

A new ichthyosaur from the Lower Jurassic of Portugal (Iberian Peninsula)
JOÃO PRATAS E SOUSA, ISABEL MORAIS ROLDÃO, MARÍA RÍOS, and EDUARDO PUÉRTOLAS-PASCUAL
Pratas e Sousa, J., Morais Roldão, I., Ríos, M., and Puértolas-Pascual, E. 2025. A new ichthyosaur from the Lower Jurassic of Portugal (Iberian Peninsula). Acta Palaeontologica Polonica 70 (1): 179–192.
Ichthyosaurs were a group of marine Mesozoic reptiles. Their presence in the Portuguese fossil record is scarce and ranges from the Sinemurian to the Toarcian. In this work we describe a new specimen—a nearly complete skull discovered in the Sinemurian of Praia de Água de Madeiros, São Pedro de Moel, Municipality of Marinha Grande (Portugal). This specimen represents the most complete and best preserved cranial remains from the Iberian Peninsula published to date. Phylogenetic analyses performed identified this specimen as a new genus and species, Gadusarus aqualigneus, the first to be identified in the Iberian Peninsula. The species is phylogenetically recovered as a baracromian ichthyosaur, part of the Neoichthyosauria and Parvipelvia clades.
Key words: Parvipelvia, Ichthyosauria, Gadusaurus aqualigneus, phylogeny, Sinemurian, São Pedro de Moel.
João Pratas e Sousa [jrp.sousa@campus.fct.unl.pt; ORCID: https://orcid.org/0000-0002-3305-6444 ] and María Ríos [maria.rios.iba@fct.unl.pt; ORCID: https://orcid.org/0000-0001-7242-1259 ], GeoBioTec, FCT-NOVA Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa, 2829-516 Caparica, Portugal; GEAL Museu da Lourinhã, Rua João Luís de Moura, Lourinhã, Portugal.
Isabel Morais Roldão [iroldao@esfrl.edu.pt; ORCID: https://orcid.org/0009-0006-8630-2756 ], ESFRL Escola Secundária Francisco Rodrigues Lobo, Rua Afonso Lopes Vieira, Leiria, Portugal], GEAL Museu da Lourinhã, Rua João Luís de Moura, Lourinhã, Portugal.
Eduardo Puértolas-Pascual [eduardo.puertolas@gmail.com; ORCID: https://orcid.org/0000-0003-0759-7105 ], Grupo Aragosaurus-IUCA, Departamento de Ciencias de la Tierra, Facultad de Ciencias, Universidad de Zaragoza, Calle Pedro Cerbuna 12, 50009 Zaragoza, Spain.
Received 10 August 2024, accepted 19 February 2025, published online 31 March 2025.
Copyright © 2025 J. Pratas e Sousa et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Ichthyosaurs were a group of Mesozoic reptiles best known for their re-adaptation to a marine environment. They ranged in age from the Early Triassic to the Late Cretaceous, but the ichthyosaur fauna of the Early Jurassic is the most frequently preserved in the fossil record of Europe (Motani 1999; Sander 2000). Although the remains of ichthyosaurs are quite abundant in marine deposits in some regions of Europe, their record is very scarce and fragmentary in southern areas such as the Iberian Peninsula (Portugal and Spain).
Ichthyosauria was first documented in Portugal in Lower Jurassic formations during stratigraphic fieldwork carried out by the Serviços Geológicos de Portugal. Paul Choffat, in 1885–1886, mentions the presence of flattened vertebrae in the Pliensbachian of Pintanheira, Alcobaça. Later in 1898, Henri Sauvage identified a rostrum fragment found in the Aalenian of Alhadas, Municipality of Figueira da Foz, as Ichthyosaurus sp. (Sauvage 1898). In 1952, Zbyszewski and Moitinho de Almeida (1952) reported maxillary fragments, an anterior right limb, and vertebrae collected by Joaquim Pintassilgo and Ernest Fleury (1878–1958) from the Sinemurian of Praia de Nossa Senhora da Victória, Alcobaça. These were identified as belonging to the species Ichthyosaurus intermedius Conybeare, 1822. Other remains, such as maxillary and limb fragments and vertebrae from the Pliensbachian of Pintanheira, were assigned to Stenopterygius uniter Huene, 1931, and maxillary fragments from Alhadas (Aalenian) and Murtede (undetermined age), Municipality of Cantanhede, and vertebrae from Cadima (undetermined age), Municipality of Cantanhede, and Casal Comba (Casal do Combo), Municipality of Mealhada, were identified as Ichthyosaurus indet. (Zbyszewski and Moitinho de Almeida 1952). In 1958, Ferreira documented a jaw fragment and vertebra from the Pliensbachian of Praia da Nossa Senhora da Victória and vertebrae from the Toarcian of Condeixa, Municipality of Condeixa-a-Nova, and the Aalenian of Tomar, Municipality of Torres Novas, as S. uniter. Vertebrae from the Sinemurian of Praia da Nossa Senhora da Victória, Águas Madeiras (Águas de Madeiros), Municipality of Marinha Grande, Casal Comba and the Pliensbachian of Alvaiázere were identified as Ichthyosaurus intermedius (Ferreira 1958).
All this documented material is fragmentary, and before more recent discoveries, the limb reported by Zbyszewski and Moitinho de Almeida, IST-MDT 85, was the most complete specimen on the Portuguese record, having been identified as Ichthyosaurus cf. communis Beche & Conybeare, 1821 (Sousa and Mateus 2021).
In Spain, Ichthyosauria are less well documented, but similarly fragmentary, with the first mention of the group being made by Suaréz Vega in 1974, who reported vertebrae from the Pliensbachian of Santa Mera (Villaviciosa, Asturias) (Suaréz Vega 1974). However, it was not until the second half of the 1990s that fossil bone remains began to be systematically recovered from this area of Asturias (Ruiz-Omeñaca et al. 2006). In 2011, Fortuny et al. reported fragmentary elements (a durophagous tooth crown and a vertebral centrum) from the Ladinian of Collbáto, (Barcelona, Catalonia), identifying them as potentially belonging to an indeterminate ichthyosaur (Fortuny et al. 2011). In 2015, Miguel Chavez et al. reported three incomplete vertebrae, identified as Ichthyosauria indet., from the Carnian of Manzanera (Teruel, Aragón) (Knoll et al. 2004; Miguel Chavez et al. 2015). In 2018, Fernández et al. reported several specimens from the Pliensbachian of the Asturian coast, between Gijón and Ribadesella. Most of this fragmentary material classified as Ichthyosauria indet, but an incomplete skeleton was identified as Leptonectes sp. (Fernández et al. 2018). More recently, in 2021, Matamales-Andreu et al. have reported an anterior caudal vertebra from the Ladinian of Mallorca, which was identified as a potential ichthyosauriform, possibly Grippia, a genus older and more basal than the Ichthyosauria clade (Matamales-Andreu et al. 2021). Other indeterminate ichthyosauria remains have been documented from the Pliensbachian of Ólvega (Soria) (Bardet et al. 2008).
Our study focuses on the recently discovered skull, ML 2750, and identified as a new genus and species. The investigation entails a detailed analysis of the cranium in order to unveil the complexities of its morphology and clarify the distinctive adaptations that differentiate this species from other coeval ones.
Institutional abbreviations.—ML, Museu da Lourinhã, Portugal; IST-MDT, Instituto Superior Técnico, Museu Décio Thadeu, Lisbon, Portugal.
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank:urn:lsid:zoobank.org:pub:6A033F31-CCE4-4738-BD96-78A9EE394A63.
Geological setting
The studied specimen ML 2750 was collected in mainland Portugal, in the São Pedro de Moel area (Água de Madeiros). Specifically, the block containing the fossil was discovered on the sands of Água de Madeiros beach, in the stream that forms the border between the municipalities of Marinha Grande and Alcobaça. Its precise location is given by the coordinates 39°44’28.20” N, 9°2’20.92” W (Fig. 1A).
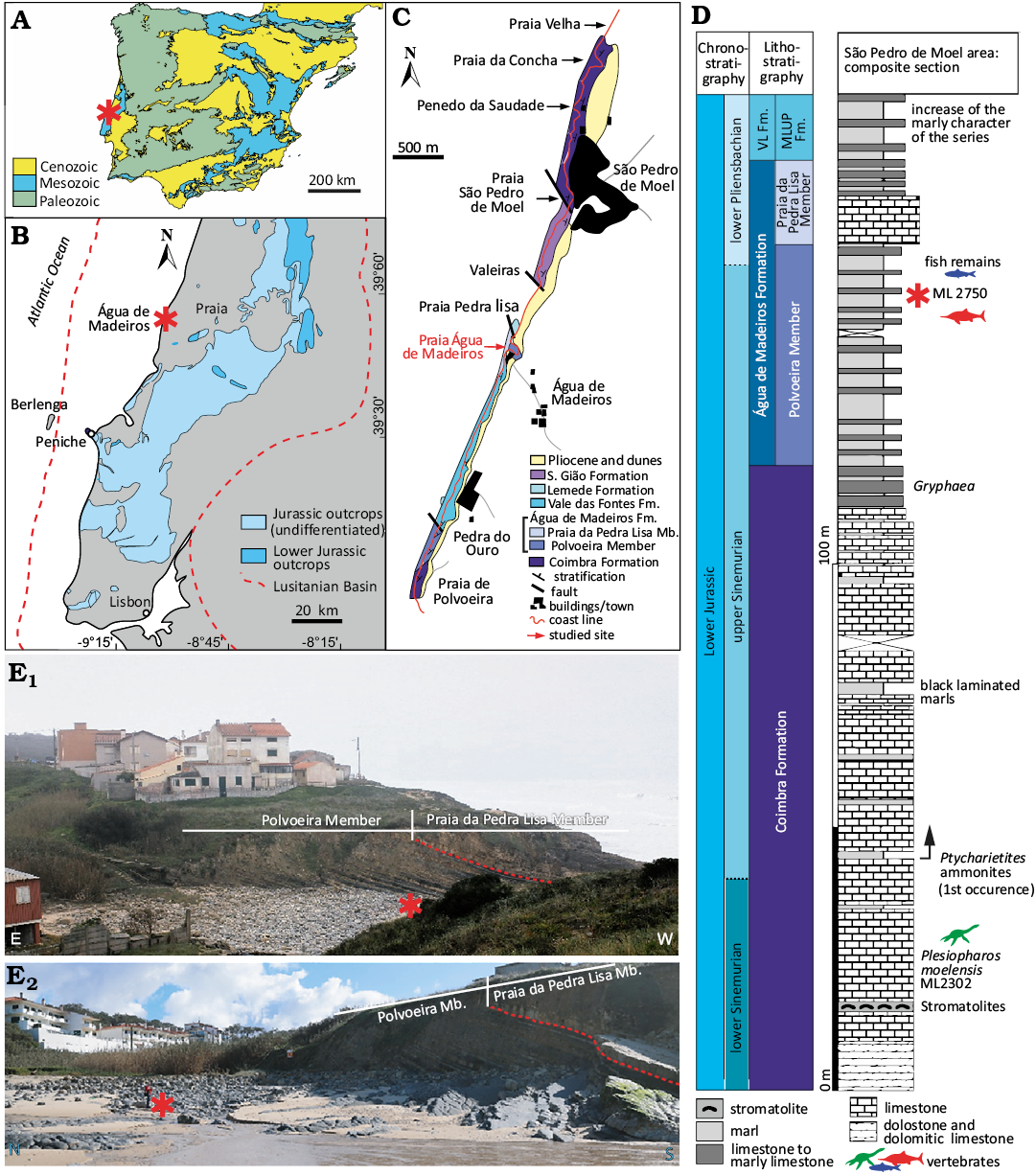
Fig. 1. Geographical and geological settings of Gadusaurus aqualigneus gen. et sp. nov. (ML 2750) locality (asterisk) from the Lower Jurassic of São Pedro de Moel (Marinha Grande, Portugal). A. Geological map of the Iberian Peninsula. B. The Jurassic outcrops in the Lusitanian Basin. C. Geological mapping of the outcropping rock units in the São Pedro de Moel region with location of the ML 2750. D. Synthetic stratigraphic column of the Lower Jurassic of São Pedro de Moel. E. Panoramic views of the outcrops (E1) and units of the Água de Madeiros Formation in Praia de Água de Madeiros (E2). Modified from Duarte et al. (2014) and Puértolas-Pascual et al. (2021). Abbreviations: Fm., Formation; Mb. Member.
This locality is situated within the confines of the Lusitanian Basin, which is located on the western Iberian passive margin of the Atlantic Ocean, and provides an excellent marine record of the Lower Jurassic. The Lusitanian Basin is a north-south elongated basin, with borders defined by the Iberian Massif to the east and by the Variscan Berlenga Horst to the west. The development of this basin occurred during the opening of the Atlantic Ocean, and it has undergone several rifting and sedimentary phases with deposits spanning from the Upper Triassic to the Upper Cretaceous (e.g., Wilson et al. 1989; Alves et al. 2002; Duarte et al. 2014).
The deposits studied in the present work were formed during the initial phase, spanning from the Triassic to the Middle Jurassic. The Lower Jurassic is primarily composed by marine carbonate deposits (Soares et al. 1993; Duarte et al. 2004, 2014). The site is situated within the Polvoeira Member of the Água de Madeiros Formation, a formation that has been dated (based on ammonite biostratigraphy) between the Oxynotum Chronozone (upper Sinemurian) and the base of the Jamesoni Chronozone (lower Pliensbachian) (Fig. 1D). This places the site in the transition between the first and second rifting events. The beach of Água de Madeiros exhibits the upper part of the Polvoeira Member and the contact with the Praia de Pedra Lisa Member of the same formation. The fossil was found in the Polvoeira Member (upper Sinemurian), which consists of an alternation of marls and limestones intercalated with black shales. With regards to the fossil content, these levels are notable for an abundance in ammonites and locally bivalves, in a succession dated between the Raricostatum and Jamesoni chronozones (Duarte et al. 2008, 2014). It is also worth noting that the skull ML 2750 is not the first described remain of a marine reptile in the São Pedro de Moel area. In older deposits from the Coimbra Formation (lower Sinemurian) of Praia da Concha (Fig. 1), a new taxon of plesiosaur, Plesiopharos moelensis, was recently described (Puértolas-Pascual et al. 2021).
Material and methods
The limestone block of the ML 2750 specimen displays characteristics and fossiliferous content consistent with the rocks of the Polvoeira Member (upper Sinemurian). It was discovered by Isabel M. Roldão (Leiria, Portugal) on the 8.04.2021, while preparing a field outing for secondary school students. The specimen is currently kept in the collections of the Museu da Lourinhã (ML, Lourinhã, Lisboa, Portugal).
The preparation of ML 2750 was conducted in two distinct phases: in the first, tools such as a chisel and hammer were employed, to remove the matrix and expose the bone; in the second, at the laboratory of ML, the fossil was cleaned with a solution of 50% acetone + 50% water, applied with a cotton swab. Subsequently, some small bone fragments were consolidated with 5% concentration of Paraloid B-72 in acetone and, later, these fragments were glued to the fossil with 50% concentration of Paraloid B-72 in acetone. Then, a micro air pneumatic hammer (air scribe) (HW65) was used to substantially decrease the thickness and width of the limestone block. In the last stage of preparation, a finer mechanical preparation was made, using a micro air pneumatic hammer (HW322) to remove the matrix between the teeth, with the aid of a stereo microscope. To expose more teeth, it was necessary to change the tools used to avoid damage. Thus, the matrix between the teeth of the specimen was moistened with the aid of a cotton swab soaked with a solution of 50% alcohol + 50% acetone, and bamboo sticks were used to facilitate its removal. The reason for changing the solution used was because it was discovered through the magnifying glass that the matrix contained pyrite.
Systematic palaeontology
Ichthyopterygia Owen, 1859
Ichthyosauria Blainville, 1835
Parvipelvia Motani, 1999
Neoichthyosauria Sander, 2000
Thunnosauria Motani, 1999
Baracromia Fischer et al., 2013
Genus Gadusaurus nov.
Zoobank LSID: urn:lsid:zoobank.org:act:36051285-11C9-47FA-A001 -32C9F98B793C.
Type species: Gadusaurus aqualigneus sp. nov., see below; monotypic.
Etymology: From combination of Latin gadu, codfish, and Ancient Greek, sauros, lizard; in reference to both the ichthyosaur fish-like body shape, and the proclivity for codfish in Portuguese cuisine.
Diagnosis.—As for the monotypic type species.
Stratigraphic and geographic range.—Type locality and horizon only.
Gadusaurus aqualigneus sp. nov.
Zoobank LSID: urn:lsid:zoobank.org:act:FB6DFDD0-E0D8-4FC5-B2 A4-9D319F5D6B07.
Etymology: From Latin aqua, water and lignum, wood; in reference to Água de Madeiros, which roughly translates to “water of woods”, the beach where the specimen was found.
Holotype: ML 2750, nearly complete skull (Fig. 2), in a flattened state, the right side fully observable in lateral view, and elements of the left side observable in dorsal view.
Type locality: Praia de Água de Madeiros, São Pedro de Moel, Municipality of Marinha Grande, (Portugal). Coordinates 39°44’28.20”N, 9°2’20.92”W.
Type horizon: Upper part of the Polvoeira Member of the Água de Madeiros Formation, upper Sinemurian (Lower Jurassic).
Diagnosis.—Small-sized ichthyosaur, with an estimated length of approximately 2 m, diagnosed by the following combination of ambiguous synapomorphies: absence of a supranarial process in the premaxilla; and the parietal foramen being entirely surrounded by the frontals. Gadusaurus aqualigneus gen. et sp. nov. is also characterized by the following potential autapomorphies within Parvipelvia: presence of a subcircular depression in the lacrimal just below the posterior region of the external naris (Fig. 3C); big and well-developed elliptical internasal foramen with about 5 times bigger surface area than that of the parietal foramen (Fig. 3A); presence of a well-marked longitudinal groove on each side of the anterolateral region of the excavatio internasalis (Fig 3B).
Description.—The specimen comprises a nearly complete, flattened cranium, approximately 35.2 cm in length (Table 1) with most of the posterior elements up to the posterior edge of the orbit being absent (Fig. 2). There are some disarticulated posterior elements observable, but they are too damaged to identify. The right side of the skull is fully visible in lateral view while only some elements of the left side of the rostrum and forehead are observable, the latter in dorsal view due to the flattened state of the specimen. This flattened state has also caused elements from dorsal edge of the orbit to fold into the orbital area.
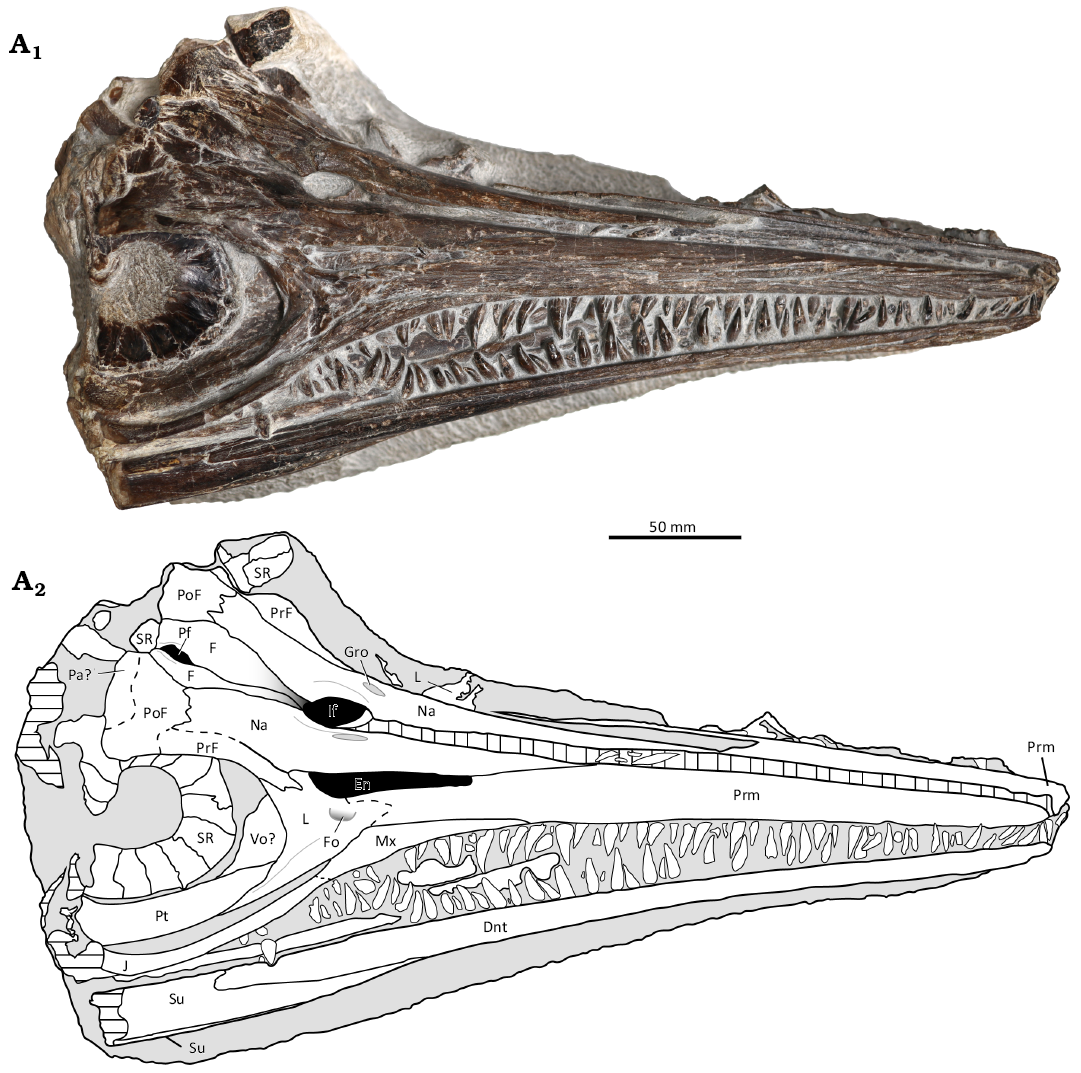
Fig. 2. Ichthyosaur reptile Gadusaurus aqualigneus gen. et sp. nov. (ML 2750) from from the Lower Jurassic of São Pedro de Moel (Marinha Grande, Portugal). Nearly complete skull in laterodorsal view. Photograph (A1), explanatory drawing (A2). Black area represent the open space of the foramens and narial opening. Abbreviations: An, angular; Dnt, dentary; En, external naris; Fo, foramen; F, frontal; Gro, groove; If, internasal foramen; J, jugal; L, lacrimal; Mx, maxilla; Na, nasal; Pa, parietal; Pt, pterygoid; Pf, parietal foramen; PoF, postfrontal; PrF, prefrontal; Prm, premaxilla; SR, sclerotic ring; Su, surangular; Vo, vomer.
Premaxilla: The right premaxilla is straight and narrows from the base to the tip of the rostrum. The supranarial process is absent, while the subnarial process is over half the length of the naris. Striations (longitudinal grooves) are observable along the tip of the premaxilla. A groove is present in the contact with the maxilla, growing increasingly shallower from the posterior to the anterior end. The left premaxilla is disarticulated and very narrow in dorsal view.
Maxilla: The left maxilla is not visible in this specimen. The right maxilla bears teeth and is reduced in size, having less than half the length of the rostrum. The bone reaches its maximum dorsoventral thickness under the external naris region, while both the premaxillary and the jugal processes of the maxilla become narrower and wedged, ending in an anterior and posterior tip respectively. Both processes extend beyond the anterior and posterior margins of the external naris, but this extension is no longer than the narial length. There is no contact with either the naris or the prefrontal since the lacrimal separates them widely. There is no ascending process (= dorsal lamina, Motani 1999; = processus postnarialis, Maisch and Matzke 2000; = postnarial process, Maxwell and Cortés 2020) posterior to the narial opening. Anteriorly it is overlapped by the premaxilla while posteriorly it contacts the jugal and the lacrimal. The jugal process of the maxilla is moderately elongated, however it does not extend to the anterior orbital margin and is considerably shorter than the suborbital process of the lacrimal. Slight striations are visible along the contacts with the premaxilla and the lacrimal.
Nasal: Both nasals are observable. There is a slight depression on the posterior end of the nasals, more observable on the left dorsal view. This is identified as the excavatio internasalis. Within the excavatio internasalis and dorsal to the naris, there is a visible wide elliptical gap between the nasals which has been identified as the internasal foramen. Both nasals are slightly separated by taphonomy and the foramen would not be as wide as it appears. If we consider them sagittally united, this foramen would still be quite large, having a surface area about 5 times larger than the parietal foramen. Considering the two nasals united sagittally, the internasal foramen would be completely surrounded by the nasals, with no involvement of the frontal bone, although it almost contacts the posterior margin of the foramen. On both nasals, next to the anterolateral margins of the internasal foramen, two parallel longitudinal elliptical grooves are observable (Fig. 3C). The left nasal, observable in a dorsal view due to the flattened state of the skull, has reduced anteroposterior exposure, comprising less than half the length of the prenarial rostrum, while its contact with the left premaxilla is disarticulated. The right nasal, visible in lateral view, reaches back over the orbit. It borders the naris dorsally, and has extensive contact with the postfrontal, separating the prefrontal from the frontal entirely. The difference in anterior extent between the left and right nasal prevents us from accurately determining the real anterior extension of the nasals along the snout. However, the well-articulated suture of the right nasal with the premaxilla seems to indicate that its anteroposterior extension would be short (less than 40% of the prenarial rostrum).
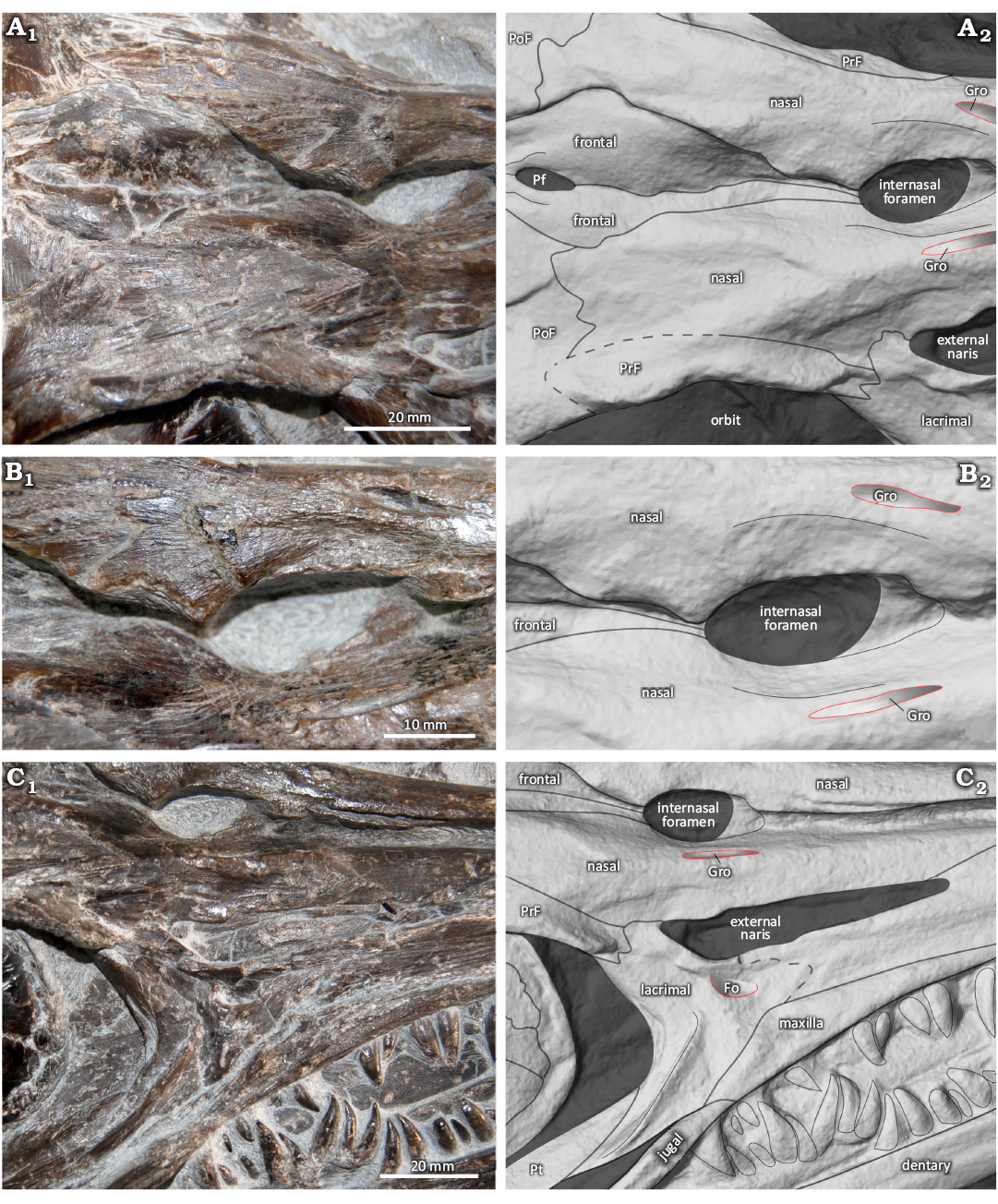
Fig. 3. Ichthyosaur reptile Gadusaurus aqualigneus gen. et sp. nov. (ML 2750) from the Lower Jurassic of São Pedro de Moel (Marinha Grande, Portugal). A. Parietal foramen, surrounded by the frontals, in dorsal view. B. Internasal foramen, with nasal markings on each side, in dorsal view. C. Semi-circular indentation of the lacrimal, in dorsal view. Photographs (A1–C1) and explanatory drawings (A2–C2). Abbreviations: Fo, foramen; Gro, groove; Pt, pterygoid; Pf, parietal foramen; PoF, postfrontal; PrF, prefrontal.
Lacrimal: Due to the flattened state of the specimen, the lacrimal area is very fractured, especially on the anterior end, and details of its anatomy are difficult to discern. The lacrimal contributes to the external naris anteriorly. The external naris is elongated and laterally oriented. There is a small anteroposteriorly oriented elliptical foramen that forms an arch of bone with the posteroventral margin of the external naris. The presence of an arc of bone in the narial region has also been observed in Ichthyosaurus somersetensis Lomax & Massare, 2017 (Massare et al. 2021).
Jugal: The jugal is fractured, with the posterior half being absent, and appears to have a broad anterior margin, covering the posterior end of the maxilla, which ends posterior to the anterior margin of the lacrimal, without reaching the external naris.
Prefrontal: The left prefrontal has some dorsal exposure, mainly on its contact with the nasal, but limited by the anterior process of the postfrontal. The right prefrontal comprises the anterodorsal side of orbit, and is kept from reaching the frontal by the nasal. There is no observable contact with the posterior side of the external naris.
Frontal: While both frontals are observable, only the left frontal has any significant exposure and appears to be dorsally flat, and more exposed at the nasal suture. The posterior-most region of the frontals appears to be eroded, so its relationship with the parietals and supratemporal fenestrae cannot be determined. There is a small elliptical parietal foramen on the posterior end of the frontals (Fig 3A). The foramen seems to be surrounded by the frontals without contacting the parietals. Next to the posterolateral margins of the foramen there are two small ridges. The frontals have a fusiform shape: starting narrow posteriorly, widening until they reach their maximum convexity at the level of the anterior edge of the parietal foramen, and becoming wedged between the nasals anteriorly. The contact with the nasals appears to be slightly disarticulated, forming a deep ridge and a medial groove leading towards the posterior margin of the internasal foramen.
Orbit: The orbit has a regularly rounded anterior shape, with its posterior margin not being preserved. Due to the flattened state of the skull, part of the pterygoid, as well as what is tentatively identified as the vomer, can be observed ventrally under the orbit. The sclerotic ring is quite fractured, and almost the entire posterior region is missing. However, at least 14 ossicles have been preserved. Applying the principle of symmetry to the ring, we can estimate that it originally consisted of approximately 18 to 20 ossicles, a relatively high number for Ichthyosauria. The size of the orbit and the sclerotic ring is similar; therefore, it would fill almost the entire orbital cavity. Although the orbit is of considerable size, there does not appear to be a marked dorsal inflection at the anterior orbital margin with the snout. Instead, this anterior narrowing is progressive.
Lower jaw: The lower jaw is well developed relative to the upper jaw, both in length and in cross-section. It has a straight ventral margin, and the dentary fossa is visible running across the length of the dentary, which has no discernible labial shelf. The surangular is visible in lateral view on the posterior end of the jaw, but it is fractured and lacking it posterior half. The angular is also visible, but it has almost no lateral exposure compared to the surangular.
Dentition: The dentition is fairly complete and well-developed, with aulacodont implantation and no discernible bony fixation. 88 teeth are observable laterally in the right side of the skull, displayed in a single row in each jaw, with a height range of 4 to 15 mm (Table 1). Eleven are observable dorsally at the edges of the left premaxilla. The teeth are conical and round in cross-section, with a well-defined base for the enamel layer, which is thin and smooth. At the root of the teeth, there are observable grooves.
Table 1. Measurements (in mm) of ML 2750.
| |
Length |
Height |
Width |
Estimated diameter |
|
Ventral to the orbit |
352 |
|
|
|
|
Dorsal to the orbit |
344 |
|
|
|
|
Preorbital segment |
280 |
|
|
|
|
Prenarial segment |
231 |
|
|
|
|
Lower jaw |
340 |
|
|
|
|
Skull |
|
165 |
|
|
|
Rostrum |
|
80 |
|
|
|
Upper jaw |
|
60 |
|
|
|
Lower jaw |
|
20 |
|
|
|
Orbit |
|
|
|
75 |
|
Sclerotic ring |
|
|
|
60 |
|
Teeth |
|
4–15 |
2–4 |
|
|
Tooth crown |
|
3–10 |
|
|
|
Basal crown |
|
|
2–4 |
|
Stratigraphic and geographic range.—Type horizon and locality only.
Phylogenetic analysis
The phylogenetic analysis was performed using TNT version 1.6 (Goloboff and Morales 2023). The specimen was added to the dataset of Laboury et al. (2022), and the scoring was done according to the same, and using Hupehsuchus nanchangensis Young, 1972, as the outgroup taxon. We performed two different analyses using a New Technology search. In both analyses some taxa have been grouped into higher-rank clades to reduce the size of the tree and make it easier to read. The complete trees have been included in the SOM (Supplementary Online Material available at http://app.pan.pl/SOM/app70-Pratas_etal_SOM.pdf).
The first analysis (Fig. 4) was conducted without implied weights, holding 1 000 000 trees in memory, with a Ratchet of 200 iterations, a Drift of 10 cycles, finding minimal length 5 times (hits), and initial add. sequences with 10 trees per replication. The resulting most parsimonious trees were then analyzed with the iter-PCR script (Pol and Escapa 2009) to remove problematic taxa from the strict consensus tree (Fig. 4), with a final result of 42 taxa pruned. For this, we used the Iter-PCR tool in TNT leaving its default values, forcing Gadusaurus aqualigneus gen. et sp. nov., not to be pruned, and saving the prunes in a group that was later excluded when building the strict consensus tree. The bootstrap frequencies over 50% were summarized in the strict consensus tree (Fig. 4).

Fig. 4. Phylogenetic relationships of Gadusaurus aqualigneus gen. et sp. nov. (ML 2750) by New Technology search method in TNT, based on the matrix of Laboury et al. (2022). Strict consensus (condensed) tree of 13 most parsimonious trees with 1639 evolutionary steps. Numbers of each node indicate the bootstrap frequencies over 50%. Some taxa have been grouped into higher-rank clades (complete version in SOM) and 42 taxa have been excluded using the Iter-PCR tool.
The first phylogenetic analysis (Fig. 4) returns 13 most parsimonious trees (MPT) with a length of 1639 steps. It has a consistency index (CI) of 0.199, a retention index (RI) of 0.662, and a rescaled consistency index (RC) of 0.132. Gadusaurus aqualigneus gen. et sp. nov. is recovered with two ambiguous synapomorphies: the premaxilla lacks a supranarial process (char. 2 = 1) the parietal foramen is completely surrounded by the frontals (char. 50 = 2).
In the strict consensus tree obtained in the first analysis (Fig. 4), ML 2750 is recovered as part of a polytomy within the clade Baracromia, and affecting the following taxa: Gadusaurus aqualigneus gen. et sp. nov., Temnodontosaurus eurycephalus McGowan, 1974, a clade composed of the remaining Temnodontosaurus spp., the clade Eurhinosauria, the clade composed of Hauffiopteryx spp., and the clade composed of Stenopterygius spp. The analysis also places Stenopterygiidae as the sister clade of Hauffiopteryx, which is in accordance with various previous cladistic analyses (Fischer et al. 2016; Moon 2019; Maxwell and Cortés 2020).
For the second analysis (Fig. 5), the same protocol was followed, but using extended implied weighting with a k = 12 (commands “piwe=” and “xpiwe(*0.5 < 5 /12)”). This method assigns greatest importance to characters that are most likely to be homologous. Therefore, it usually results in improved clade supports and better recovery of taxonomic groups with fewer polytomies (Goloboff 2014).
This second phylogenetic analysis returned 8 most parsimonious trees (MPT) with a best score of 61.72684. It has a consistency index (CI) of 0.196, a retention index (RI) of 0.655, and a rescaled consistency index (RC) of 0.128. The iter-PCR script only pruned 2 taxa: Pervushovisaurus bannovkensis Arkhangelsky, 1998, and Undorosaurus trautscholdi Arkhangelsky & Zverkov, 2014.
The strict consensus tree obtained in the second analysis (Fig. 5) is much better resolved, eliminating almost all polytomies. As a result, Gadusaurus aqualigneus gen. et sp. nov. also appears within Baracromia, but as the sister taxon to Chacaicosaurus cayi Fernández, 1994, and the clade formed by both appears as the sister group to Ophthalmosauria.

Fig. 5. Phylogenetic relationships of Gadusaurus aqualigneus gen. et sp. nov. (ML 2750) by New Technology search method in TNT using extended implied weighting (k = 12), based on the matrix of Maxwell and Cortés (2020). Strict consensus tree of 8 most parsimonious trees with best score 61.72684. Numbers of each node indicate the bootstrap frequencies over 50%. Some taxa have been grouped into higher-rank clades (complete version in SOM) , and 2 taxa have been excluded using the Iter-PCR tool.
Discussion
Both phylogenetic analysis recovered ML 2750 as part of the clade Baracromia. The node-based definition of Baracromia was erected by Fischer et al. (2013) as the clade that includes Stenopterygius quadriscissus Quenstedt, 1858, and Ophthalmosaurus icenicus Seeley, 1874, and all descendants of their most recent common ancestor, but not Ichthyosaurus communis. Within Baracromia, Acamptonectes, and Platypterygius are documented from the Cretaceous (Kuhn 1946; Fischer et al. 2012). Brachypterygius, Cryopterygius, Grendelius, Ophthalmosaurus, and Palvennia, are genera documented from the Late Jurassic (Férnandez and Campos 2015; Tyborowski 2016; Moon and Kirton 2018). It can thus be inferred that ML 2750, found in units from the upper Sinemurian, is unlikely to belong to or be closely related to any of them. To clarify the phylogeny of ML 2750, comparisons of its cranial elements with those of other genera from the Early Jurassic within Baracromia will be required, and so those are the diagnostical characters that will be focused on. Protoichthyosaurus and Ichthyosaurus are two genera outside Baracromia but part of its sister clade, Ichthyosauridae, and still coeval with ML 2750, and so they will also be compared with the specimen.
Comparisons.—Eurhinosaurus and Excalibosaurus are two Early Jurassic genera, from the Toarcian of Germany, France, Switzerland, and England, and Sinemurian of England respectively (McGowan 1994, 2003; Maisch 2010; Reisdorf et al. 2011), which are best identified by the upper jaw being much longer than the lower jaw, which only makes 1/2 the full length of the skull in Eurhinosaurus and 3/4 in Excalibosaurus, resulting in a considerable overbite (McGowan 1989). While the anterior end of the rostrum of ML 2750 has some damage, including a noticeable split between the right and the left premaxilla, the size difference between upper and lower jaw seems negligible and there is no distinct overbite.
Wahlisaurus is a genus from the Hettangian of England, which, alongside Excalibosaurus, Eurhinosaurus, and Leptonectes, belongs to the often recovered family Leptonectidae Maisch, 1998 (Lomax 2017). Its defining characteristic being the noticeable overbite formed by the lower jaw being shorter than the dentary, while its type and only species Wahlisaurus massarae Lomax, 2017, is mostly identified by postcranial elements, as well as long and slender teeth with marginally recurved crowns and bulbous, infolded roots (Lomax 2017). ML 2750 lacks any such overbite, and its teeth are noticeably smaller and less slender.
Leptonectes is a genus that ranges from the Rhaetian to the Pliensbachian, and like the others of the Leptonectidae clade, it is characterized by the overbite caused by a size disparity between the upper and lower jaw, if one that is less significant than in the previously discussed genus (McGowan and Milner 1999). The genus is comprised of the Leptonectes tenuirostris Conybeare, 1822, Leptonectes solei McGowan, 1993, and Leptonectes moorei McGowan & Milner, 1999. Leptonectes tenuirostris and L. solei have both been observed to have long and slender snouts with relatively small teeth, unlike the shorter snout of ML 2750. Leptonectes moorei has a noticeably shorter snout, more comparable with ML 2750. However, the external naris is rounder and more complex, with a noticeable notch in the posterior dorsal margin, as well as a more developed maxilla, unlike elongated external naris and markedly reduced maxilla in ML 2750.
Stenopterygius is a genus that ranges from Toarcian to the Aalenian of Germany, Luxembourg, France, Belgium, and England. The cranial elements that identify the genus include the size of the temporal fenestrae; a suborbital groove of the surangular; the exposure of the angular on posterior lower jaw; and the lack of ornamentation of the crown of the teeth (Maisch 2010; Maxwell 2012). It is comprised of four species, Stenopterygius quadriscissus, Stenopterygius triscissus Quenstedt, 1856, Stenopterygius uniter, and Stenopterygius aaleniensis Maxwell et al., 2012 (Maxwell 2012; Maxwell et al. 2012). In all four, with some intraspecific variability, the premaxilla sports a supranarial process, and the maxilla contacts the external naris, both traits that ML 2750 is lacking.
Suevoleviathan is a genus from the Toarcian of Germany. It is characterized by a small lateral exposure of the maxilla and anterodorsal exposure of the nasal; large upper temporal fenestrae; a broad temporal region; and a well-developed, heterodont dentition that is reduced posteriorly (Maxwell 2018). Unlike this genus, ML 2750 sees little to no reduction of the posterior dentition, the teeth are lacking rough enamel ornamentation, the angular has very reduced exposure laterally, and the premaxilla is lacking a supranarial process.
Temnodontosaurus is a genus from the Hettangian to the Toarcian of from England, Belgium, France, Germany, Luxenbourg and possibly Chile (Swaby and Lomax 2021; Laboury et al. 2022). The genus is diagnosed by its significant size (7–12 m); long and robust skull; contact between the nasal and parietal; maxilla with long anterior process; relative small orbit; and relatively large teeth (Swaby and Lomax 2021). While the premaxillary process of the maxilla of ML 2750 does continue anterior to the external naris, it does not do so to the same extent that is documented in Temnodontosaurus, and the premaxilla is lacking the supranarial process that is observable within this genus.
Hauffiopteryx is a genus from the Pliensbachian to the Toarcian, with two recognized species, Hauffiopteryx typicus Huene, 1931, and Hauffiopteryx altera Maxwell & Cortés, 2020. The genus is diagnosed by a relatively short and slender antorbital rostrum; a slight overbite; the prefrontal participating in the external naris; nasals with a greater pre-narial exposure dorsally than the premaxilla; prefrontals with larger external exposure dorsally than the postfrontals; the parietal foramen located anterior to the supratemporal fenestra; supratemporal fenestra small and circular in dorsal view; supratemporal palmate in posterodorsal view; and teeth with smooth enamel (Maxwell and Cortés 2020). ML 2750 shares some similarities with the genus, including a lack of a supranarial process in the premaxilla, but its antorbital rostrum is slender, there is no contact between the prefrontal and the external naris, and the anterior margin of the jugal is broader and covers part of the maxilla instead of tapering.
Protoichthyosaurus is a genus from the Hettangian, possibly ranging from the Rhaetian to the Sinemurian. It is considered the sister taxon of Ichthyosaurus. The distinction between the two genera is made by fin elements such as the number of elements in the carpal row. It has two recognized species, Protoichthyosaurus prostaxalis Appleby, 1979, and Protoichthyosaurus appleby Lomax et al., 2017 (Lomax et al. 2017). This genus is diagnosed by cranial elements such as the parietal foramen being bordered posteriorly by the parietals; the anterior process of the prefrontal separating the lacrimal from the orbital margin, partially or completely; a large, triangular external naris; an asymmetrical maxilla with an elongated anterior process; and prominent grooves at the root of the teeth that extend to the base of the crown (Lomax and Massare 2018). In ML 2750, the foramen is completely surrounded by the frontals, and the anterior process of the prefrontal doesn’t limit the participation of the lacrimal in the orbital margin too extensively from what can be observed.
Ichthyosaurus is the most common Early Jurassic ichthyosaur genus, ranging from the Hettangian to the Pliensbachian and being very commonly found in England, while also occurring in Wales, Ireland, Belgium, Germany, and Portugal (Massare and Lomax 2018; Sousa and Mateus 2021). The genus is comprised of the currently recognized species Ichthyosaurus anningae Lomax & Massare, 2015, Ichthyosaurus breviceps Owen, 1881, Ichthyosaurus communis, Ichthyosaurus conybeari Lydekker, 1888, Ichthyosaurus larkini Lomax & Massare, 2017, and Ichthyosaurus somersetensis. ML 2750 shares some observable similarities and dissimilarities with the various species of this genus. It lacks a supranarial process of the premaxilla as in I. anningae, while having a larger subprocess of the premaxilla and a broader anterior margin of the jugal. The ornamentation of the enamel is thin and smooth as in I. breviceps, while the maxilla is located more anteriorly in ML 2750, with a shorter jugal process. The jugal has a broad anterior process, and frontals have a convex anterior edge at the parietal foramen, as in I. larkini. But unlike I. larkini, ML 2750 lacks a supranarial process of the premaxilla and has a shorter jugal. The parietal foramen is completely surrounded by the frontals as in I. somersetensis, while the enamel lacks the ornamentation of grooves and ridges, and the crown of the teeth is circular. The teeth of ML 2750 have observable grooves in the roots, unlike what is observed in species of Ichthyosaurus. These disparities lead us to conclude it is unlikely for ML 2750 to belong to this genus.
Chacaicosaurus is an early Bajocian genus from Argentina. It is comprised of its type and only species, Chacaicosaurus cayi. It is identified by cranial elements such as an elongated, slender snout; a slender lower jaw; a large naris; a small orbit; a complete loss or extreme loss of teeth in mature individuals (Fernández 1994). While the maturity of ML 2750 is ambiguous, there is no observable reduction or loss of teeth. This, along with the disparity in age, makes it unlikely for ML 2750 to belong to the genus.
These comparisons, along with the previously mentioned autapomorphies, make us posit ML 2750 as an entirely new genus and species, Gadusaurus aqualigneus gen. et sp. nov., the first to be documented in the Iberian Peninsula.
Ontogenetic remarks.—Recent studies on the ontogeny of ichthyosaurs, using Stenopterygius quadriscissus as a reference, have identified four prenatal and three postnatal stages of ontogenetic development. Although many of the sutures observed in ML 2750 appear to be well fused, sometimes even indistinguishable, Miedema and Maxwell (2022) notably indicate that an unfused midline of the cranium is a sign of sexual immaturity. While this open midline is observable in ML 2750, making it possible that the specimen is a juvenile or a subadult, it is difficult to discern how much of this is due to taphonomic deformation.
Paleobiogeographic implications.—Various ichthyosaur genera of the Early Jurassic, are known to have existed during the early Toarcian Oceanic Anoxic Event, which had a significant impact on the composition and abundance of both vertebrate and invertebrate faunas in locations such as the southwest German basins. Hauff (1921), and Maxwell and Vincent (2016), have documented this event thoroughly. It has been suggested that Early Jurassic ichthyosaurs had a specific diet or hunting strategy during this period, due to changes in the availability of prey or environmental circumstances, including bottom-water anoxia that could coerce fish and belemnites into a limited area near the surface of the ocean (Ullmann et al. 2014).
The specimen ML 2750 was found in Polvoeira Member of the Água de Madeiros Formation of the late Sinemurian age, which places it chronologically prior to the Toarcian Oceanic Anoxic Event. The section of the Água de Madeiros Formation, where the specimen was discovered, is marked by the Raricostatum Chronozone. This chronozone is known to represent a major transgressive event that occurred in this time period, which is supported by the increase in nektonic fauna and organic deposition in the upper part of the Polvoeira Member (Duarte et al. 2014). It can be postulated that such a transgressive event in southern Europe would have had an analogous effect on the lifestyle of Gadusaurus aqualigneus gen. et sp. nov. as the Toarcian Oceanic Anoxic Event had for the latter Posidonia Shale Sea ichthyosaur fauna in northern areas such as England and Germany, thereby creating a region rich in prey items. Nevertheless, this remains a preliminary hypothesis, and further studies linking these events and their paleobiogeographical and paleoecological consequences are still necessary. Understanding the ecological responses of Gadusaurus aqualigneus gen. et sp. nov. during transgressive or anoxic events like those evidenced in both southern Europe and northern regions could provide crucial insights into the adaptive strategies of this ichthyosaur species and the broader impacts of Jurassic environmental changes.
Conclusions
ML 2750, a specimen uncovered in an upper Sinemurian outcrop of São Pedro de Moel in the central western coast of Portugal, was identified as a new genus and species. It exhibits distinct synapomorphies in the lack of a supranarial process in the premaxilla and the parietal foramen being completely surrounded by the frontals. It further shows two autapomorphies: a big semi-circular foramen in the lacrimal bone, and elliptical nasal grooves on either side of the internasal foramen. These characters have led to the identification of the specimen as the new species Gadusaurus aqualigneus gen. et sp. nov. The discovery of this specimen represents the most complete and best-preserved remains of an ichthyosaur described to date in the Iberian Peninsula. As a result, it is also the first new species of this group of marine reptiles described in Iberia (Portugal and Spain). Having been found in units of the late Sinemurian age, this specimen precedes the Toarcian Oceanic Anoxic Event that is theorized to have affected the lifestyle of later ichthyosaur genera. It can be hypothesized that the transgressive events that occurred in this time period may have had similar effects on the lifestyle of this Sinemurian species, but further research is required.
Acknowledgements
We would like to thank Erin Maxwell (Staatliches Museum für Naturkunde, Stuttgart, Germany) for her advice regarding some anatomical aspects of the specimen, Luís V. Duarte (Universidade de Coimbra, Portugal) for his assistance regarding the stratigraphy of the locality, Carla Alexandra Tomás and Micael Martinho (both Museu da Lourinhã, Portugal) for their help in the preparation of the specimen. Erin Maxwell and Feiko Miedema (both Staatliches Museum für Naturkunde, Stuttgart, Germany) are thanked for their reviews. Thanks to the projects from the Fundação para a Ciência e Tecnologia (FCT) of Portugal: BioGeoSauria (PTDC/CTA-PAL/2217/2021), and Research Unit GeoBioTec (UIDB/04035/2020). JPS was founded by a Portuguese PhD grant from the FCT (bolsa 2021.08727.BD); MRI was funded by CEECIND/ 02199/2018. EPP was funded by SFRH/BPD/116759/2016 (FCT, Portugal), PID2021-122612OB-I00 (Ministerio de Ciencia e Innovación, Spain), E18_23R, Grupos Consolidados (Gobierno de Aragón) and by a postdoctoral contract María Zambrano (Ministerio de Universidades, Spain, through the Next Generation EU funds of the European Union).
Editor: Daniel Barta.
References
Alves, T.M., Gawthorpe, R.L., Hunt, D. W., and Monteiro, J.H. 2002. Jurassic tectono-sedimentary evolution of the Northern Lusitanian Basin (offshore Portugal). Marine and Petroleum Geology 19: 727–754. Crossref
Arkhangelsky, M.S. 1998. On the ichthyosaurian genus Platypterygius. Paleontological Journal 32: 611–615.
Arkhangelsky, M.S. and Zverkov, N.G. 2014. On a new ichthyosaur of the genus Undorosaurus. Trudy Zoologičeskogo Instituta RAN 318 (3): 187–196. Crossref
Appleby, R.M. 1979. The affinities of Liassic and later ichthyosaurs. Palaeontology 22: 921–946.
Bardet, N., Pereda Suberbiola, X., and Ruiz Omeñaca, J.I. 2008. Mesozoic marine reptiles of the Iberian Peninsula. Geo-Temas 10: 1245–1248.
Blainville, H.M.D., de 1835. Description de quelques espèces de la Californie, précedée de l’analyse d’un systéme général d’érpetologie et d’amphibiologie. Nouvelles Archives du Muséum d’Histoire Naturelle 4: 236–296.
Beche, H.T., de la and Conybeare, W.D. 1821. Notice of the discovery of a new fossil animal, forming a link between the Ichthyosaurus and Crocodile, together with general remarks on the osteology of the Ichthyosaurus. Transactions of the Geological Society of London 5 (1): 559–594. Crossref
Conybeare, W.D. 1822. Additional notices on the fossil genera Ichthyosaurus and Plesiosaurus. Transactions of the Geological Society of London 1 (1): 103–123. Crossref
Duarte, L.V., Comas-Rengifo, M.J., Silva, R.L., Paredes, R., and Goy, A. 2014. Carbon isotope stratigraphy and ammonite biochronostratigraphy across the Sinemurian–Pliensbachian boundary in the western Iberian margin. Bulletin of Geosciences 89: 719–736. Crossref
Duarte, L.V., Silva, R., Duarte, C.B., Azerêdo, A.C., and Comas Rengifo, M.J. 2009. Litostratigrafia do Jurássico Inferior da região de S. Pedro de Moel (Bacia Lusitânica, Portugal). In: A Terra, Conflitos e Ordem. Homenagem ao Professor Ferreira Soares, 175–185. Museu Mineralgógico e Geológico da Universidade de Coimbra, Coimbra.
Duarte, L.V., Wright, V.P., Fernandez-Lopez, S., Elmi, S.., Krautter, M., Azerêdo, A.C., Henriques, M.H., Rodrigues, R., and Perilli, N. 2004. Early Jurassic carbonate evolution in the Lusitanian Basin: facies, sequence stratigraphy and cyclicity. In: L.V. Duarte and M.H. Henriques (eds.), Carboniferous and Jurassic Carbonate Platforms of Iberia. 23rd IAS Meeting of Sedimentology, Coimbra 2004, Field Trip Guide Book Volume 1, 45–71. International Association of Sedimentologists, Coimbra.
Fernandez, M. 1994. A new long-snouted ichthyosaur from the Early Bajocian of Neuquén Basin (Argentina). Ameghiniana 31: 283–290.
Fernández, M.S. and Campos, L. 2015. Ophthalmosaurids (Ichthyosauria: Thunnosauria): Alpha taxonomy, clades and names. Reptiles Extintos Volumen en Homenaje a Zulma Gasparini. Publicación Electrónica de la Asociación Paleontológica Argentina 15 (1): 20–30. Crossref
Fernández, M.S., Piñuela, L., and García-Ramos, J.C. 2018. First report of Leptonectes (Ichthyosauria: Leptonectidae) from the Lower Jurassic (Pliensbachian) of Asturias, northern Spain. Palaeontologia Electronica 21: 21.2.29A. Crossref
Ferreira, O.V. 1958. Novos restos de “Ichthyosauridae” e “Stenopterygidae” encontrados no Lias de Portugal. Comunicações dos Serviços Geológicos de Portugal 42: 175–181.
Fischer, V., Appleby, R.M., Naish, D., Liston, J., Riding, J.B., Brindley, S., and Godefroit, P. 2013. A basal thunnosaurian from Iraq reveals disparate phylogenetic origins for Cretaceous ichthyosaurs. Biology Letters 9 (4): 20130021. Crossref
Fischer, V., Bardet, N., Benson, R.B., Arkhangelsky, M.S., and Friedman, M. 2016. Extinction of fish-shaped marine reptiles associated with reduced evolutionary rates and global environmental volatility. Nature Communications 7 (1): 10825. Crossref
Fischer, V., Maisch, M.W., Naish, D., Kosma, R., Liston, J., Joger, U., and Appleby, R.M. 2012. New ophthalmosaurid ichthyosaurs from the European Lower Cretaceous demonstrate extensive ichthyosaur survival across the Jurassic–Cretaceous boundary. PLoS One 7 (1): e29234. Crossref
Fortuny, J., Bolet, A., Sellés, A.G., Cartanyà, J., and Galobart, À. 2011. New insights on the Permian and Triassic vertebrates from the Iberian Peninsula with emphasis on the Pyrenean and Catalonian basins. Journal of Iberian Geology 37: 65–86. Crossref
Goloboff, P.A. 2014. Extended implied weighting. Cladistics 30: 260–272. Crossref
Goloboff, P.A. and Morales, M.E. 2023. TNT version 1.6, with a graphical interface for MacOS and Linux, including new routines in parallel. Cladistics 39: 144–153. Crossref
Hauff, B. 1921. Untersuchung der Fossilfundstatten von Holzmaden im Posidonienschiefer des Oberen Lias Wurttembergs Fossilfundstatten. Palaeontographica 64: 1–42.
Huene, F.V., von 1931. Neue Studien uber Ichthyosaurier aus Holzmaden. Abhandlungen der Senkenbergischen Naturforschenden Gesellschaft Band 42 (4): 1–38.
Knoll, F., López-Antoñanzas, R., and Molina-Anadón, J.A. 2004. Filling a gap: Late Triassic nonmarine tetrapods from the Iberian Peninsula. Journal of Vertebrate Paleontology 24 (3): 79A.
Kuhn, O. 1946. Ein skelett von Ichthyosaurus hercynicus n. sp. aus dem Aptien von Gitter. Berichte der Naturforschenden Gesellschaft Bamberg 29: 69–82.
Laboury, A., Bennion, R.F., Thuy, B., Weis, R., and Fischer, V. 2022. Anatomy and phylogenetic relationships of Temnodontosaurus zetlandicus (Reptilia: Ichthyosauria). Zoological Journal of the Linnean Society 195: 172–194. Crossref
Lomax, D. R. 2017. A new leptonectid ichthyosaur from the Lower Jurassic (Hettangian) of Nottinghamshire, England, UK, and the taxonomic usefulness of the ichthyosaurian coracoid. Journal of Systematic Palaeontology 15: 387–401. Crossref
Lomax, D.R. and Massare, J.A. 2015. A new species of Ichthyosaurus from the Lower Jurassic of West Dorset, England, UK. Journal of Vertebrate Paleontology 35 (2): e903260. Crossref
Lomax, D.R. and Massare, J.A. 2017. Two new species of Ichthyosaurus from the lowermost Jurassic (Hettangian) of Somerset, England. Papers in Palaeontology 3: 1–20. Crossref
Lomax, D.R. and Massare, J.A. 2018. A second specimen of Protoichthyosaurus applebyi (Reptilia: Ichthyosauria) and additional information on the genus and species. Paludicola 11: 164–178.
Lomax, D.R., Massare, J.A., and Mistry, R.T. 2017. The taxonomic utility of forefin morphology in Lower Jurassic ichthyosaurs: Protoichthyosaurus and Ichthyosaurus. Journal of Vertebrate Paleontology 37 (5): e1361433. Crossref
Lydekker, R. 1888. Note on the classification of the Ichthyopterygia (with a notice of two new species). Geological Magazine 5 (7): 309–314. Crossref
Massare, J.A. and Lomax, D.R. 2018. A taxonomic reassessment of Ichthyosaurus communis and I. intermedius and a revised diagnosis for the genus. Journal of Systematic Palaeontology 16: 263–277. Crossref
Massare, J.A., Wahl, W.R., and Lomax, D.R. 2021. Narial structures in Ichthyosaurus and other Early Jurassic ichthyosaurs as precursors to a completely subdivided naris. Paludicola 13: 128–139.
Matamales-Andreu, R., Oms, O., Galobart, À., and Fortuny, J. 2021. Middle–Upper Triassic marine vertebrates of Mallorca (Balearic Islands, western Mediterranean). Historical Biology 33: 2520–2533. Crossref
Maisch, M.W. 1998. A new ichthyosaur genus from the Posidonia Shale (Lower Toarcian, Jurassic) of Holzmaden, SW-Germany with comments on the phylogeny of post-Triassic ichthyosaurs. Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen 209: 47–78. Crossref
Maisch, M.W. 2001. Neue Exemplare der seltenen Ichthyosauriergattung Suevoleviathan Maisch, 1998 aus dem Unteren Jura von Südwestdeutschland. Geologica et Palaeontologica 35: 145–160.
Maisch, M.W. 2008. Revision der Gattung Stenopterygius Jaekel, 1904 emend. von Huene, 1922 (Reptilia: Ichthyosauria) aus dem unteren Jura Westeuropas. Palaeodiversity 1: 227–271.
Maisch, M.W. 2010. Phylogeny, systematics, and origin of the Ichthyosauria—the state of the art. Palaeodiversity 3: 151–214.
Maisch, M.W. and Matzke, A.T. 2000. The Ichthyosauria. Stuttgarter Beiträge zur Naturkunde Serie B (Geologie und Paläontologie) 298: 1–159.
Maxwell, E.E. 2012. New metrics to differentiate species of Stenopterygius (Reptilia: Ichthyosauria) from the Lower Jurassic of southwestern Germany. Journal of Paleontology 86: 105–115. Crossref
Maxwell, E.E. 2018. Redescription of the ‘lost’ holotype of Suevoleviathan integer (Bronn, 1844) (Reptilia: Ichthyosauria). Journal of Vertebrate Paleontology 38 (2): e1439833. Crossref
Maxwell, E.E. and Cortés, D. 2020. A revision of the Early Jurassic ichthyosaur Hauffiopteryx (Reptilia: Ichthyosauria), and description of a new species from southwestern Germany. Palaeontologia Electronica 23 (2): 1–43. Crossref
Maxwell, E.E. and Vincent, P. 2016. Effects of the early Toarcian Oceanic Anoxic Event on ichthyosaur body size and faunal composition in the Southwest German Basin. Paleobiology 42: 117–126. Crossref
Maxwell, E.E., Fernandez, M.S., and Schoch, R.R. 2012. First diagnostic marine reptile remains from the Aalenian (Middle Jurassic): a new ichthyosaur from southwestern Germany. PLoS ONE 7: e41692. Crossref
McGowan, C. 1974. A revision of the longipinnate ichthyosaurs of the Lower Jurassic of England, with descriptions of two new species (Reptilia: Ichthyosauria). Life Sciences Contributions 97: 1–37. Crossref
McGowan, C. 1989. Computed tomography reveals further details of Excalibosaurus, a putative ancestor for the swordfish-like ichthyosaur Eurhinosaurus. Journal of Vertebrate Paleontology 9: 269–281. Crossref
McGowan, C. 1993. A new species of large, long-snouted ichthyosaur from the English lower Lias. Canadian Journal of Earth Sciences 30: 1197–1204. Crossref
McGowan, C. 1994. The taxonomic status of the Upper Liassic ichthyosaur Eurhinosaurus longirostris. Palaeontology 37: 747–754.
McGowan, C. 2003. A new specimen of Excalibosaurus from the English Lower Jurassic. Journal of Vertebrate Paleontology 23: 950–956. Crossref
McGowan, C. and Milner, A.C. 1999. A new Pliensbachian ichthyosaur from Dorset, England. Palaeontology 42: 761–768. Crossref
Miedema, F. and Maxwell, E.E. 2022. Ontogenetic variation in the skull of Stenopterygius quadriscissus with an emphasis on prenatal development. Scientific Reports 12 (1): 1707. Crossref
Miguel Chavez, C. de, Pérez-García, A., Cobos, A., Royo-Torres, R., Ortega, F., and Alcala, L. 2015. A diverse Late Triassic tetrapod fauna from Manzanera (Teruel, Spain). Geobios 48: 479–490. Crossref
Moon, B.C. 2019. A new phylogeny of ichthyosaurs (Reptilia: Diapsida). Journal of Systematic Palaeontology 17: 129–155. Crossref
Moon, B.C. and Kirton, A.M. 2018. Ichthyosaurs of the British Middle and Upper Jurassic. Part 2. Brachypterygius, Nannopterygius, Macropterygius and taxa invalida. Monographs of the Palaeontographical Society 172 (650): 85–177. Crossref
Motani, R. 1999. Phylogeny of the Ichthyopterygia. Journal of Vertebrate Paleontology 19: 473–496. Crossref
Owen, R. 1881. A monograph of the fossil Reptilia of the Liassic formations. Plesiosaurus, Dimorphodon and Ichthyosaurus. Monographs of the Palaeontographical Society 1861–1881 (3): 83–134. Crossref
Owen, R. 1859. On the orders of fossil and recent Reptilia and their distribution in time. Report of the British Asociation for the Advancement of Science 1859: 153–166.
Pol, D. and Escapa, I.H. 2009. Unstable taxa in cladistic analysis: identification and the assessment of relevant characters. Cladistics 25: 515–527. Crossref
Puértolas-Pascual, E., Marx, M., Mateus, O., Saleiro, O., Fernandes, A.E., Marinheiro, J.O., Tomas, C., and Mateus, S. 2021. A new plesiosaur from the Lower Jurassic of Portugal and the early radiation of Plesiosauroidea. Acta Palaeontologica Polonica 66: 369–388. Crossref
Quenstedt, F.A. 1856–1858. Der Jura. 842 pp. H. Laupp’schen Buchhandlung, Tübingen.
Reisdorf, A.G., Maisch, M.W., and Wetzel, A. 2011. First record of the leptonectid ichthyosaur Eurhinosaurus longirostris from the Early Jurassic of Switzerland and its stratigraphic framework. Swiss Journal of Geosciences 104: 211–224. Crossref
Ruiz-Omeñaca, J.I., García-Ramos, J.C., Piñuela, L., Bardet, N., Bermúdez-Rochas, D.D., Canudo, J.I., and Pereda Suberbiola, X. 2006. Restos directos de vertebrados del Jurásico de Asturias. XXII Jornadas de la Sociedad Española de Paleontología y simposios de los proyectos PICG 493 (503): 499.
Sander, P.M. 2000. Ichthyosauria: their diversity, distribution, and phylogeny. Paläontologische Zeitschrift 74: 1–35. Crossref
Sauvage, H.É. 1898. Vertébrés fossiles du Portugal: contributions à l’étude des poissons et des reptiles du jurassique et du crétacique. 22 pp. L’Académie royale des Sciences, Lisbon.
Seeley, H.G. 1874. On the pectoral arch and fore limb of Ophthalmosaurus, a new ichthyosaurian genus from the Oxford Clay. Quarterly Journal of the Geological Society 30: 699–707. Crossref
Soares, A.F., Rocha, R.B., Elmi, S., Henriques, M.H., Mouterde, R., Almeras, Y., Ruget, C., Marques, J., Duarte, L.V., Carapito, M.C., and Kullberg, J. 1993. Le sous-bassin nord lusitanien (Portugal) du Trias au Jurassique moyen: histoire d’un “rift avorté”. Comptes rendus de l’Académie des sciences. Série II, Mécanique, physique, chimie, sciences de l’univers, sciences de la terre 317: 1659–1666.
Sousa, J. and Mateus, O. 2021. The southernmost occurrence of Ichthyosaurus from the Sinemurian of Portugal. Fossil Record 24 (2): 287–294. Crossref
Suaréz Vega, L.C. 1974. Estratigrafía del jurásico en Asturias. Cuadernos de Geología Ibérica 3: 1–100.
Swaby, E.J. and Lomax, D.R. 2021. A revision of Temnodontosaurus crassimanus (Reptilia: Ichthyosauria) from the Lower Jurassic (Toarcian) of Whitby, Yorkshire, UK. Historical Biology 33: 2715–2731. Crossref
Tyborowski, D. 2016. A new ophthalmosaurid ichthyosaur from the Late Jurassic of Owadów-Brzezinki Quarry, Poland. Acta Palaeontologica Polonica 61: 791–803. Crossref
Ullmann, C.V., Thibault, N., Ruhl, M., Hesselbo, S.P., and Korte, C. 2014. Effect of a Jurassic oceanic anoxic event on belemnite ecology and evolution. Proceedings of the National Academy of Sciences 111: 10073–10076. Crossref
Wilson, R.C.L., Hiscott, R.N., Willis, M.G., and Gradstein, F.M. 1989. The Lusitanian Basin of West-Central Portugal: Mesozoic and Tertiary Tectonic, Stratigraphic, and Subsidence History. Chapter 22: European-African Margins. AAPG Memoir 46: 341–361 Crossref
Young, C.C., and Dong, Z.M. 1972. Chaohusaurus geishanensis from Anhui province. Memoirs of the Institute of Vertebrate Paleontology and Paleoanthropology, Academia Sinica A 9: 11–14.
Zbyszewski, G. and Moitinho de Almeida, F. 1952. Restes d’ichthyosauriens dans de Lias de S. Pedro de Muel. Comunicações dos Serviços Geológicos de Portugal 33: 5–10.
Acta Palaeontol. Pol. 70 (1): 179–192, 2025
https://doi.org/10.4202/app.01199.2024