstorrs@cincymuseum.org; ORCID: https://orcid.org/0000-0001-8542-1557 ], Cincinnati Museum Center, 1301 Western Avenue, Cincinnati, OH 45203, USA.
Raymond R. Rogers [rogers@macalester.edu; ORCID: https://orcid.org/0000-0002-1557-2058 ], Macalester College, 1600 Grand Ave, St Paul, MN 55105, USA.
Anthony E. Maltese [anthony@rmdrc.com; ORCID: https://orcid.org/0009-0002-0066-9399 ], Rocky Mountain Dinosaur Resource Center, 201 S Fairview St, Woodland Park, CO 80863, USA.
Received 15 February 2024, accepted 25 October 2024, published online 24 March 2025.
Copyright © 2024 E. Warner-Cowgill et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Tyrannosauroidea was a successful radiation of Laurasian coelurosaurian theropods that originated in the Jurassic as small-bodied forms and ultimately gave rise to the Tyrannosauridae, a clade characterized by large to gigantic body size, enlarged skulls, and famously reduced forelimbs (Brusatte et al. 2010). Tyrannosaurids were the dominant terrestrial predators of Asia and Western North America (Laramidia) in the final Campanian and Maastrichtian stages of the Cretaceous (Loewen et al. 2013). Their rich fossil record and popular appeal have inspired a wealth of research and speculation relating to their life appearance (e.g., Cullen et al. 2023), ontogeny (e.g., Carr 1999), ecology (e.g., Farlow et al. 2023), behavior (e.g., Witmer and Ridgley 2009), biomechanics (e.g., Gignac and Erickson 2017), biogeography (e.g., Thomson et al. 2013), evolutionary tempo and mode (e.g., Warshaw and Fowler 2022), and phylogenetic relationships (e.g., Loewen et al. 2013). Two subfamilies of tyrannosaurids are recognized, the Albertosaurinae represented by Gorgosaurus and Albertosaurus from the upper Campanian and lower Maastrichtian of Alberta, respectively, and the Tyrannosaurinae, the last surviving clade of tyrannosaurids that includes the gracile alioramins as well as Tyrannosaurus rex and its close relatives such as Daspletosaurus and Tarbosaurus (Brusatte and Carr 2016).
In the more than half-century that has passed since the inaugural description of Daspletosaurus torosus Russell, 1970, surprisingly little descriptive work has been conducted on this species (although see Paulina-Carabajal et al. 2021). Recent discoveries have led to the recognition of two new species of Daspletosaurus, D. horneri Carr et al., 2017, and D. wilsoni Warshaw & Fowler, 2022, from the Two Medicine and Judith River formations of Montana, respectively (Fig. 1). Horner et al. (1992) hypothesized that D. torosus, D. horneri (then unnamed), and Tyrannosaurus rex represented a successive evolutionary series that evolved through anagenesis. Anagenesis can be supported if the taxa in question are close phylogenetic relatives, do not overlap stratigraphically, and occupy similar geographic ranges (Carr et al. 2017; Zietlow 2020). Carr et al. (2017) rejected an anagenetic relationship between Daspletosaurus and Tyrannosaurus, but found support for evolution through anagenesis between D. torosus and D. horneri. Warshaw and Fowler (2022) introduced D. wilsoni as a transitional form that evolved from D. torosus and into D. horneri through anagenesis (D. torosus → D. wilsoni → D. horneri), a claim partly contingent on the hypothesized lack of temporal overlap between these taxa, with D. torosus at ~77 Ma, D. wilsoni at ~76.5 Ma, and D. horneri at ~75.1 Ma (Warshaw and Fowler 2022). A subsequent study found that the phylogenetic position of each species conflicted with their stratigraphic position, indicating the existence of multiple contemporary lineages of Daspletosaurus rather than a series of successive sister taxa (Scherer and Voisculescu-Holvad 2024). Another phylogenetic analysis (Warshaw et al. 2024) incorporating additional specimens, however, found additional support for the phylogenetic results of Warshaw and Fowler (2022) and, by extension, the hypothesis that Daspletosaurus evolved through anagenesis.
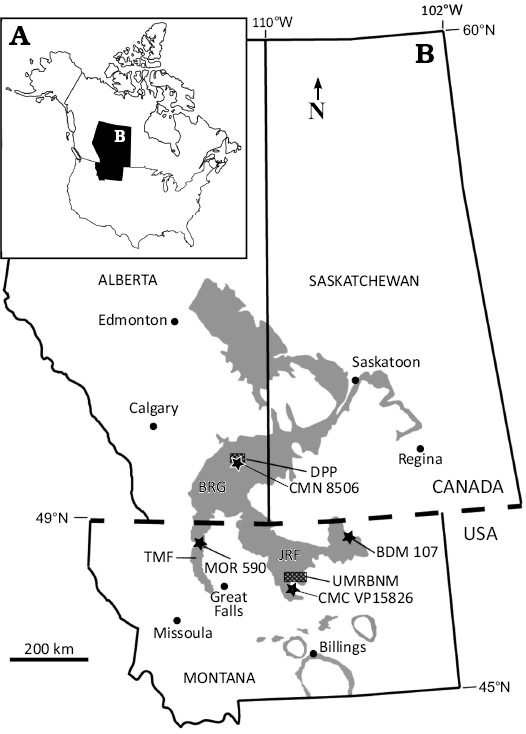
Fig. 1. A. Location of the study area within North America. B. Location of Daspletosaurus sp. (CMC VP15826) in the Judith River Formation (JRF) of Montana. The new specimen was preserved in JRF strata just south of the Upper Missouri River Breaks National Monument (UMRBNM). Additional specimens discussed in the text include Daspletosaurus torosus (CMN 8506) from the Oldman Formation, Belly River Group (BRG) in Dinosaur Provincial Park (DPP), Daspletosaurus wilsoni (BDM 107) from JRF exposures north and east of the UMRBNM, and Daspletosaurus horneri (MOR 590) from the Two Medicine Formation (TMF) in northwestern Montana (Rogers et al. 2025). Gray color represents the outcrop belt of Campanian terrestrial strata in the region. Modified from Rogers et al. (2024).
Here we report the discovery of a new specimen of Daspletosaurus from the Judith River Formation in central Montana. At ~76.3–75.8 Ma, the new specimen is chronologically older than the oldest occurrence of D. horneri (~75.2 Ma), and younger than the holotype of D. torosus (~76.8 Ma), falling within the temporal range of D. wilsoni (~76.5–75.6 Ma) proposed by Warshaw et al. (2024: fig. 6). This specimen can be excluded from D. horneri based on cranial morphology, but has numerous features present in both D. torosus and D. wilsoni. This specimen shares one character state (dorsal quadrate process of quadratojugal not broadly visible laterally) with D. torosus to the exclusion of D. wilsoni. If this character is to be considered diagnostic, this specimen cannot be assigned to the contemporary D. wilsoni, thereby weakening the hypothesis that Daspletosaurus consists of a single anagenetic lineage. Because this character state is apparently variable in D. horneri, however, and can only be assessed in one specimen of D. torosus (CMN 8506), we instead suggest that this character is intraspecifically variable in Daspletosaurus and therefore insufficient for distinguishing D. wilsoni from D. torosus. If the new specimen is assigned to D. wilsoni, therefore, only two characters (anteroposteriorly narrow orbit, “inflated rostrodorsal ala” of lacrimal) may distinguish D. wilsoni from the holotype of D. torosus.
Institutional abbreviations.—AMNH, American Museum of Natural History, New York, USA; BDM, Badlands Dinosaur Museum, Dickinson, USA; CMC, Cincinnati Museum Center, USA; CMN, Canadian Museum of Nature, Ottawa, Canada; FMNH, Field Museum of Natural History, Chicago, USA; MOR, Museum of the Rockies, Bozeman, USA; NHMUK, Natural History Museum of the United Kingdom, London, UK; RSM, Royal Saskatchewan Museum, Regina, Saskatchewan, Canada; TMP, Royal Tyrrell Museum, Drumheller, Canada; UALVP, University of Alberta Laboratory for Vertebrate Paleontology, Edmonton, Canada; UMNH, Natural History Museum of Utah, Salt Lake City, USA; UWBM, University of Washington, Burke Museum of Natural History and Culture, Seattle, USA.
Geological setting
The dinosaur assemblage of the Judith River Formation has become better known in recent years due to a resurgence of fieldwork and collecting on both public and private lands as well as reanalysis of historical collections. Ornithischian dinosaur taxa currently recognized from the Judith River Formation include the hadrosaurids Brachylophosaurus canadensis (Cuthbertson and Holmes 2010), Corythosaurus sp. (Takasaki et al. 2022), Probrachylophosaurus bergei Freedman-Fowler & Horner, 2015, the ceratopsids Spiclypeus shipporum Mallon et al., 2016, Judiceratops tigris Longrich, 2013, Mercuriceratops gemini Ryan et al., 2014, Lokiceratops rangiformis Loewen et al., 2024, Medusaceratops lokii Ryan et al., 2010, Avaceratops lammersi Dodson, 1986, Furcatoceratops elucidans Ishikawa et al., 2023, and the potentially dubious Monoclonius crassus Cope, 1876, and Ceratops montanus Marsh, 1888, as well as the ankylosaurid Zuul crurivastator Arbour & Evans, 2017. Theropods currently documented in the Judith River assemblage include the dromaeosaurid Saurornitholestes (Wilson and Fowler 2020) and the tyrannosaurids Gorgosaurus sp. (Dalman and Lucas 2015) and Daspletosaurus wilsoni Warshaw & Fowler, 2022.
The new specimen (CMC VP15826) was collected from the Coal Ridge Member of the Judith River Formation (Rogers et al. 2016) on private ranchland near Upper Missouri River Breaks National Monument south of the Missouri River in Fergus County, north-central Montana in 2006. The skeleton was located close to the surface and was deeply penetrated by plant roots. Articulated and closely associated skeletal remains were preserved within a gray fine-grained silty sandstone with mudstone intraclasts immediately above the base of the bone-bearing horizon in a deposit interpreted as a crevasse splay by Maltese (2009). The softness of the matrix and the minimal amount of overburden resulted in extensive weathering and degradation of the specimen, which sometimes obscures details of morphology. Nevertheless, enough anatomical information survives to allow for adequate description and comparison with other specimens.
Deposits of the Coal Ridge Member both entomb the specimen and crop out above the quarry site (Fig. 2). Carbonaceous mudstones, siltstones, and fine-grained sandstones typify the unit in the local outcrop belt, and thin cm-scale beds of orange ironstone are developed along some bed contacts. Approximately 13 m separate the new specimen from the base of the overlying Woodhawk Member. The sharp contact between members is underlain by carbonaceous brown sandy mudstone and capped by fine-grained gray to tan sandstone (Fig. 2C). Small vertical burrows (3–5 mm diameter) attributable to Skolithos occur in the basal meter of the Woodhawk Member. Overall exposure is rather limited on the weathered and vegetated slopes that rise above the site, but indurated outcrop near the top of the ridge exhibits hummocky and swaley bedding consistent with deposition on a shallow marine shelf impacted by storm waves (Fig. 2D). Exposures of the Woodhawk Member continue to the top of local exposure, and the contact with the overlying Bearpaw Formation has not been identified in the immediate vicinity of the site.

Fig. 2. View of Daspletosaurus sp. site and associated strata in upper Judith River Formation. A. Drone view of Daspletosaurus sp. site in Coal Ridge Member. The fossil site is located ~15 m below the contact with the marine Woodhawk Member (contact marked by white dashed line). Locations of rocks featured in images C and D are indicated. B. View from the Daspletosaurus sp. site looking south toward exposures of Coal Ridge and Woodhawk members. C. Contact between Coal Ridge and Woodhawk members on ridge adjacent to fossil site. D. View of hummocky and swaley bedding in Woodhawk Member near top of ridge.
The age of CMC VP15826 can be approximated in the context of the local stratigraphy, which was recently calibrated with a suite of new U-Pb zircon ages by Ramezani et al. (2022). Significantly, the site occurs without question in the Coal Ridge Member, which is positioned above the Judith River-Belly River discontinuity. This chronostratigraphically significant discontinuity, which can be tracked from north-central Montana to the environs of Dinosaur Provincial Park in southern Alberta, has been dated to ~76.3 Ma (Rogers et al. 2016, 2024). Thus, we can confidently conclude that the specimen is younger than ~76.3 Ma.
The age of CMC VP15826 can be further refined using a Bayesian age model recently developed for the Judith River Formation by Ramezani et al. (2022: fig. 5). To apply this model and assess the age of the specimen, it is essential to know the stratigraphic distance between the Daspletosaurus sp. site and a bounding contact of the Judith River Formation, in this case the top of the formation. Unfortunately, as clarified above, the upper contact of the Judith River Formation is not exposed in the local outcrop belt where the site occurs, but the critical distance can be approximated by tracking the base of the Woodhawk Member, which can be tied to the top of the formation with reasonable precision in nearby exposures along the Missouri River corridor (Fig. 3A).
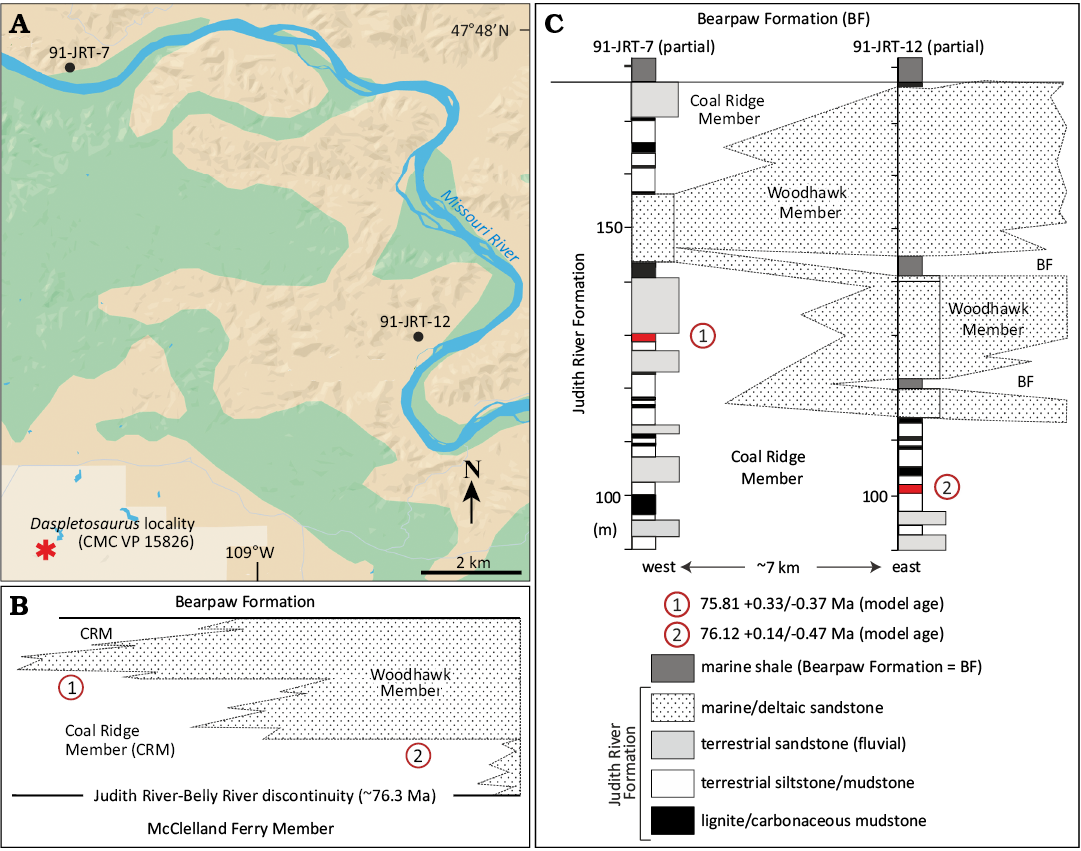
Fig. 3. A. Map view of site, which is in the eastern portion of the Judith River Formation type area. Locations of stratigraphic logs featured in C are indicated. B. Schematic cross section of stratigraphy in vicinity of Daspletosaurus sp. site. Marine sandstones of the Woodhawk Member crop out widely in this part of the type area. This marine unit thins to the west, eventually pinching out and passing laterally to coastal plain facies of the Coal Ridge Member. The approximate position of the Judith River-Belly River discontinuity and underlying McClelland Ferry Member are also shown. C. Nearby stratigraphic logs measured along the Missouri River corridor help contextualize the Daspletosaurus sp. site relative to the overlying Woodhawk Member, the base of which is ~13 m above the site. Two alternative placements of the Daspletosaurus sp. site are indicated by numbered red circles. Given that the Woodhawk Member thins to the west, and given that the site is ~7 km west of section 91-JRT-12, we contend that alternative 1, which yields a model age of 75.81 +0.33/-0.37 Ma, is a more reliable approximation of the age of the Daspletosaurus sp. site (see text for additional discussion). Model ages from Ramezani et al. (2022). Map in A based on Google Maps. Abbreviation: BF, Bearpaw Formation.
Existing evidence indicates that the Woodhawk Member thins to the west, eventually pinching out and passing laterally to the terrestrial Coal Ridge Member. At the unit’s stratotype locality (section 91-JRT-12, see Rogers et al. 2016: fig. 3), which is located ~8 km to the north and east of the Daspletosaurus sp. site (Fig. 3A), approximately 60 m of the sandstone-dominated Woodhawk Member are overlain by the Bearpaw Formation, and ~30 m of Coal Ridge strata underlie the unit. Here the new Daspletosaurus sp. site would presumably occur ~75 m beneath the Judith River-Bearpaw contact (Fig. 3B, C), yielding a model age of 76.12 +0.14/-0.47 Ma (Ramezani et al. 2022). We view this outcome as a maximum model age for the occurrence of CMC VP15826. Shifting westward approximately 7 km to section 91-JRT-7 (see Fig. 3A), the Woodhawk Member has climbed stratigraphically by ~30 m, thinned to ~15 m in thickness, and is bounded above and below by heterolithic facies of the Coal Ridge Member (Fig. 3B, C). Section 91-JRT-7 is positioned roughly along strike with the Daspletosaurus sp. quarry (see Fig. 3A), and here the site would fall roughly 48 m beneath the Judith River-Bearpaw contact, yielding a model age of 75.81 +0.33/-0.37 Ma (Ramezani et al. 2022). We consider section 91-JRT-7 a more appropriate proxy for the local section hosting the new Daspletosaurus sp. skeleton given the along-strike relationship of the site and the section. Moreover, the subdued nature of the Woodhawk outcrop on the ridge above the Daspletosaurus sp. site is consistent with the thinning of the unit and its intercalation within Coal Ridge strata. The model age of 75.81 +0.33/-0.37 Ma is therefore deemed a reliable approximation of the specimen’s age.
Systematic paleontology
Dinosauria Owen, 1842
Theropoda Marsh, 1881
Tetanurae Gauthier, 1986
Coelurosauria Huene, 1914
Tyrannosauridae Osborn, 1906
Tyrannosaurinae Osborn, 1906
Daspletosaurini Voris et al., 2020
Genus Daspletosaurus Russell, 1970
Type species: Daspletosaurus torosus Russell, 1970; Dinosaur Provincial Park, Q72, Alberta, Canada; Oldman Formation, Upper Cretaceous.
Daspletosaurus sp.
Figs. 4–10.
Material.—CMC VP15826, partial skull and nearly complete postcranium from the Upper Cretaceous (Late Campanian) Coal Ridge Member of the Judith River Formation, Fergus County, Montana.
Description.—The specimen CMC VP15826, is approximately 80% complete, was partially articulated when discovered, and preserves portions of the skull and most of the vertebral column (Fig. 4). The skull and cervical vertebrae were situated at the erosional edge of the specimen where preservation was poor. No tooth bearing elements other than a fragment of the left maxilla are present nor is the braincase or skull roof, but bones of the suspensoria and palate are preserved. A severely weathered portion of the posterior mandible may be present, although it is too poorly preserved to be described. The postcranial skeleton is very well-represented. Only three cervical vertebrae and a dozen or so caudal vertebrae are lacking, the latter primarily the distalmost elements. Most of the ribs and chevrons are present as is most, if not all, of the gastral basket. The pectoral girdle is complete with both scapulocoracoids, as well as the furcula. The forearms are similarly preserved, lacking only the ungual phalanges. Both ilia and ischia are preserved, but not the pubes. The left leg lacks only pedal phalanx IV-1 and most of metatarsal III; the right leg was not recovered.
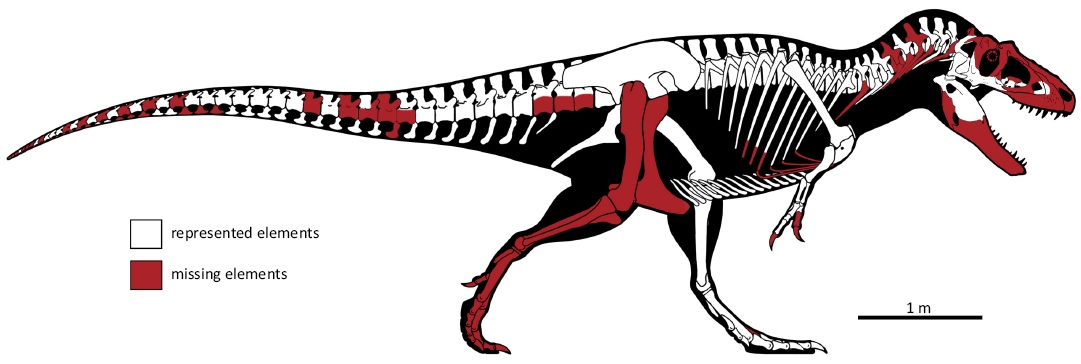
Fig. 4. Skeletal reconstruction of the tyrannosaurine dinosaur Daspletosaurus sp. (CMC VP15826) from the Upper Cretaceous (Late Campanian) Coal Ridge Member of the Judith River Formation, Fergus County, Montana. Modified from original drawing by Scott Hartman (University of Wisconsin, Madison, USA), courtesy of Cincinnati Museum Center.
CMC VP15826 is very large, with a reconstructed total body length greater than 9 meters. The ontogenetic maturity of this specimen is attested to not only by its size but also by the complete fusion of almost all neurocentral sutures down to the most distally preserved caudal vertebrae, fused scapulae and coracoids, and in particular the extreme rugosity of the bone surfaces of the neural spines. The distal ends of the spines, and especially the anterior and posterior surfaces of the spines are heavily textured with hyperostoses reflecting partial ossification of the spinous ligaments. Similar patterning is present at the distal ends of the transverse processes.
CMC VP15826 is assigned to Daspletosaurus by the presence of two cranial synapomorphies: (i) a deep keel on the ventral surface of the vomer and (ii) a posterior pneumatic recess of the palatine located posterior to the anterior margin of the vomeropterygoid neck (Carr et al. 2017; Voris et al. 2020). CMC VP15826 further agrees with Daspletosaurus and differs from the daspletosaurin Thanatotheristes in having a mediolaterally thin suborbital margin of the jugal (Voris et al. 2020). CMC VP15826 can be excluded from D. horneri (Carr et al. 2017) by the presence of a maxillary fenestra that is anteriorly tapered, longer than tall, and dorsally offset from the ventral margin of the antorbital fossa, a ventral pneumatic recess of the squamosal that is not entirely undercut, the absence of a pneumatic foramen on the neck of the quadratojugal, the presence of a dorsal quadratojugal process of the jugal positioned lateral to the ventral quadratojugal process, an inflation of the jugal that extends below the pneumatic recess, and a humerus-femur length ratio of ~0.37 (Carr et al. 2017). CMC VP15826 differs from D. wilsoni (sensu Warshaw and Fowler 2022; Warshaw et al. 2024) and agrees with D. torosus (CMN 8506) in having a medially directed dorsal quadrate process of the quadratojugal.
Because D. horneri and D. wilsoni have been primarily distinguished from D. torosus by their skulls, the present study focuses on, but is not limited to, cranial elements. A comprehensive description of the well-represented postcranial skeleton of CMC VP15826 is beyond the scope of this paper but will be the subject of a future study. A table of measurements of the postcranial skeleton is provided in SOM (Supplementary Online Material available at http://app.pan.pl/SOM/app70-WarnerCowgill_etal_SOM.pdf).
A humerus-to-femur length ratio of ~0.34 has been considered autapomorphic of D. horneri (Carr et al. 2017). The holotype of D. torosus does not preserve a femur, so Carr et al. (2017) used the length of the skull and the ilium as a proxy and found ratios of 0.34 and 0.32, respectively. The femur of CMC VP15826 is 97 cm long as measured from the proximal surface to the medial condyle. The right humerus is 34 cm, whereas the left humerus is 38 cm, a difference that is probably a result of postmortem distortion. By averaging the length of both humeri, the humerus to femur length ratio in the present specimen is ~0.37, a ratio most similar to Gorgosaurus libratus as described by Lambe (1917), but much higher than Tyrannosaurus rex (0.29, Brochu 2003) and Tarbosaurus bataar (0.26, Carr et al. 2017). The ilia are 108 cm (right) and 111 cm (left) in length, providing a femur-to-ilium length ratio of 0.87, and a humerus-to-ilium length ratio of ~0.33. Carr et al. (2017) report that D. horneri has stout epipophyses that either just reach or only slightly project past the postzygapophyses, whereas D. torosus is coded in that study as having long epipophyses. Though incompletely preserved, the epipophyses on the anterior-mid cervicals of CMC VP15826 are short and do not reach the postzygapophyses, reflecting the reported condition in D. horneri. Based on unpublished photographs of the original specimens, we have independently verified the observations of Carr et al. (2017) regarding the morphology of the cervical epipophyses in D. horneri and the holotype of D. torosus. Additional specimens of D. torosus are, however, necessary to determine if the presence of long epipophyses is taxonomically informative or intraspecifically variable.
Maxilla: Although only a fragment of the left maxilla is present (Fig. 5A), important features are preserved. The fragment represents the area immediately surrounding the anterior part of the maxillary fenestra. As in other specimens of Daspletosaurus, the promaxillary fenestra is hidden from lateral view and the maxillary fenestra was large. Although the absolute size of the maxillary fenestra is not known because the interfenestral strut is not preserved, it was greater than 98 mm in length and greater than 78 mm in height. The maxillary fenestra is anteriorly tapered as in D. torosus and D. wilsoni (Warshaw and Fowler 2022), unlike the round condition in D. horneri (Carr et al. 2017). As in D. torosus and D. wilsoni the maxillary fenestra of CMC VP15826 is dorsally offset from the ventral margin of the antorbital fossa whereas in subadult (MOR 590) and adult (MOR 1130) D. horneri the fenestra is positioned closer to the ventral margin of the antorbital fossa. The promaxillary fenestra measures ~13 mm in width and is ~43 mm long and has a ventral margin located slightly below the anterior corner of the maxillary fenestra. The presence of anastomosing sulci on the subcutaneous surface of the maxilla, a synapomorphy of Daspletosaurus (Carr et al. 2017), cannot be assessed in CMC VP15826 due to poor preservation.

Fig. 5. Tyrannosaurine dinosaur Daspletosaurus sp. (CMC VP15826) from the Upper Cretaceous (Late Campanian) Coal Ridge Member of the Judith River Formation, Fergus County, Montana. A. Fragment of left maxilla in lateral (A1, A2) and medial (A3) views. B. Left jugal in medial (B1) and lateral (B2) views. Abbreviations: acp, accessory cornual process; af, antorbital fossa; dqp, dorsal quadratojugal process; lc, lacrimal contact; li, lateral inflation; lmp, lateral maxillary process; mf, maxillary fenestra; pf, promaxillary fenestra; por, postorbital ramus; pr, pneumatic recess; rmaf, reconstructed margin of antorbital fossa; rmmf, reconstructed margin of maxillary fenestra; ss, subcutaneous surface; vcp, ventral cornual process; vqp, ventral quadratojugal process.
Jugal: The left jugal (Fig. 5B) is nearly complete and ca. 480 mm long, but taphonomic distortion has left the distal portion of the postorbital ramus severely crushed, and the entire ramus bent posteriorly. This may be explained by the fact that the jugal was the only cranial element that was oriented vertically in the quarry during excavation. This element aligns with D. torosus rather than D. horneri in that the lateral inflation extends below the pneumatic recess, the ventral quadratojugal process is positioned medial to the dorsal quadratojugal process, and the lateral maxillary process is visible in medial view (Carr et al. 2017).
As in D. torosus and D. wilsoni the ventral margin of the orbit is extremely thin mediolaterally as opposed to the thin but rounded margin of Lythronax argestes and the wide rounded margin in Thanatotheristes degrootorum (Voris et al. 2020; Warshaw and Fowler 2022). The posterior portion of the lacrimal contact is very shallowly inclined as in D. wilsoni and D. torosus (Warshaw and Fowler 2022), whereas this surface is very steep in D. horneri, albertosaurines (Currie 2003), and Lythronax argestes. Anterior to the orbit the jugal is directed straight anteriorly as in D. torosus and D. wilsoni (Warshaw and Fowler 2022).
The jugal of CMC VP15826 differs from the holotypes of D. horneri, D. wilsoni, and D. torosus in a few ways. The suborbital height of the jugal is much more dorsoventrally constricted than in the holotypes of D. torosus, D. wilsoni, and D. horneri. However, the same is also true for TMP 2001.36.1, a complete skull from the Oldman Formation of Alberta provisionally identified as D. torosus by Voris et al. (2019: fig. 6B), a new taxon by Paulina-Carabajal et al. (2021), and most recently referred to D. wilsoni by Warshaw et al. (2024). The antorbital margin of the jugal in both TMP 2001.36.1 and CMC VP15826 also share a more shallowly inclined dorsal margin than in D. horneri (MOR 590 and MOR 1130), or the holotypes of D. torosus (CMN 8506) and D. wilsoni (BDM 107). The proximal portion of the postorbital ramus of the jugal in both TMP 2001.36.1 and CMC VP15826 is relatively narrow anteroposteiorly, whereas in D. horneri, CMN 8506, and BDM 107 it is anteroposteriorly broad. Whereas D. torosus (CMN 8506) and D. wilsoni (BDM 107) have a large pneumatic opening on the maxillary ramus of the jugal (pneumatic recess, Fig. 5B), in both CMC VP15826 and TMP 2001.36.1 they are reduced in size. The extreme reduction in size of the pneumatic recess in CMC VP15826 may be the result of an overgrowth of bone reflecting the advanced ontogenetic stage of this individual. A laterally protuberant accessory cornual process is present posterior to and above the primary ventral cornual process. This process is located well anterior to the quadratojugal contact and is therefore distinct from the knob that is autapomorphic of Teratophoneus curriei (Carr et al. 2011). This process is undercut and has a rounded, convex surface. Such an accessory cornual process has not been reported in other specimens of Daspletosaurus and may be pathological.
Squamosal: The left squamosal (Fig. 6) is largely complete and 216 mm in maximum length, although the anteroventral edge of the quadratojugal process is missing. In the absence of the postorbitals, it cannot be positively stated that the squamosal terminates posterior to the anterior margin of the laterotemporal fenestra, a synapomorphy of Daspletosaurus (Carr et al. 2017). However, the squamosal of CMC VP15826 does not significantly differ from D. torosus in any important respect. As in D. torosus and the holotype of D. wilsoni (Warshaw and Fowler 2022), the anteromedial margin of the ventral pneumatic recess of the squamosal is not undercut, whereas this margin is autapomorphically undercut in D. horneri (Carr et al. 2017). A large foramen on the medial surface of the posterior process indicates that this process was pneumatic as in all species of Daspletosaurus.
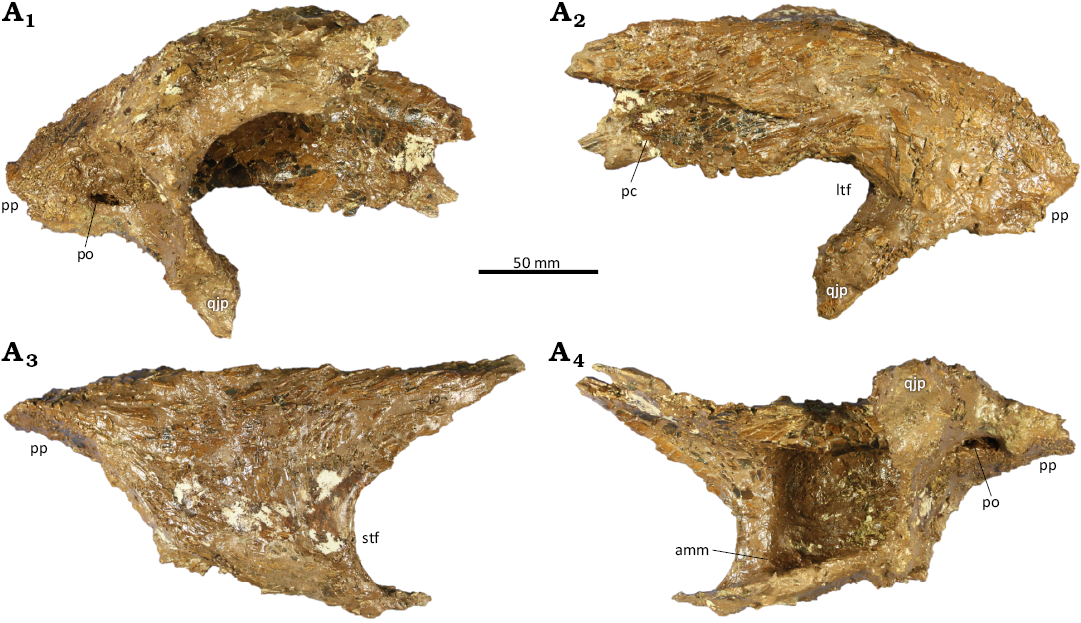
Fig. 6. Tyrannosaurine dinosaur Daspletosaurus sp. (CMC VP15826) from the Upper Cretaceous (Late Campanian) Coal Ridge Member of the Judith River Formation, Fergus County, Montana. Left squamosal in medial (A1), lateral (A2), dorsal (A3), and ventral (A4) views. Abbreviations: amm, anteromedial margin of ventral pneumatic recess; ltf, laterotemporal fenestra; pc, postorbital contact; po, pneumatic opening; pp, posterior process; qjp, quadratojugal process; stf, supratemporal fenestra.
Quadratojugal: Both quadratojugals (Fig. 7) are preserved, but the right is complete and well preserved, whereas the left is lacking half of its anterior (jugal) ramus and the process forming the dorsal quadrate process. The maximum dorsoventral height of the right quadratojugal is 217 mm and the length along its ventral margin is 180 mm. The quadratojugals agree with the holotype of D. torosus and differ from the holotypes of D. horneri and D. wilsoni in having a posteromedially directed, rather than a posterolaterally directed dorsal quadrate process. Unlike D. horneri, the anterior ramus is oriented anteroventrally rather than anterodorsally. It agrees with D. torosus and D. wilsoni but differs from D. horneri (MOR 1130) in lacking a pneumatic foramen on the lateral surface of the neck and in having a deeply notched ventral quadrate process. CMC VP15826 shares one feature in common with D. horneri absent from the holotypes of D. torosus (CMN 8506) and D. wilsoni (BDM 107); the facet for the dorsal jugal process is partly obscured from lateral view whereas in CMN 8506 and BDM 107 this facet is entirely exposed in lateral view (Warshaw and Fowler 2022; Fig. 7A2). The anterior margin of the anterior ramus of the quadratojugal is squared off, rather than tapered or forked.

Fig. 7. Tyrannosaurine dinosaur Daspletosaurus sp. (CMC VP15826) from the Upper Cretaceous (Late Campanian) Coal Ridge Member of the Judith River Formation, Fergus County, Montana. A. Right quadratojugal in medial (A1) and lateral (A2) views. B. Right quadrate of in lateral (B2) and medial (B1) views. C. Left quadrate in lateral (C1) and medial (C2) views. Abbreviations: djc, dorsal jugal contact; dqp, dorsal quadrate process; jr, jugal ramus; mc, mandibular condyles; mf, medial fossa; pf, pterygoid flange; qf, quadrate foramen; sc, squamosal contact; vqc, ventral quadrate contact.
Quadrate: Both left and right quadrates (Fig. 7B) are complete and relatively well preserved despite some transverse crushing. They average 242 mm in maximum dorsoventral height, 290 mm in maximum length through the pterygoid flange (“orbital process” of Carr et al. 2017), and 144 mm in maximum mediolateral width across the mandibular condyles. Among species of Daspletosaurus, only the quadrates of D. wilsoni have been adequately figured, limiting comparisons between specimens. The quadrates of CMC VP15826 do not appear to differ markedly from the holotype of D. wilsoni (Warshaw and Fowler 2022: fig. 9) but do differ from D. horneri in lacking fossae ventral to the anterior pneumatic recess, in lacking an undercut posteroventral margin of the medial fossa of the pterygoid flange, and for having a pneumatic recess that does not reach the medial surface of the pterygoid flange (Carr et al. 2017).
The left and right quadrates of CMC VP15826 are asymmetric with respect to one another in several ways. In the right quadrate the dorsal surface of the pterygoid flange is straight for most of the length of the flange, whereas in the left quadrate this margin is deflected sharply ventrally at the anterior third of its length similar to the condition in D. wilsoni (BDM 107). A groove is reportedly present between the articular condyles of D. torosus, but not in D. horneri (Carr et al. 2017). In CMC VP15826 there is a groove on the right quadrate only, with the condyles being divided instead on the left quadrate by a broad concavity. The appearance of a groove on the right quadrate may be a result of postmortem distortion, however. The angle of the anteroventral margin of the pterygoid flange of the left quadrate appears to be essentially identical to that of D. wilsoni, though this angle is reportedly steeper in D. horneri than in D. torosus (Warshaw and Fowler 2022; Carr et al. 2017). In the present specimen, however, this margin is considerably less steep in the right quadrate. Such asymmetries in a single individual may indicate a high degree of plasticity in the morphology of this element.
Vomer: The fused vomers (Fig. 8) are nearly complete anterior to the point where both the left and right sides of the element come together, and approximately 446 mm long as preserved. Though twisted and crushed, it is largely consistent with the morphology of other tyrannosaurids (Brochu 2003; Currie 2003). The left and right sides together enclose an elongated groove on the posterior half of the dorsal surface of the element. The anterior plate is broad relative to the mediolateral width of the stem, but not to the extraordinary degree of lateral expansion present in Tyrannosaurus rex (Molnar 1991; Larson 2008). No foramina are present within the anterior plate as there are in some specimens of Tyrannosaurus rex (Larson 2008). As in Tyrannosaurus rex, there is a mediolaterally narrow, dorsoventrally flat process that continues anterior to the anterior plate and that forms the anterior limit of the element (Brochu 2003), although it appears to be incomplete in the present specimen. A deep keel is present at the midline of the element’s ventral surface, a synapomorphy of Daspletosaurus sp. (Carr et al. 2017). A keel is also present on the ventral side of the vomer in Tyrannosaurus rex below the posterior end of the anterior plate, whereas in Daspletosaurus sp. this keel is positioned well posterior to the plate (Larson 2008).

Fig. 8. Tyrannosaurine dinosaur Daspletosaurus sp. (CMC VP15826) from the Upper Cretaceous (Late Campanian) Coal Ridge Member of the Judith River Formation, Fergus County, Montana. Vomer in right lateral (A1) and right lateral (A2) aspects. Abbreviations: ap, anterior process; apl, anterior plate; dg, dorsal groove; vk, ventral keel.
Pterygoid: Both pterygoids are represented, but only the right element (Fig. 9) is nearly complete and relatively well preserved. The right pterygoid is approximately 440 mm long and is similar in essence to that of T. rex and other tyrannosaurids (Larson 2008). The element is mediolaterally thin for its entire length. Posteriorly it possesses a dorsally projecting quadrate process that overlaps the anterior part of the medial surface of the pterygoid flange of the quadrate. A prominent anterodorsally directed ramus of the pterygoid contacts the mediodorsal surface of the palatine, and together these elements enclose a rounded fenestra (Figs. 9 and 10) that is roughly ~38 mm tall and ~30 mm long.
Ectopterygoid: The left ectopterygoid (Fig. 10A) is mostly complete, but much of its surface is badly crushed. The anterior or jugal ramus curls anteroventrally from the body of the element and bears a dorsoventrally broad, flattened surface for contacting the jugal. The ventral ramus curls posteroventrally from the body and has a single anteroposteriorly elongate (~40 mm) pneumatic excavation on its medial surface as in D. horneri, whereas two foramina are present here in the holotype of D. torosus (Carr et al. 2017). There is a ~10 mm wide accessory pneumatic foramen in the body of the ectopterygoid between the jugal and ventral processes.
Palatine: The main body of the right palatine (Fig. 10B) is largely intact, but most of the vomeropterygoid process as well as the distal ends of the jugal process, medial process, and maxillary ramus are missing. As preserved, the bone has an anteroposterior length along its ventral margin of approximately 245 mm; its maximum dorsoventral height is just over 183 mm. The posterior pneumatic recess is located behind the anterior margin of the vomeropterygoid neck, a synapomorphy of the genus Daspletosaurus (Carr et al. 2017). Posteromedially, there is a process for contacting the lateral surface of the anteroventral part of the pterygoid which, together with the posterior margin of the dorsal ramus of the palatine, forms a fenestra between these elements (Figs. 9 and 10).
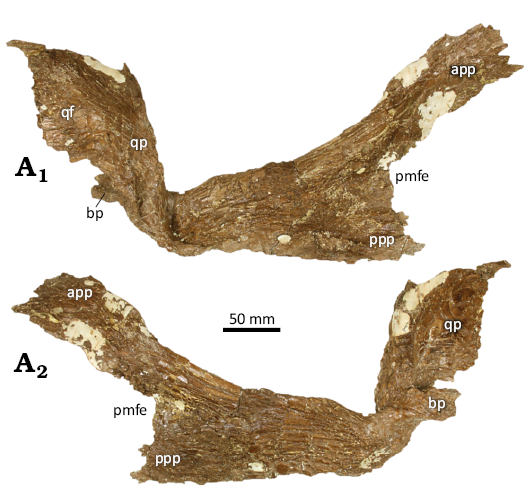
Fig. 9. Tyrannosaurine dinosaur Daspletosaurus sp. (CMC VP15826) from the Upper Cretaceous (Late Campanian) Coal Ridge Member of the Judith River Formation, Fergus County, Montana. Right pterygoid in lateral (A1) and medial (A2) aspects. Abbreviations: app, anterior palatine process; bp, basipterygoid process; pmfe, posterior margin of palatine-pterygoid fenestra; ppp, posterior palatine process; qp, quadrate process.

Fig. 10. Tyrannosaurine dinosaur Daspletosaurus sp. (CMC VP15826) from the Upper Cretaceous (Late Campanian) Coal Ridge Member of the Judith River Formation, Fergus County, Montana. A. Left ectopterygoid in ventral (A1), dorsal (A2), and medial (A3) views. B. Right palatine in lateral (B1) and medial (B2) views. Abbreviations: amfe, anterior margin of palatine-pterygoid fenestra; apr, accessory pneumatic recess; jr, jugal ramus; lf, lateral flange; mf, maxillary facet; mr, maxillary ramus; pf, pterygoid facet; pp, pterygoid process; ppr, posterior pneumatic recess; pr, pneumatic recess; vpn, vomeropterygoid neck; vr, ventral ramus.
As the holotype of D. wilsoni does not include a palatine, CMC VP15826 is here compared only to D. horneri and D. torosus. The palatine agrees closely with the holotype of D. torosus. Features shared with the holotype of D. torosus but not D. horneri include closely spaced pneumatic recesses, an internal partition between the pneumatic recesses, a posterior pneumatic recess that is as tall as the maxillary ramus, and the presence of a medial pneumatic opening (Carr et al. 2017). The vomeropterygoid neck of the palatine is oriented anterodorsally as in D. torosus (CMN 8506) rather than nearly vertically as in the holotype of D. horneri (Carr et al. 2017). The posterior pneumatic recess is rounded posteriorly and narrows anterodorsally as in D. torosus, whereas the recess in the paratype of D. horneri is not as tapered and has a more vertically oriented anterior margin (Carr et al. 2017: fig 2F).
Discussion
Review of specimens previously assigned to Daspletosaurus torosus and Daspletosaurus wilsoni.—Russell (1970) referred several tyrannosaurid specimens from Alberta to D. torosus, but a comprehensive review of these referrals has not been published. The holotype of D. torosus (CMN 8506) was collected from the Brachylophosaurus–Coronosaurus Assemblage Zone of the Oldman Formation in Dinosaur Provincial Park, Alberta, Canada (Eberth et al. 2023). Based on a review of the literature as it stood in 2017, Fowler (2017) placed Coronosaurus brinkmani at ~77.1 Ma, and Warshaw and Fowler (2022) placed D. torosus specifically at ~77 Ma. The upper age limit of this assemblage zone has been recently placed at 76.80+ Ma (Eberth et al. 2023). Postcranial elements of CMN 350 have been discussed in the literature, but diagnostic cranial elements (postorbital and lacrimal) are associated with the specimen contra Russell (1970) and Paulina-Carabajal et al. (2021). CMN 350 bears a massive cornual process of the postorbital and a prominent, triangular lacrimal cornual process, allowing for a confident referral to Daspletosaurus. CMN 11594 (includes the articulated posterior 1/2 of a skull and dentaries) from the Oldman Formation of southern Alberta possesses established synapomorphies of Daspletosaurus (massive cornual process of postorbital that approaches laterotemporal fenestra; tall, triangular lacrimal cornual process; pronounced interlocking ridges at mandibular symphysis) supporting Russell’s (1970) original referral to that genus. AMNH 5346, a maxilla from Dinosaur Provincial Park, Alberta, was referred to D. torosus by Russell (1970) and later Carr (1999) but has not been figured or redescribed. These specimens (CMN 350, CMN 11594, AMNH 5346) warrant further anatomical description and, if possible, stratigraphic placement.
At least three albertosaurine specimens have been misidentified as Daspletosaurus. NHMUK PV R4863 (formerly BMNH R4863), an articulated tyrannosaurid rostrum (dentaries, left maxilla, premaxillae, nasals), was referred to D. torosus by Russell (1970). NHMUK PV R4863 was regarded as a specimen of Gorgosaurus sp. at the time of its acquisition by the NHMUK from William E. Cutler in the 1920s. More recently, this specimen has been referred to Daspletosaurus sp. by Currie (2003). Study of this specimen by the senior author (EW-C), however, failed to identify the synapomorphies of Daspletosaurus. Specifically, the specimen lacks a coarse subcutaneous surface of the maxilla (Carr et al. 2017), and also lacks a large maxillary fenestra. Furthermore, the maxillary fenestra is well separated from the anterior margin of the antorbital fossa as in albertosaurines. We do not, therefore, regard NHMUK PV R4863 as a specimen of Daspletosaurus. Its occurrence in the “Belly River Series” rather than the Edmonton Group (Horseshoe Canyon Formation) suggests that it likely pertains to the genus Gorgosaurus, as it is the only albertosaurine known from the Belly River Group (Dinosaur Park Formation). The referral of the partial skeleton CMN 11315 from the Horseshoe Canyon Formation of Alberta to cf. Daspletosaurus by Russell (1970) indicated that this genus continued into the Maastrichtian, but a recent study reassigned this specimen to Albertosaurus sarcophagus (Mallon et al. 2019), making specimens of the late Campanian D. horneri (~75.1–74.4 Ma) the youngest confirmed examples of Daspletosaurus (Carr et al. 2017). Currie (2003) referred an ontogenetically immature (“large juvenile-subadult”; Voris et al. 2019) skull (TMP 1994.143.0001) to Daspletosaurus sp., and this diagnosis has been tentatively accepted (e.g., Hone and Tanke 2015; Carr et al. 2017). However, an isolated postorbital (TMP.2013.18.11) from a juvenile tyrannosaurid (<50% adult size) from the Dinosaur Park Formation more closely resembles the postorbital of adult Daspletosaurus than does the ontogenetically older TMP.1994.143.1 (Voris et al. 2019). This, in addition to phylogenetic data, lead Voris et al. (2019) to reassign TMP.1994.143.0001 to Gorgosaurus, though this has been questioned (Paulina-Carabajal et al. 2021).
The partial skull and skeleton of a tyrannosaurid (FMNH PR308) from the Centrosaurus apertus Zone (76.5–75.8 Ma, Eberth et al. 2023) of the Dinosaur Park Formation was assigned to Albertosaurus (= Gorgosaurus) libratus by Russell (1970), but was subsequently reassigned to Daspletosaurus torosus by Carr (1999). Despite being heavily reconstructed, FMNH PR308 can be referred to Daspletosaurus by, among other features, the presence of a massive cornual process of the postorbital that approaches the laterotemporal fenestra. Due to its stratigraphic overlap with D. wilsoni, Warshaw et al. (2024) inferred that this specimen may possibly be assignable to D. wilsoni. Currie (2003) referred the undescribed “skull and skeleton” TMP 92.36.1220 from Dinosaur Provincial Park and the undescribed TMP 98.48.1 to Daspletosaurus sp.
Pending further study, the only definitive specimen of Daspletosaurus torosus with reliable stratigraphic provenance is the holotype (CMN 8506). Carr et al. (2017) stated that D. torosus is “restricted to the lower two-thirds of the Dinosaur Park Formation (~76.7–75.2 Ma).” However, Carr et al. (2017) did not indicate which specimens of D. torosus represent the upper limit of this stratigraphic range. CMN 8506 was recovered from the Oldman Formation, not the overlying Dinosaur Park Formation (Paulina-Carabajal et al. 2021). An isolated tyrannosaurid frontal collected by Charles Sternberg in 1921 within the present boundaries of Dinosaur Provincial Park was referred to D. torosus by Yun (2020), but its stratigraphic position is unknown, and its diagnostic value has been questioned (Paulina-Carabajal et al. 2021). Voris et al. (2019) regard all documented Daspletosaurus specimens from the Oldman and Dinosaur Park formations (including the complete skull TMP 2001.36.1 and the partial skull TMP 1985.62) as provisional specimens of D. torosus. However, some of these specimens, including TMP 2001.36.1, and others from the Dinosaur Park Formation and equivalent beds in the Oldman Formation have yet to be thoroughly described, and have been hypothesized to represent a new, undescribed species (Paulina-Carabajal et al. 2021; Scherer and Voiculescu-Holvad 2024). Most recently, Warshaw et al. (2024) referred TMP 2001.36.1 to D. wilsoni, but further descriptions of TMP 2001.36.1 and TMP 1985.62 are ongoing by other workers (Paulina-Carabajal et al. 2021; Colton Coppock, personal communication 2023).
Several specimens have recently been assigned to D. wilsoni by Warshaw et al. (2024) based on the presence of a dorsal quadrate process of the quadratojugal that is broadly exposed in lateral view, a narrow orbit, and an inflated “rostrodorsal ala” of the lacrimal. Among these are at least three specimens from a single, multi-taxon bonebed in the Two Medicine Formation of Montana (Currie et al. 2005). Although the stratigraphic position and numeric age of this material is not well resolved, these specimens can confidently be assigned to Daspletosaurus by the presence of a massive postorbital cornual process that approaches the laterotemporal fenestra among other features. These specimens were hypothesized to belong to the same species as MOR 590 prior to the description of D. horneri (Currie et al. 2005). This collection shares several features with both D. torosus and D. wilsoni to the exclusion of D. horneri, including the presence of a long, anteriorly tapered and dorsally offset maxillary fenestra as well as a tall cornual process of the lacrimal. Further study of the morphology and stratigraphic position of this collection is clearly warranted. TMP 2003.10.3, a largely complete skull of an ontogenetically mature specimen of Daspletosaurus from the Upper Oldman Formation of the Milk River region of southern Alberta, was recently assigned to D. wilsoni (Warshaw et al. 2024). This specimen reportedly occurs in beds correlative to the Corythosaurus–Centrosaurus Assemblage of the lower Dinosaur Park Formation and has thus been estimated to be ~76.5–75.6 Ma by Warshaw et al. (2024).
Taxonomic identity of CMC VP15826.—CMC VP15826 has a combination of features that has not been previously reported in Daspletosaurus. Although it lacks most diagnostic characters of D. horneri, CMC VP15826 shares some features with that species (distal end of dorsal jugal facet of quadratojugal not visible laterally, one foramen in the ventral ramus of the ectopterygoid, short cervical epipophyses; Carr et al. 2017) that are not present in the holotype of D. torosus. This unique suite of features could be interpreted as evidence that CMC VP15826 represents a new species with a transitional morphology between the chronologically older D. torosus (or D. wilsoni) and the younger D. horneri. Alternatively, such features may be intraspecifically variable within these taxa and therefore of no taxonomic significance, as the limited sample size of previously described specimens of D. torosus and D. wilsoni cannot be considered representative of the potential range of morphological disparity within these taxa.
One of the three characters purported to distinguish D. wilsoni from D. torosus is the presence of a dorsal quadrate “contact” of the quadratojugal that is broadly visible in lateral view (Warshaw et al. 2024). Because the dorsal quadrate contact is a surface that is necessarily only visible in medial view, we will hereon refer to it as the “dorsal quadrate process” instead of the “dorsal quadrate contact”. Based on a photograph of the specimen (Maltese 2009: fig. 3), Warshaw et al. (2024) argue that the dorsal quadrate process of the quadratojugal in CMC VP15826 is broadly visible in lateral view as in D. wilsoni because there is a “cleft” between the dorsal quadrate process and the body of the quadratojugal that “separates the two structures”. During the process of preparation and restoration, this “cleft” was interpreted as a break and was therefore filled in with white putty (Fig. 7A, B). It is unclear to us if this notch is homologous to the cleft present in the quadratojugal of BDM 107, or if it represents a break, as this part of the bone is extremely thin and fragile. The dorsal margin of the quadratojugal often forms an irregular margin in tyrannosaurids, and such a cleft may be present in taxa that do not have a laterally exposed dorsal quadrate process. In a specimen of Tyrannosaurus rex (“Stan”, now formerly BHI 3033 in the collection of the natural history museum currently under construction in Abu Dhabi, UAE) for example, a taxon coded as having a medially directed dorsal quadrate process (Warshaw and Fowler 2022; Scherer and Voiculescu-Holvad 2024; Warshaw et al. 2024), there is a similar cleft on the left quadratojugal, but no such cleft on the right quadratojugal (Larson 2008). Such a cleft, therefore, may be present in specimens that do not have a laterally exposed dorsal quadrate process. Contra Warshaw et al. (2024), the dorsal quadrate process in CMC VP15826 is oriented sharply medially, giving the dorsal ramus of the quadratojugal an anteroposteriorly narrow lateral profile as in D. torosus (CMN 8506) and a specimen of D. horneri (MOR 1130) as opposed to the broad profile of D. wilsoni (BDM 107) and the holotype of D. horneri (MOR 590). Warshaw et al. (2024) observe that the dorsal quadrate process of the quadratojugal in FMNH PR308 is not broadly visible laterally. Two specimens (CMC VP15826 and FMNH PR308) that occur within the proposed temporal range of D. wilsoni therefore lack one of the purportedly diagnostic characters of that species. The presence of this character state in specimens of this age indicates either that this character is intraspecifically variable in Daspletosaurus or that there were two sympatric species of Daspletosaurus in the late Campanian. We favor the former explanation over the latter due to the small sample size at hand and because two congeneric tyrannosaurd species have never been shown to conclusively overlap temporospatially. If CMC VP15826 and FMNH PR308 are interpreted as being taxonomically distinct from D. wilsoni, however, then the sympatry of these taxa would support cladogenesis within Daspletosaurus. Tyrannosaurus rex has been described as having a medially directed dorsal quadrate process of the quadratojugal (Warshaw and Fowler 2022). It does appear that in some specimens of T. rex (AMNH 5027, MOR 008, MOR 555, FMNH PR2081, LACM 23844, UWBM) the dorsal quadrate process is directed more medially than it is in D. wilsoni (BDM 107) and the holotype of D. horneri (MOR 590). However, in a large adult specimen of T. rex (RSM P2523.8; Persons et al. 2020: fig. 15), this process appears to be laterally exposed and entirely visible in lateral view as in BDM 107 and MOR 590, thus indicating that this feature may be intraspecifically variable in T. rex. Given its potential for variability within Tyrannosaurus, a genus represented by many more specimens than Daspletosaurus, we are skeptical of the taxonomic significance of this feature for diagnosing species of Daspletosaurus. This character is known to be variable in D. horneri, and because the only definitive specimen of D. torosus is the holotype (CMN 8506), its potential variability cannot be assessed in that taxon. If this character is variable within either D. torosus or D. wilsoni, CMC VP15826 cannot at present be confidently assigned to or excluded from either D. wilsoni or D. torosus. We interpret the apparent variability of this character in Daspletosaurus specimens from the proposed temporal range of D. wilsoni (~76.5–75.8 Ma) as intraspecific morphological disparity rather than contemporary taxonomic diversity, thereby reducing the number of features that may distinguish D. wilsoni from D. torosus to two (narrow orbit and “inflated rostrodorsal ala” of lacrimal). Study of additional Daspletosaurus specimens with stratigraphic controls are necessary to determine if these remaining two characters are sufficiently robust to support the validity of D. wilsoni as distinct from D. torosus. However, cladogenesis remains a viable model due to the rarity of stratigraphically well-constrained specimens.
Conclusions
CMC VP15826 is a large, ontogenetically mature specimen of Daspletosaurus from the upper Campanian Coal Ridge Member of the Judith River Formation of Central Montana. This specimen is necessarily less than ~76.3 Ma, but potentially as young as ~75.8 Ma, with a maximum model age of 76.12 +0.14/-0.47 Ma and a minimum model age of 75.81 +0.33/-0.37 Ma. CMC VP15826 can be excluded from D. horneri, as it is chronologically older (by ~0.6–1.1 Myr) than and lacks most diagnostic features of that taxon. CMC VP15826 shares numerous features with both D. wilsoni and D. torosus, but lacks one of only three characters proposed by Warshaw et al. (2024) to distinguish D. wilsoni from D. torosus (laterally exposed dorsal quadrate process of quadratojugal). Although the morphological disparity between the new specimen and the stratigraphically equivalent D. wilsoni could be interpreted as evidence of contemporary taxonomic diversity within Daspletosaurus, we contend that this feature is intraspecifically variable and therefore taxonomically uninformative, thereby weakening the case that D. wilsoni is distinct from D. torosus. The presence of the other two characters purported to distinguish D. wilsoni from D. torosus (narrow orbit, inflated “rostrodorsal ala” of lacrimal) cannot be evaluated in CMC VP15826, because the lacrimal and postorbital are not preserved. CMC VP15826 cannot at present be confidently assigned to or excluded from either D. torosus or D. wilsoni. At ~75.1–74.4 Ma, D. horneri remains the stratigraphically youngest species of Daspletosaurus, and may therefore have arisen from either D. torosus or D. wilsoni through anagenesis as proposed by Carr et al. (2017). Future work should be focused on deciphering the interspecific and intraspecific morphological variation in Daspletosaurus and determining the specific identities and stratigraphic ranges of available specimens.
Acknowledgments
We thank the late Glenn Peterson who owned the ranch on which CMC VP15826 was discovered. We thank an anonymous local rancher and amateur paleontologist for the discovery and reporting of the site, Michael Triebold, Jacob Jett, and Kraig Derstler of the Rocky Mountain Dinosaur Resource Center (Woodland Park, Colorado, USA) for the collection and preparation of the specimen. Cameron Schwalbach and Spencer Shroyer (both CMC) aided with photography of CMC VP15826. For on-site access to collections in their care we thank the following individuals: Carrie Levitt-Bussian, Randall B. Irmis, and Tylor Birthisel (all UMNH), Marc E.H. Jones and Paul Barrett (both NHMUK). The following individuals greatly improved the quality of this paper through their generous sharing of photographs and/or discussions on the content contained herein: Eric T. Metz and Sebastian Dalman (MOR), Jordan C. Mallon and Shyong Pen (both CMN), James I. Kirkland, Donald DeBlieux (both Utah Geological Survey), Mariangelica Groves, Ronald Meyers, and Elizabeth Balgord (all Weber State University, Ogden, USA), Matthew Lamanna (Carnegie Museum of Natural History, Pittsburgh, USA), Colton Coppock (University of Alberta, Edmonton, Canada), Brian Curtice (Fossil Crates, Tempe, USA), Jahan Ramezani (Massachusetts Institute of Technology, Cambridge, USA), Tut Tran (Bryce Canyon National Park, Utah, USA), Loren Brunken (Spyhop, Salt Lake City, USA), Thomas Carr (Carthage College, Kenosha, USA). Thomas R. Holtz (University of Maryland, College Park, USA) and James Napoli (University of New York at Stony Brook, USA) provided useful commentary and greatly improved the manuscript.
Editor: Daniel Barta.
References
Arbour, V.M. and Evans, D.C. 2017. A new ankylosaurine dinosaur from the Judith River Formation of Montana, USA based on an exceptional skeleton with soft tissue preservation. Royal Society open Science 4: 161086. Crossref
Brochu, C.A. 2003. Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull. Journal of Vertebrate Paleontology 22 (Supplement 4): 1–138. Crossref
Brusatte, S.L. and Carr, T.D. 2016. The phylogeny and evolutionary history of tyrannosauroid dinosaurs. Scientific Reports 6: 20252. Crossref
Brusatte, S.L., Norell, M.A., Carr, T.D., Erickson, G.M., Hutchinson, J.R., Balanoff, A.M., Bever, G.S., Choiniere, J.N., Makovicky, P.J., and Xing, X. 2010. Tyrannosaur paleobiology: new research on ancient exemplar organisms. Science 329: 1481–1485. Crossref
Carr, T.D. 1999. Craniofacial ontogeny in tyrannosauridae (Dinosauria, Coelurosauria). Journal of vertebrate Paleontology 19: 497–520. Crossref
Carr, T.D., Varricchio, D.J., Sedlmayr, J.C., Roberts, E.M., and Moore, J.R. 2017. A new tyrannosaur with evidence of anagenesis and crocodile-like facial sensory system. Scientific Reports 7: 44942. Crossref
Carr, T.D., Williamson. T.E., Britt, B.B., and Stadtman, K. 2011. Evidence for high taxonomic and morphologic tyrannosauroid diversity in the Late Cretaceous (Late Campanian) of the American Southwest and a new short-skulled tyrannosaurid from the Kaiparowits Formation of Utah. Naturwissenschaften 98: 241–246. Crossref
Cope, E.D. 1876. Descriptions of some vertebrate remains from the Fort Union Beds of Montana. Proceedings of the Academy of Natural Sciences of Philadelphia 28: 248–261.
Cullen, T.M., Larson, D.W., Witton, M.P., Scott, D., Maho, T., Brink, K.S., Evans, D.C., and Reisz, R. 2023. Theropod dinosaur facial reconstruction and the importance of soft tissues in paleobiology. Science 379: 1348–1352. Crossref
Currie, P.J. 2003. Cranial anatomy of tyrannosaurid dinosaurs from the Late Cretaceous of Alberta, Canada. Acta Palaeontologia Polonica 48: 191–226.
Currie, P.J., Trexler, D., Koppelhus, E.B., Wicks, K., and Murphy, N. 2005. An unusual multi-individual bonebed in the Two Medicine Formation (Late Campanian) of Montana (USA). In: K. Carpenter (ed.), The Carnivorous Dinosaurs, 313–324. Indiana University Press, Bloomington.
Cuthbertson, R.S. and Holmes, R.B. 2010. The first complete description of the holotype of Brachylophosaurus canadensis Sternberg, 1953 (Dinosauria: Hadrosauridae) with comments on intraspecific variation. Zoological Journal of the Linnean Society 159: 373–397. Crossref
Dalman, S.G. and Lucas, S.G. 2015. New data on the tyrannosaurid theropods from the middle Campanian lower Judith River Formation of north-central Montana. New Mexico Museum of Natural History and Science Bulletin 67: 19–31.
Dodson, P. 1986. Avaceratops lammersi: a new ceratopsid from the Judith River Formation of Montana. Proceedings of the Academy of Natural Sciences, Philadelphia 138: 305–31.
Eberth, D.E., Evans, D.C., Ramezani, J., Kamo, S.L., Brown, C.M., Currie, P.J., and Braman, D.R. 2023. Calibrating geologic strata, dinosaurs, and other fossils at Dinosaur Provincial Park (Alberta, Canada) using a new CA-ID-TIMS U-Pb geochronology. Canadian Journal of Earth Sciences 60: 1627–1646. Crossref
Farlow, J.O., Coroian, D., Currie, P.J., Foster, J.R., Mallon, J.C., and Therrien, F. 2023. “Dragons” on the landscape: modeling the abundance of large carnivorous dinosaurs of the Upper Jurassic Morrison Formation (USA) and the Upper Cretaceous Dinosaur Park Formation (Canada). The Anatomical Record 306: 1669–1696. Crossref
Fowler, D.W. 2017. Revised geochronology, correlation, and dinosaur stratigraphic ranges of the Santonian–Maastrichtian (Late Cretaceous) formations of the Western Interior of North America. PLOS ONE 12 (11): e0188426. Crossref
Freedman-Fowler, E.A. and Horner, J.R. 2015. A new brachylophosaurin hadrosaur (Dinosauria: Ornithischia) with an intermediate nasal crest from the Campanian Judith River Formation of northcentral Montana. PLOS ONE 10 (11): e0141304. Crossref
Gignac, P.M. and Erickson, G.M. 2017. The biomechanics behind extreme osteophagy in Tyrannosaurus rex. Scientific Reports 7 (12): 1–10. Crossref
Hone, D. and Tanke, D. 2015. Pre- and postmortem tyrannosaurid bite marks on the remains of Daspletosaurus (Tyrannosaurinae: Theropoda) from Dinosaur Provincial Park, Alberta, Canada. PeerJ 3: e885. Crossref
Horner, J.R., Varricchio, D.J., and Goodwin, M.B. 1992. Marine transgressions and the evolution of Cretaceous dinosaurs. Nature 358: 59–61. Crossref
Huene, F.V. 1914. Das natürliche System der Saurischia. Zentralblatt für Mineralogie, Geologie und Paläontologie B 1914: 154–158.
Ishikawa, H., Tsuihiji, T., and Manabe, M. 2023. Furcatoceratops elucidans, a new centrosaurine (Ornithischia: Ceratopsidae) from the upper Campanian Judith River Formation, Montana, USA. Cretaceous Research 151: 105660. Crossref
Lambe, L.M. 1917. The Cretaceous theropodous dinosaur Gorgosaurus. Geological Survey of Canada Memoir 100: 1–84. Crossref
Larson, P. 2008. Atlas of the skull bones of Tyrannosaurus rex. In: P. Larson and K. Carpenter (eds.), Tyrannosaurus rex, the Tyrant King, 233–243. Indiana University Press, Bloomington.
Loewen, M.A., Irmis, R.B., Sertich, J.J., Currie, P.J., and Sampson, S.D. 2013. Tyrant dinosaur evolution tracks the rise and fall of Late Cretaceous oceans. PLOS ONE 8 (11): e79420. Crossref
Loewen, M.A, Sertich J.J.W., Sampson S., O’Connor, J.K., Carpenter, S., Sisson, B., Øhlenschlæger, A., Farke, A.A., Makovicky, P.J., Longrich, N., and Evans, D.C. 2024. Lokiceratops rangiformis gen. et sp. nov. (Ceratopsidae: Centrosaurinae) from the Campanian Judith River Formation of Montana reveals rapid regional radiations and extreme endemism within centrosaurine dinosaurs. PeerJ 12:e17224. Crossref
Longrich, N.R. 2013. Judiceratops tigris, a new horned dinosaur from the Middle Campanian Judith River Formation of Montana. Bulletin of the Peabody Museum of Natural History 54: 51–65. Crossref
Mallon, J.C., Bura, J.R., Schumann, D., and Currie, P.J. 2019. A problematic tyrannosaurid (Dinosauria: Theropoda) skeleton and its implications for tyrannosaurid diversity in the Horseshoe Canyon Formation (Upper Cretaceous) of Alberta. The Anatomical Record 303: 673–690. Crossref
Mallon, J.C., Ott, C.J., Larson, P.L., Iuliano, E.M., and Evans, D.C. 2016. Spiclypeus shipporum gen. et sp. nov., a boldly audacious new chasmosaurine ceratopsid (Dinosauria: Ornithischia) from the Judith River Formation (Upper Cretaceous: Campanian) of Montana, USA. PLOS ONE 11 (5): e0154218. Crossref
Maltese, A.E. 2009. Difficult excavation and preparation of a large Daspletosaurus specimen. In: M.A. Brown, J.F. Kane, and W.G. Parker (eds.), Methods in Fossil Preparation: Proceedings of the First Annual Fossil Preparation and Collections Symposium, 63–68.
Marsh, O.C. 1881. Principal characters of American Jurassic dinosaurs, Part V. American Journal of Science 21: 417–423. Crossref
Marsh, O.C. 1888. A new family of horned Dinosauria, from the Cretaceous. The American Journal of Science, Series 3, 36: 477–478. Crossref
Molnar, R.E. 1991. The cranial morpholgy of Tyrannosaurus rex. Palaeontographica. Abteilung A, Paläozoologie, Stratigraphie 217: 137–176.
Osborn, H.F. 1906. Tyrannosaurus, Upper Cretaceous carnivorous dinosaur. Bulletin of the American Museum of Natural History 22: 281–296.
Owen, R. 1842. Report on British fossil reptiles. Part II. Report of the British Association for the Advancement of Science 11: 60–204.
Paulina-Carabajal, A., Currie, P.J., Dudgeon, T., Larsson, H.C.E., and Miyashita, T. 2021. Two braincases of Daspletosaurus (Theropoda: Tyrannosauridae): anatomy and comparison. Canadian Journal of Earth Sciences 58: 885–910. Crossref
Persons, W.S., Currie, P.J., and Erickson, G.M. 2020. An older and exceptionally large adult specimen of Tyrannosaurus rex. The Anatomical Record 303 (4): 656–672. Crossref
Ramezani, J., Beveridge, T.L., Rogers, R.R., Eberth, D.A., and Roberts, E.M. 2022. Calibrating the zenith of dinosaur diversity in the Campanian of the Western Interior Basin by CA-ID-TIMS U-Pb geochronology. Scientific Reports 12: 16026. Crossref
Rogers, R.R., Eberth, D.A., and Ramezani, J. 2024. The “Judith River-Belly River problem” revisited (Montana-Alberta-Saskatchewan): New perspectives on the correlation of Campanian dinosaur bearing strata based on a revised stratigraphic model updated with CA-ID-TIMS U-Pb geochronology. The Geological Society of America Bulletin 136: 1221–1237. Crossref
Rogers, R.R., Horner, J.R., Ramezani, J., Roberts, E.M., and Varricchio, D.J. 2025. Updating the Upper Cretaceous (Campanian) Two Medicine Formation of Montana: Lithostratigraphic revisions, new CA-ID-TIMS U-Pb ages, and a calibrated framework for dinosaur occurrences. The Geological Society of America Bulletin 137: 315–340. Crossref
Rogers, R.R., Kidwell, S.M., Deino, A.L., Mitchell, J.P., Nelson, K., and Thole, J.T. 2016. Age, correlation, and lithostratigraphic revision of the Upper Cretaceous (Campanian) Judith River Formation in its type area (north-central Montana), with a comparison of low- and high-accommodation alluvial records. The Journal of Geology 124: 99–135. Crossref
Russell, D.A. 1970. Tyrannosaurs from the Late Cretaceous of western Canada. Canadian Museum of Nature, Publications in Paleontology 1: 1–34.
Ryan, M.J., Evans, D.C., Currie, P.J., and Loewen, M.A. 2014. A new chasmosaurine from northern Laramidia expands frill disparity in ceratopsid dinosaurs. Naturwissenschaften 101: 505–512. Crossref
Ryan, M.J., Russell, A.P., and Hartman, S.C. 2010. A new chasmosaurine ceratopsid from the Judith River Formation, Montana. In: M.J. Ryan, B.J. Chinnery-Allgeier, and D.A. Eberth (eds.), New Perspectives on Horned Dinosaurs. The Royal Tyrrell Museum Ceratopsian Symposium, 181–188. Indiana University Press, Bloomington.
Scherer, C.R. and Voiculescu-Holvad, C. 2024. Reanalysis of a dataset refutes claims of anagenesis within Tyrannosaurus-line tyrannosaurines (Theropoda, Tyrannosauridae). Cretaceous Research 155: 105780. Crossref
Takasaki C.R., Chiba, K., Fiorillo, A.R., Brink, K.S., Evans, D.C., Fanti, F., Saneyoshi, M., Maltese, A., and Ishigaki, S. 2022. Description of the first definitive Corythosaurus (Dinosauria, Hadrosauridae) specimens from the Judith River Formation in Montana, USA and their paleobiogeographical significance. The Anatomical Record 306: 1918–1938. Crossref
Thomson, T.J., Irmis, R.B., and Loewen, M.A. 2013. First occurrence of a tyrannosaurid dinosaur from the Mesaverde Group (Neslen Formation) of Utah: implications for upper Campanian Laramidian biogeography. Cretaceous Research 43: 70–79. Crossref
Voris, J.T., Therrien, F., Zelenitsky, D.K., and Brown, C.M. 2020. A new tyrannosaurine (Theropoda: Tyrannosauridae) from the Campanian Foremost Formation of Alberta, Canada, provides insight into the evolution and biogeography of tyrannosaurids. Cretaceous Research 110: 104388. Crossref
Voris, J.T., Zelenitsky, D.K., Therrien, F., and Currie, P.J. 2019. Reassessment of a juvenile Daspletosaurus from the Late Cretaceous of Alberta, Canada with implications for the identification of immature tyrannosaurids. Scientific Reports 9 (1): 1–10. Crossref
Warshaw, E.A. and Fowler, D.W. 2022. A transitional species of Daspletosaurus Russell, 1970 from the Judith River Formation of eastern Montana. PeerJ 10:e14461. Crossref
Warshaw, E.A., Guevara, D.B., and Fowler, D.W. 2024. Anagenesis and the tyrant pedigree: A response to “Re-analysis of a dataset refutes claims of anagenesis within Tyrannosaurus-line tyrannosaurines (Theropoda, Tyrannosauridae)”. Cretaceous Research 163: 105957. Crossref
Wilson, J.P. and Fowler, D.W. 2020. The easternmost occurrence of Saurornitholestes from the Judith River Formation, Montana, indicates broad biogeographic distribution of Saurornitholestes in the Western Interior of North America. Historical Biology 12: 3302–3306. Crossref
Witmer, L.M. and Ridgely, R.C. 2009. New insights into the brain, braincase, and ear region of tyrannosaurs (Dinosauria, Theropoda), with implications for sensory organization and behavior. The Anatomical Record 292: 1266–1296. Crossref
Yun, C.-G. 2020. A subadult frontal of Daspletosaurus torosus (Theropoda: Tyrannosauridae) from the Late Cretaceous of Alberta, Canada with implications for tyrannosaurid ontogeny and taxonomy. PalArch’s Journal of Vertebrate Palaeontology 17: 1–13.
Zietlow, A.R. 2020. Craniofacial ontogeny in Tylosaurinae. PeerJ 8: e10145. Crossref
Acta Palaeontol. Pol. 70 (1): 159–174, 2025
https://doi.org/10.4202/app.01143.2024