

New Late Cretaceous zhelestid mammal from the Bayanshiree Formation, Mongolia
TSUKASA OKOSHI, RYUJI TAKASAKI, KENTARO CHIBA, MASAHITO NATORI, MOTOTAKA SANEYOSHI, AKIO TAKAHASHI, SHOTA KODAIRA, SHOJI HAYASHI, SHINOBU ISHIGAKI, BUUVEI MAINBAYAR, and KHISHIGJAV TSOGTBAATAR
Okoshi, T., Takasaki, R., Chiba, K., Natori, M., Saneyoshi, M., Takahashi, A., Kodaira, S., Hayashi, S., Ishigaki, S., Mainbayar, B., and Tsogtbaatar, K. 2025. New Late Cretaceous zhelestid mammal from the Bayanshiree Formation, Mongolia. Acta Palaeontologica Polonica 70 (1): 193–203.
Numerous exquisitely preserved mammal fossils unearthed from Upper Cretaceous strata in the Gobi Desert of Mongolia have played a key role in understanding Mesozoic mammalian evolution. These splendid mammal fossils have been recovered mainly from the Baruungoyot and Djadokhta formations, but only two fragmentary remains of mammals have been collected from the underlying Bayanshiree Formation. Here we report a new species of Zhelestidae, Ravjaa ishiii gen. et sp. nov., based on a new mammal specimen recently discovered from the Bayanshiree Formation at the Bayan Shiree locality. The new specimen is represented by a well-preserved partial right dentary with the distal portion of an ultimate premolar and the first to the third molars. The gross observation and the phylogenetic analyses demonstrate zhelestid affinities: the subequally tall protoconid and metaconid, closely approximated hypoconulid and entoconid. Zhelestidae was widely distributed from Eurasia to North America in the Late Cretaceous but has not previously been recovered in the abundant Late Cretaceous mammalian fauna of Mongolia. This species represents the first zhelestid from the Mongolian Upper Cretaceous strata and contributes to further understanding of their paleobiogeographic and ecological insights.
Key words: Mammalia, Eutheria, Cretaceous, Bayanshiree Formation, Mongolia.
Tsukasa Okoshi [r23ndd7ap@ous.jp; ORCID: https://orcid.org/0009-0001-4884-315X ], Graduate School of Science and Engineering, Okayama University of Science, Okayama, Japan.
Ryuji Takasaki [ryuji.takasaki@utoronto.ca; ORCID: https://orcid.org/0000-0002-5093-7681 ], Department of Ecology and Evolution, University of Toronto, Toronto, Canada.
Kentaro Chiba [chiba@ous.ac.jp; ORCID: https://orcid.org/0000-0003-2613-6187 ], Mototaka Saneyoshi [saneyoshi@big.ous.ac.jp; ORCID: https://orcid.org/0000-0002-4296-6896 ], Akio Takahashi [takahashi@big.ous.ac.jp; ORCID: https://orcid.org/0000-0002-2766-7775 ], Shoji Hayashi [hayashi@big.ous.ac.jp; ORCID: https://orcid.org/0000-0002-6459-6515 ], Department of Biosphere-Geosphere Science, Okayama University of Science, Okayama, Japan.
Masahito Natori [natori@zool.ous.ac.jp; ORCID: https://orcid.org/0009-0005-6742-5358 ], Department of Zoology, Okayama University of Science, Okayama, Japan.
Shota Kodaira [kodaira_shota@city.nagasaki.lg.jp; ORCID: https://orcid.org/0009-0001-5866-1683 ], Nagasaki City Dinosaur Museum, Nagasaki, Japan.
Shinobu Ishigaki [ishigaki@ous.ac.jp; ORCID: https://orcid.org/0009-0003-1945-6286 ], Museum of Dinosaur Research, Okayama University of Science, Japan.
Buuvei Mainbayar [Mainbayarb@mas.ac.mn; ORCID: https://orcid.org/0000-0003-0608-2871 ] and Khishigjav Tsogtbaatar [tsogtbaatar@mas.ac.mn; ORCID: https://orcid.org/0000-0001-5604-4318 ], Institute of Paleontology, Mongolian Academy of Science, Ulaanbaatar, Mongolia.
Received 29 September 2024, accepted 18 February 2025, published online 31 March 2025.
Copyright © 2025 T. Okoshi et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Upper Cretaceous strata of Mongolia provided over 20 mammalian species, mostly known from the aeolian-dominated Djadokhta and Baruungoyot formations (16 species from the former and seven species from the latter, Lopatin 2020). The mammalian fossils from these aeolian-dominated formations include nearly complete skulls and skeletons (e.g., Kielan-Jaworowska 1970; Novacek et al. 1997; Wible et al. 2007; Hurum and Kielan-Jaworowska 2008). The mammalian faunae in these strata are dominated by multituberculates (Averianov and Archibald 2005; Kielan-Jaworowska et al. 2004) with some metatherians (e.g., Gregory and Simpson 1926; Trofimov and Szalay 1994) and eutherian Asioryctitheria (e.g., Kielan-Jaworowska 1969, 1975; Novacek et al. 1997), Zalambdalestidae (e.g., Gregory and Simpson 1926; Kielan-Jaworowska 1975; Fostowicz-Frelik 2016), and Cimolestidae (e.g., Wible et al. 2007). Mammalian remains from the overlying Nemegt Formation, which is fluvial-dominated and partially contemporaneous with the Baruungoyot Formation, are scarce but represented by two taxa, a fragmentary cranial material of a multituberculate and a relatively well-preserved cranium of a metatherian (e.g., Kielan-Jaworowska and Sochava 1969; Szalay and Trofimov 1996). However, the understanding of mammalian fauna in the Bayanshiree and the Javkhlant formations, the lowermost Upper Cretaceous strata in Mongolia, are much more limited; conference abstracts of unnamed eutherians from the Javkhlant Formation (Giallombardo and Novacek 2006; Giallombardo 2007) and the recent description of a metatherian Tsagandelta dashzevegi Rougier et al., 2015, and an asioryctitherian Bayshinoryctes shuvalovi Lopatin & Averianov, 2023, from the Bayanshiree Formation. Of particular interest in Mongolia is the absence of zhelestids, which are one of the Late Cretaceous eutherian clades (Wible et al. 2007, 2009; Archibald and Averianov 2012), well-known from contemporaneous Cenomanian to Maastrichtian strata across multiple continents: Asia, Europe, North America, and potentially Africa (Wible et al. 2009; Archibald and Averianov 2012). Among these continents, Asia represents the highest taxonomic diversity and the numeric abundance of zhelestids, mostly from central Asian countries such as Uzbekistan and Kazakhstan. Here we describe the first zhelestid specimen from Mongolia, which is also the third mammalian material from the Bayanshiree Formation. This specimen provides new insights into the paleobiogeographical hypothesis of this clade that previous studies provoked (Nessov et al. 1998).
Institutional abbreviations.—MPC, Mongolian Paleontological Center, Mongolian Academy of Sciences, Ulaanbaatar, Mongolia (formerly Institute of Paleontology, Mongolian Academy of Science).
Nomenclatural acts.—This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:CC5A9DA0-2082-410D-9DA0-B54744E8197F.
Material and methods
The new eutherian specimen described here, MPC-M 100s/001, was recovered from the Bayanshiree Formation exposed at the Bayan Shiree locality of the Gobi Desert, Mongolia (Fig. 1A). This formation is widely distributed throughout the eastern and southern Gobi (Martinson 1982) and approximately 35 m of this formation is exposed at Bayan Shiree (Fig. 1B). The depositional age of the Bayanshiree Formation had been recognized as Cenomanian to Santonian based on biostratigraphical data and K-Ar dating on basalt (e.g., Jerzykiewicz and Russel 1991; Shuvalov 2000; Averianov and Sues 2012). This age is supported by the recent U-Pb dating recovered 95.9±6.0 and 89.6±4.0 Ma (Cenomanian–Santonian) on caliches recovered from the Khongil Tsav locality, which is 19 km east of the Bayan Shiree locality (Kurumada et al. 2020). The section at Bayan Shiree mainly consists of mudstone and sandstone. Laterally continuous red mudstone layers, each about 5 to 10 m thick, are observed throughout the section. Most of these mudstone layers are directly overlain by sandstones exhibiting fining-upward sequences with well-developed sedimentary structures such as trough cross-stratification, planar cross-stratification, and current ripple lamination. The sandstone layers of the middle to upper part tend to be thicker, laterally continuous and have well-developed sedimentary structures, but those of the lower part are thinner and less laterally continuous with poorly developed sedimentary structures. These lithostratigraphic features suggest that the section was composed of fluvial deposits. The lower part of the sections is likely to be braided river deposits, whereas the middle and upper parts are deposited with the influence of meandering rivers.

Fig 1. Map of the locality of Ravjaa ishiii gen. et sp. nov. in Mongolia. A. Southern Mongolia with the position of the Bayan Shiree locality. B. Combined stratigraphic columns at Bayan Shiree locality. Photograph (C) and stratigraphic column (D) of the layer from which the specimen was derived. The locality is indicated by a specimen silhouette.
In 2019, a member of the joint expedition of the Institute of Paleontology and Geology (currently the Institute of Paleontology) of the Mongolian Academy of Sciences and Okayama University of Science, Japan found a layer with abundant microvertebrate remains (Ishigaki et al. 2019). The fossil remains from the layer include isolated bones of fishes, anurans, lizards, turtles, crocodilians, non-avian dinosaurs, and the mammal described herein. The mammal specimen was surface-collected from near the top of the mound, indicating the specimen is not a float from the higher stratigraphic position (Fig. 1C). The specimen-bearing layer is in an approximately 1-meter thick point bar sandstone of a laterally accretional meandering channel deposit. Although the sedimentological feature of the host facies indicates that the specimen was transported prior to the final burial event, the pristine preservation of the specimen without abrasion suggests that the distance of the transportation was not significant.
We describe MPC-M 100s/001 following Nessov et al. (1998) for dental terminologies unless noted. Molar measurements were taken based on the scheme of Archibald (1982). The comparable morphological data of other zhelestid specimens were compiled from previous literature. We conducted micro X-ray computed tomography (CT) scanning on MPC-M 100s/001 in addition to the gross observation. The scan images were obtained by Nikon XT H 225 ST at the Industrial X-ray/CT Systems Rental Lab, Nikon Solutions CO., LTD., in Yokohama, Kanagawa Prefecture, Japan. The obtained CT images were rendered using VGSTUDIO MAX 3.4 (Volume Graphics, Germany) and segmented using 3DSlicer version 5.3 (Fedorov et al. 2012).
The phylogenetic analyses were performed using TNT 1.5, May 2022 edition “No Taxon Limit” (Goloboff and Catalano 2016), based on the data matrix of Averianov and Archibald (2016), which is composed of a broad taxonomic sampling of Cretaceous eutherian mammals, including zhelestids. We added Valentinella vitrollense Tabuce et al., 2004, Mistralestes arcensis Tabuce et al., 2013, and Azilestes ragei Gheerbrant and Teodori 2021, to this dataset (see SOM 1, Supplementary Online Material available at http://app.pan.pl/SOM/app70-Okoshi_etal_SOM.pdf). The data matrix consists of 95 taxa and 416 characters (see Appendix 1 for scorings of the newly described specimen in this study), including Nanolestes drescherae Martin, 2002, as the outgroup. All of the characters are equally weighed and not ordered. The maximum number of trees was set to 99 999, and then a traditional search with 100 random seeds and 1000 replicates of Wagner trees, followed by the TBR branch swapping that held 100 trees per replicate, was performed. The branch supports were evaluated by bootstrap resampling using standard absolute frequencies (10 000 replicates). The resulting phylogenetic relationships were visualized using R version 4.3.0, employing two packages: APE version 5.8 (Paradis et al. 2004) and strap version 1.6-1 (Bell and Lloyd 2015). The locality map was produced using GMT6 (Wessel et al. 2019).
Systematic palaeontology
Mammalia Linnaeus, 1758
Theria Parker and Haswell, 1897
Eutheria Gill, 1872
Zhelestidae Nessov, 1985a
Genus Ravjaa nov.
Zoobank LSID: urn:lsid:zoobank.org:act:500CED48-4C63-406B-8D17-50FCD17513D3.
Etymology: Derived from the 5th Noyon Khutugt lama of the Gobi, Dulduityn Danzanravjaa, the famous Buddhist scholar who was deeply respected by people in the Dornogobi region, where this specimen was unearthed. Given that the genus name “Ravjaa” originates from the personal name of a male figure, it should be considered masculine.
Type species: Ravjaa ishiii sp. nov.; see below.
Diagnosis.—As for the type species by monotypy.
Stratigraphic and geographic range.—Bayanshiree Formation, Cenomanian–Santonian, Bayan Shiree, Gobi Desert, Mongolia.
Ravjaa ishiii sp. nov.
Figs. 2–4.
Zoobank LSID: urn:lsid:zoobank.org:act:416ECE52-5728-493F-8DAB-2632FF8228D4.
Etymology: In honor of Ken-ichi Ishii, the director of Hayashibara Museum of Natural Sciences, Okayama, Japan, for his leading role in the Japanese team of the Mongolia-Japan joint expedition, which tremendously contributes to Mongolian paleontology.
Holotype: MPC-M 100s/001, partial right dentary with a distal portion of the ultimate premolar, damaged m1, and well-preserved m2–3.
Type locality: Bayan Shiree, located approximately 25 km southwest of Dzüünbayan, Dornogovi, Mongolia.
Type horizon: Cenomanian–Santonian, Upper Cretaceous, Bayanshiree Formation.
Diagnosis.—A zhelestid eutherian that differs from all other zhelestids in: the faint coronoid crest of the dentary with the shallow masseteric fossa, the concave dorsal alveolar margin of the dentary, the complete absence of the labial mandibular foramen, and the significantly high m2 crown that reaches approximately 70% of the height of the horizontal ramus directly below it on labial view.
Ravjaa ishiii sp. nov. differs from Borisodon kara Archibald & Averianov, 2012, in the rounded mesiolingual vertical crest of the paraconid, the coronoid process tilting 110–115°, and the lack of Meckelian groove; differs from Gallolestes spp. (Gallolestes pachymandibularis Lillegraven, 1976, and Gallolestes agujaensis Cifelli, 1994) in the less bulbous, more anteroposteriorly compressed protoconid; differs from Azilestes ragei in the lack of hypolophid; differs from Parazhelestes spp. (Parazhelestes robustus Nessov, 1993, and Parazhelestes mynbulakensis Nessov, 1985b) in the absence of a marked tuberosity or concavity in the medial side of the area of the coronoid facet between the m3 and base of the coronoid process; differs from Borisodon kara and Eoungulatum kudukensis Nessov et al., 1998, by the absence of coronoid facet; differs from Eozhelestes in the complete labial attachment of the cristid oblique to the notch of the protocristid; differs from Zhalmouzia bazhanovi Averianov et al. 2014, in the lack of cingulid of the lingual and labial side of molars; differs from Borisodon kara and Eozhelestes mangit Nessov, 1997, by the trigonid height is less than twice the talonid height; differs from Avitotherium utahensis Cifelli, 1990, and Lainodon orueetxebarriai Gheerbrant & Astibia, 1994, by the slight labial shift of the paraconid; differs from Sheikhdzheilia rezvyii Averianov & Archibald, 2005, and Lainodon orueetxebarriai by presence of the labial postcingulid; differs from Mistralestes arcensis, the entoconid subequal to the hypoconulid; differs from Mistralestes arcensis in the precingulid with a distinct posteroventral cingular shelf; differs from Eozhelestes mangit, Sheikhdzheilia rezvyii, Lainodon orueetxebarriai, Mistralestes arcensis in the transversed protocristid; differs from Zhalmouzia bazhanovi, Mistralestes arcensis, and Valentinella vitrollense in the position of the posterior mental foramen below the ultimate premolar; differs from Eozhelestes mangit, Parazhelestes spp., Aspanlestes aptap Nessov, 1985a, Zhelestes termirkazyk Nessov, 1985a, Eoungulatum kudukensis, and Zhalmouzia bazhanovi by the ultimate molar hypoconulid being tall and recurved; differs from Eozhelestes mangit, Borisodon kara, Parazhelestes spp., Aspanlestes aptap, Zhelestes termirkazyk, Eoungulatum kudukensis, and Zhalmouzia bazhanovi by the absence of the labial mandibular foramen; differs from Eozhelestes mangit, Shikhdzheilia rezvyii, Borisodon kara, Lainodon orueetxebarriai, Aspanlestes aptap, Zhelestes termirkazyk, Eoungulatum kudukensis, and Zhalmouzia bazhanovi by the compressed trigonid angle being less than 35°.
Description.—The new specimen, MPC-M 100s/001, is a partial right mandible retaining roughly the posterior half of the horizontal ramus, the base of the coronoid process, the anteroventral part of the ascending ramus, the talonid of the ultimate premolar, and m1–3 (Fig. 2). MPC-M 100s/001 represents an adult individual based on the presence of the m3 and the absence of replacement tooth, observed in micro CT images (Fig. 3). Despite the adult status, this specimen represents one of the smallest zhelestids reported to date, together with Sheikhdzheilia rezvyii (Averianov and Archibald 2005) and Aspanlestes aptap (Archibald and Averianov 2012). The dorsoventral depth of the horizontal ramus is consistent throughout its preserved region. The alveolar margin of the ramus is slightly concave in the labial and lingual view, unlike the other zhelestids in which the margin is straight throughout its length (Fig. 2A, C). The dorsal margin of the ramus is higher lingually than labially (Fig. 2A2). The ventral margin of the horizontal ramus is convex in labial and lingual views (Fig. 2A, C). Ventral to the ultimate premolar, the labial surface of the mandible bears a shallow sulcus that runs anteroposteriorly (Fig. 2A). The sulcus likely connects to a mental foramen anteriorly, as in Borisodon kara, Parazhelestes spp., and Eoungulatum kudukensis (Nessov et al. 1998; Archibald and Averianov 2012). The coronoid crest emerges as a low eminence, ventrally to the m2 talonid, at the level of the middle of the horizontal ramus (Fig. 2A2). Musculus buccinator is likely to insert anteromedial to the eminence. The crest extends posterodorsally and becomes prominent but is much less developed than the other zhelestids (e.g., Borisodon kara, Aspanlestes aptap, Eoungulatum kudukensis; Archibald and Averianov 2012), which may be suggestive of a reduced M. temporalis on this individual. The posterior surface of the coronoid crest is rugose and lacks the labial mandibular foramen (Fig. 2A2). The coronoid crest defines the dorsal border of the masseteric fossa. The masseteric fossa is shallow, and its dorsal region likely provides the insertion for M. zygomaticomandibularis, whereas its ventral region is likely to be the insertion for M. masseter (Turnbull 1970). Ventral to the masseteric fossa, the ventral margin of the mandible, anterior to the angular process, is gently concave (Fig. 2A). Lingually, the ascending process lacks the coronoid facet, which can be observed in Eoungulatum kudukensis and Parazhelestes spp. (Archibald and Averianov 2012). The Meckelian sulcus is absent, as in all members of the Zhelestinae (Archibald and Averianov 2012). The CT images demonstrate the presence of the mandibular canal that runs anteroposteriorly throughout the mandible and the absence of replacement teeth (Fig. 3). Since no replacement teeth are present, all teeth observed in this specimen are considered to be permanent teeth.
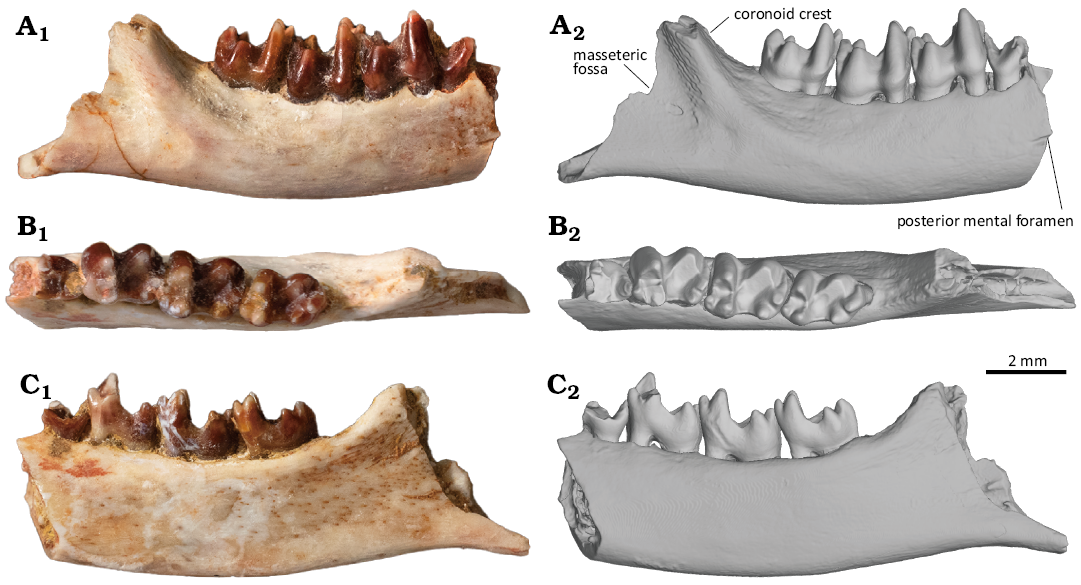
Fig. 2. Holotype of the zhelestid eutherian mammal Ravjaa ishiii gen. et sp. nov. (MPC-M 100s/001) from the Upper Cretaceous Bayanshiree Formation, Dornogovi, Mongolia. Right mandible in labial (A), occlusal (B) and lingual (C) views. Photographs (A1–C1) and 3D model (A2–C2) produced from a CT scan.
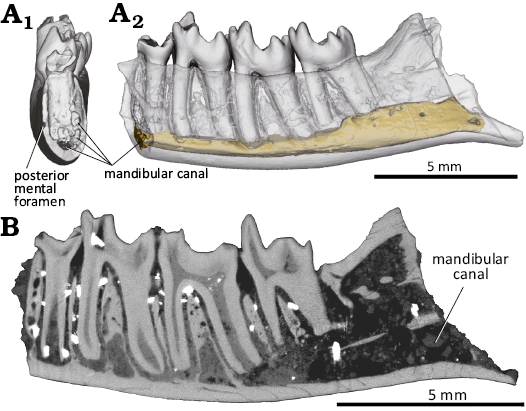
Fig 3. Digital reconstruction and CT image of the zhelestid eutherian mammal Ravjaa ishiii gen. et sp. nov. from the Upper Cretaceous Bayanshiree Formation, Dornogovi, Mongolia A. 3D model in mesial (A1) and lingual (A2) views. B. Longitudinal section of mandible.
MPC-M 100s/001 retains a partial ultimate premolar and all three molars (Fig. 3). While the anterior half of the ultimate premolar is severely damaged, the talonid and the posterior root are well preserved. Despite the damage, the anterior portion of the premolar width is clearly wider than the talonid. The talonid has a narrow and shallow basin anteriorly and has a single cusp posteriorly. All three molars are nearly complete, including two roots per molar, except for the apex of the m1 metaconid. The lower molars m1–3 are tribosphenic molars, and their shapes generally resemble each other. The overall sizes of the molars are subequal to each other, although the m3 is slightly smaller than the other two. The height of the m2 crown is 71% of the height of the horizontal ramus directly below it on labial view, which is markedly larger than in the other zhelestids (50–63%, with 56% on average).
The molars have moderately compressed trigonid with trigonid angles of 33° in m2 and m3 (Fig. 4C1 and D1). The maximum height of the trigonid is higher than that of the associated talonid but does not reach twice the talonid height (Fig. 4B3–D3). Their cusps are rounded as in Parazhelestes spp. and Eoungulatum kudukensis but unlike Borisodon kara. Among the trigonid cusps, paraconid is the smallest and is placed mesiolabially (Fig. 4B1–D1). This is also the only trigonid cusp demonstrating a clear sign of attrition, especially pronounced in the m1. The other two trigonid cusps, the protoconid and the metaconid, are subequal in height (Fig. 4B2–D2 and B4–D4). The protoconid is labially expanded at its base, making it the most robust cusp in the trigonid (Fig. 4B2–D2 and B5–D5). The anteroposterior length of the protoconid is the longest in the m1 and becomes successively thinner in the posterior molars (Fig. 4B1–D1). Compared to the protoconid, the metaconid is slightly less developed and lacks lingual expansion at its base, though it is still more robust than the paraconid (Fig. 4B3–D3 and B4–D4). The paraconid and the metaconid are separated by a prominent gap (Fig. 4B1–D1), which tends to be more prominent in the posterior molars. The trigonid basin is V-shaped in anterior and posterior views (Fig. 4B4–D4). The protocristid is oriented transversely in the m2 and the m3, while it is slightly oblique in the m1 (Fig. 4B1–D1). The talonid width is subequal to the trigonid, and the talonid length accounts for more than half of the total length of the molar. Among the talonid cusps, the hypoconid is the most robust and the shortest cusp (Fig. 4B1–D1 and B5–D5). The attrition surface of the hypoconid is sloped lingually toward the talonid basin (Fig. 4B4–D4). The talonid basin of the m3 bears a fait spherical depression (Fig. 4D1). The hypoconulid and the entoconid are in close approximation to each other (Fig. 4B1–D1), as in all other zhelestids (Archibald and Averianov 2012; Averianov et al. 2014). These two cusps are subequal in size, although the hypoconulids of the m1 and m2 are slightly worn mesiolabially (Fig. 4B1–D1). The hypoconulid of the m3 is more massive than that of the m1 and the m2 and markedly procurved (Fig. 4D3 and D5). The cristid obliqua emerges from the labial margin of the hypoconid and attaches slightly labially to the notch of the protocristid (Fig. 4B1–D1). The postcristid is distinctly present between the hypoconid and the hypoconulid, while it is absent between the hypoconulid and the entoconid. The precingulid is present anterior to the trigonid in all molars and is more prominent in the posterior molars. The labial and lingual margins of the molars are smooth and lack a defined cingulid as in the other zhelestids except Zhalmouzia bazhanovi, in which the molar exhibits faint labial and lingual cingulids (Averianov et al. 2014). A prominent labial postcingulid is present posterolabial to the talonid and bears a cusp-like tubercle protruding from the hypoconulid base; this structure might be comparable to the hypocingulid, the term coined in Van Valen (1993).
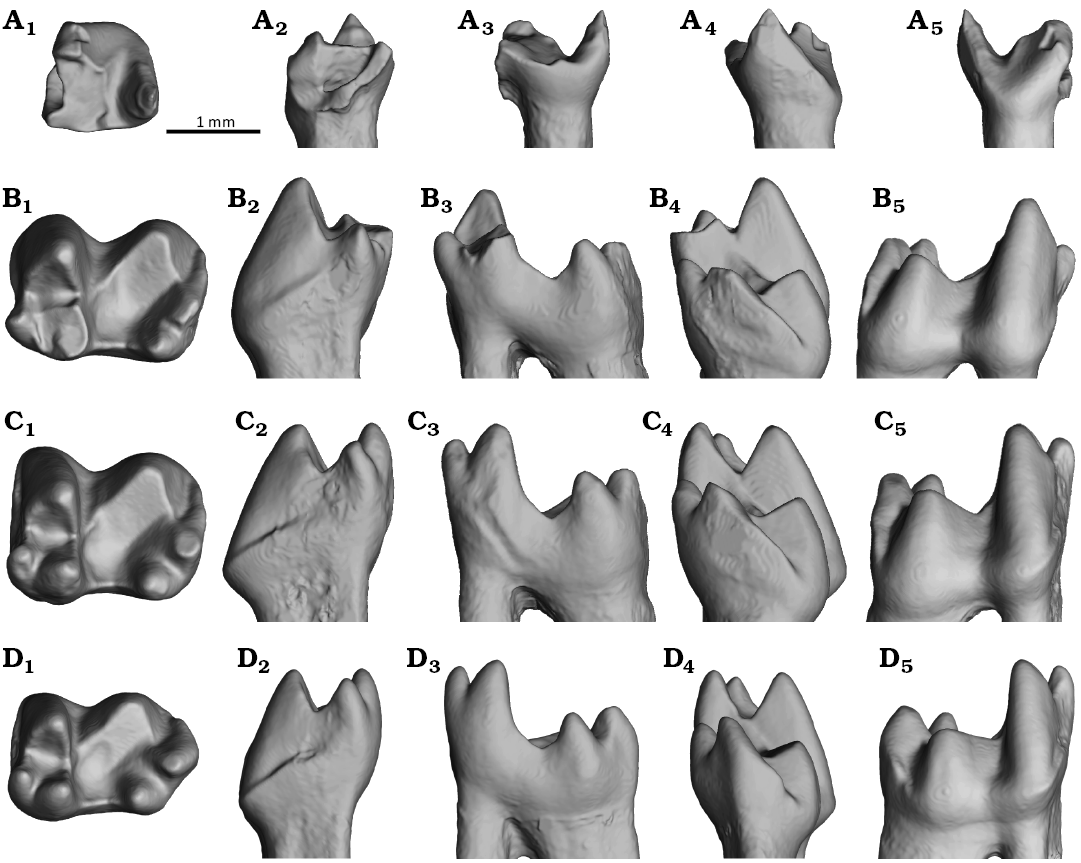
Fig 4. 3D model of ultimate premolar (A) and molars (B, m1; C, m2; D, m3) of the zhelestid eutherian mammal Ravjaa ishiii gen. et sp. nov. from the Upper Cretaceous Bayanshiree Formation, Dornogovi, Mongolia. Tooth in occlusal (A1–D1), mesial (A2–D2), lingual (A3–D3), distal (A4–D4), and labial (A5–D5) views.
Measurements (in mm).—m1: length 2.05; trigonid length 0.96, width 1.53; talonid length 1.09, width 1.55. m2: length 2.17; trigonid length 0.84, width 1.58; talonid length 1.33, width 1.59; distance between paraconid and metaconid 0.190. m3: length 2.09; trigonid length 0.76, width 1.43; talonid length 1.33, width 1.37; distance between paraconid and metaconid 0.19.
Stratigraphic and geographic range.—Type horizon and locality only.
Phylogenetic analysis
We conducted cladistic analyses to demonstrate the phylogenetic position of the new taxon within Zhelestidae (Fig. 5). According to Wible et al. (2009), which defined Zhelestidae as the clade formed by Sheikhdzheilia rezvyii, Zhelestes termirkazyk, and all their descendants, we treated this clade as the smallest clade containing Sheikhdzheilia rezvyii and Zhelestes termirkazyk. Additionally, Zhelestinae was treated as the smallest clade containing Aspanlestes aptap, Parazhelestes spp., and Eoungulatum kudukensis, according to Archibald and Averianov (2012), which listed these three genera as the included taxa of Zhelestinae. The phylogenetic analyses resulted in 64 of the most parsimonious trees (MPTs) with 2784 steps, each with a consistency index of 0.219 and a retention index of 0.570. The strict consensus tree is poorly resolved (SOM 2); early-branching eutherians such as zhelestids and some asioryctids are polyphyletic, as in the strict consensus trees of Tabuce et al. (2013) and Gheerbrant and Teodori (2021). The 50% majority-rule consensus tree demonstrates a better-resolved relationship, recovering the new taxon as the sister taxon of Zhelestinae within Zhelestidae (Fig. 5; see SOM 3 for the full majority consensus tree). Paranyctoides sternbergi Fox, 1979, and Paranyctoides quadrans Nessov, 1982, are recovered as a sister clade of Zhelestidae as in most recent works (e.g., Archibald and Averianov 2012; Gheerbrant and Teodori 2021) other than Averianov and Archibald (2016) in which Paranyctoides spp. was nested within Zhelestidae. It should be noted that two taxa previously reported as possible zhelestids, Mistralestes arcensis (Tabuce et al. 2013) and Azilestes ragei (Gheerbrant and Teodori 2021), are not recovered as zhelestids in the present analysis. Other than these differences, the topology within the Zhelestidae is congruent with the recent works (e.g., Archibald and Averianov 2012; Averianov et al. 2014). In our phylogenetic analyses, Zhelestidae is supported by six unambiguous synapomorphies, and the close approximation of the hypoconulid to the entoconid (120:3) is scored for the new taxon among the synapomorphies. The clade [Zhelestinae + Ravjaa ishiii] is supported by a single synapomorphy, absence of the “coronoid” facet (156:1), and Ravjaa ishiii sp. nov. has a single autapomorphy, the absence of the labial mandibular foramen (139:0), which is broadly seen among placental mammals. Eozhelestes mangit has been suggested to be a non-zheletine zhelestid in Averianov et al. (2014) but recovered as a member of Zhelestinae in our analysis. Zhelestinae is supported by two shared synapomorphies: the ultimate lower premolar talonid as wide as the anterior portion (57:0) and the short and erect hypoconulid of the ultimate molar (121:0).
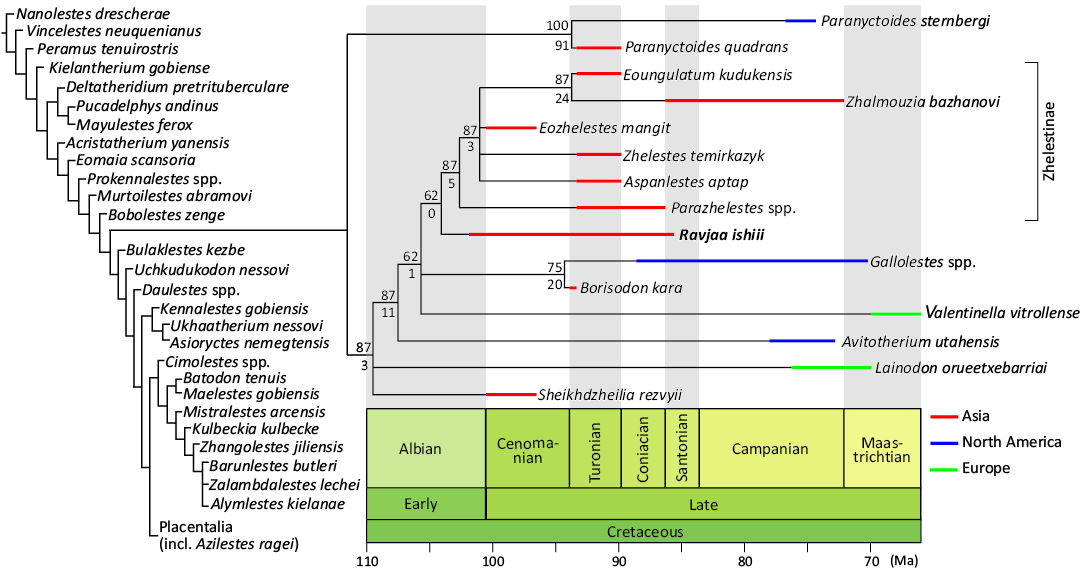
Fig 5. 50% majority-rule consensus tree of the phylogenetic analysis in this study. Numbers on nodes indicate the percentage of MPTs that contain a particular clade (above) and bootstrap supports (below). The figure provides a visualization of the fossil stratigraphic range within the Zhelestidae + Paranyctoides clade, with branching ages and branch lengths shown schematically. For further details see SOM 4.
Discussion
Our phylogenetic analyses recovered the new specimen within the Zhelestidae as the single sister taxon of the Zhelestinae (Fig. 5). Although the branch supports of the recovered tree are generally weak, the topology is well resolved compared to those of recent studies focused on zhelestid phylogeny (Tabuce et al. 2013; Gheerbrant and Teodori 2021), and attribution of the new specimen within Zhelestidae is supported by possession of a zhelestid synapomorphy (120:3). Among Zhelestidae, Ravjaa ishiii sp. nov. is unique in the large molars relative to the horizontal ramus of the dentary, the concave dorsal margin of the dentary, the absence of the labial mandibular foramen posterior to the coronoid crest of the dentary, and the undeveloped coronoid crest of the ascending ramus of the dentary. Together with the phylogenetic autapomorphy (139:0), the data provided here demonstrate that new specimens cannot be assigned to any known taxon and represent the new taxon within Zhelestidae.
The new taxon, Ravjaa ishiii, presented herein, is the first zhelestid from the Bayanshiree Formation and in fact the first discovered in Mongolia. The Upper Cretaceous strata in the Gobi Desert of Mongolia are known for extraordinarily well-preserved mammalian fossils and contribute to understanding the evolutionary history of the Asian Mesozoic mammals (Kielan-Jaworowska et al. 2000), but they have been mainly recovered from the overlying aeolian Djadokhta and Baruungoyot formations (Shuvalov 2000). Up to this study, a single dentary of a metatherian mammal Tsagandelta dashzevegi (Rougier et al. 2015) and a fragmentary skull of a eutherian mammal Bayshinoryctes shuvalovi (Lopatin and Averianov 2023) are the only known mammalian specimens from the Bayanshiree Formation. The occurrence of this new zhelestid further suggests the higher mammalian diversity in the Bayanshiree Formation than previously known.
The new occurrence of zhelestid reported here may suggest the environmental preference of this clade and/or ecological competition with the multituberculates as proposed in Nessov et al. (1998). Although only three mammalian taxa are known so far from the Bayanshiree Formation, none of the mammalian taxa from the Bayanshiree Formation is a multituberculate, which dominates the mammalian fauna in the overlying Djadokhta and Baruungoyot formations (Kielan-Jaworowska and Nessov 1992). The lack of zhelestids in the Djadokhta and Baruungoyot formations is in striking contrast with the mammalian fauna of southwestern Asia, which is characterized by abundant zhelestids and rare multituberculates (Kielan-Jaworowska 1992). The localities of southwestern Asia, dominated by zhelestids, are mostly represented by the Bissekty and Aitym formations of Uzbekistan, deposited in a semi-humid coastal plain and marginal marine settings (Nessov et al. 1998; Redman and Leighton 2009). Contrarily, the multituberculate-dominated Djadokhta and Baruungoyot formations are interpreted to have been deposited under semi-arid to arid conditions in continental inland (Jerzykiewicz et al. 1993; Eberth 2018). The paleoenvironmental condition of the Bayanshiree Formation has been generally considered to be less arid than the aeolian-dominated Baruungoyot and Djadokhta formations (Jerzykiewicz and Russell 1991; Jerzykiewicz et al. 1993). This paleoenvironmental interpretation can be supported by the fluvial-dominated sequence described in this study, indicating the Bayan Shiree locality was generally in close proximity to a water resource compared to the aeolian-dominated Djadokhta and Baruungoyot localities. Thus, the new occurrence of zhelestid in the Bayanshiree Formation suggests that the paleoenvironmental condition allowed zhelestid to extend their habitat to the continental inland area.
Although the occurrence of zhelestid in the Bayanshiree Formation might support the habitat preference hypothesis of this clade, the anatomical feature of Ravjaa ishiii sp. nov. could also support the ecological competition hypothesis between zheletids and multituberculates. The features such as the moderately compressed trigonid, the short protoconid, and more bulbous cusps seen on Ravjaa ishiii sp. nov. and most of the other zhelestids have been interpreted as reflecting more herbivorous tendency (Archibald 1996; Rose 2006) relative to the majority of Cretaceous therian mammals, which are generally considered to be insectivores with the developed trigonid, the tall protoconid, and the sharp cusps (e.g., Kielan-Jaworowska et al. 2004; Martin et al. 2020; Popowics and Mulimani 2023). This interpretation is further supported by morphometric analysis of the lower molar in early therian mammals from the K/Pg boundary, which suggests that zhelestid molar morphology reflects a frugivore/omnivore region, and it is noted that potentially contributes to increased functional diversity in the early Late Cretaceous (Grossnickle and Newham 2016). Since the majority of multituberculates have been generally thought to be herbivores (e.g., Wilson et al. 2012; Xu et al. 2015; Yuan et al. 2013), the inferred herbivorous tendency in Ravjaa ishiii sp. nov. suggests this taxon could have been an ecological competitor of multituberculates.
Ravjaa ishiii sp. nov. potentially represents the oldest member among zhelestids or as old as the currently known oldest zhelestids collected in Uzbekistan (Fig. 5), suggesting the emergence of this clade occurred around the Early/Late Cretaceous boundary to early Late Cretaceous. This period coincides with the early stage of the Cretaceous Terrestrial Revolution (KTR), a macroevolutionary event in the rapid diversification of various terrestrial animals in response to the radiation of angiosperms (e.g., Lloyd et al. 2008; Benton et al. 2022; Weaver et al. 2024). Through this event, angiosperms have been suggested to provide nutritious dietary resources to terrestrial ecosystems as leaves, flowers, fruits, and nuts (Weaver et al. 2024). The occurrence of potential angiosperm megafossils (Khand et al. 2000), including fruit fossils (Ksepka and Norell 2006) from a Bayanshirenian adjacent locality, suggests nutritious angiosperms indeed comprised the ecosystem that Ravjaa ishiii sp. nov. inhabited. Thus, the inferred herbivorous tendency in Ravjaa ishiii sp. nov. and zhelestids might have been an adaptation to an increase of angiosperms in the terrestrial ecosystem.
Conclusions
This study reports the new zhelestid specimen from the Bayanshiree Formation at Bayan Shiree. The zhelestid affinity of this species is supported by the subequally tall protoconid and metaconid, closely approximated hypoconulid and entoconid. Furthermore, this specimen shows unique characters among zhelestids, such as the faint coronoid crest, the concave dorsal alveolar margin, and the complete absence of the labial mandibular foramen. Thus, we erect a new taxon, Ravjaa ishiii based on the specimen. This species represents the first zhelestid from the Mongolian Late Cretaceous strata, suggesting a more prosperous mammalian diversity of the Bayanshiree formation than previously known and bringing potential paleobiogeographic and ecological insights of zhelestids. Zhelestids, including Ravjaa ishiii sp. nov., preferred relatively humid conditions, had an herbivorous tendency, and the evolution of this clade might have been tied to the KTR. These insights, however, should be thoroughly tested in the future, especially with more discoveries of mammalian fossils of the Mongolian Late Cretaceous strata, updated sedimentological data with the paleoenvironmental reconstruction of the Bayanshiree Formation, which is yet to be documented in detail, and thorough dietary reconstructions as performed in recent studies (Grossnickle and Newham 2016; Grossnickle et al. 2021).
Acknowledgements
We would like to thank Sanjaadas Ulziitseren (Institute of Paleontology, Mongolian Academy of Science, Ulaanbaatar, Mongolia) for specimen access. We gratefully acknowledge Kirstin S. Brink (Department of Earth Sciences, University of Manitoba, Winnipeg, Manitoba, Canada) for her editorial support. We also thank Yu Okuda (Department of Veterinary Medicine, Okayama University of Science, Imabari, Ehime, Japan) for the discussion. We sincerely appreciate Toshimasa Maruyama (Industrial X-ray/CT Systems Rental Lab, Nikon Solutions CO., LTD., Yokohama, Kanagawa, Japan) for their invaluable assistance in conducting the CT scanning, which greatly contributed to our study. We thank the Willi Henning Society for access to TNT. We are also deeply grateful to David Archibald (Department of Biology, San Diego State University, San Diego, USA) and Elis Newham (Faculty of Engineering and the Environment, University of Southampton, Southampton, UK) for their insightful comments and constructive suggestions, which greatly helped improve the quality of our manuscript. Last but not least, we would like to thank the crew of the Mongolian Institute of Paleontology-Okayama University of Science Joint Expedition in 2019 for the collection of the specimen. Additionally, this study was supported by the Private University Research Branding Project (2016–2019, Okayama University of Science) and grants in aid from the Japanese Society for the Promotion of Science (JPJSBP120219941, 24K00162).
Editor: Eli Amson.
References
Archibald, J.D. 1982. A study of Mammalia and geology across the Cretaceous–Tertiary boundary in Garfield County, Montana. University of California Publications in Geological Sciences 122: 1–286.
Archibald, J.D. 1996. Fossil evidence for a Late Cretaceous origin of “hoofed” Mammals. Science 272: 1150–1153. Crossref
Archibald, J.D. and Averianov, A.O. 2012. Phylogenetic analysis, taxonomic revision, and dental ontogeny of the Cretaceous Zhelestidae (Mammalia: Eutheria). Zoological Journal of the Linnean Society 164: 361–426. Crossref
Averianov, A.O. and Archibald, J.D. 2005. Mammals from the mid-Cretaceous Khodzhakul Formation, Kyzylkum Desert, Uzbekistan. Cretaceous Research 26: 593–608. Crossref
Averianov, A.O. and Archibald, J.D. 2016. New evidence on the stem placental mammal Paranyctoides from the Upper Cretaceous of Uzbekistan. Palaeontologica Polonica 67: 25–33.
Averianov, A.O. and Sues, H.D. 2012. Correlation of Late Cretaceous continental vertebrate assemblages in Middle and Central Asia. Journal of Stratigraphy 36: 462–485.
Averianov, A.O., Archibald, J.D., and Dyke, G.J. 2014. A new eutherian mammal from the Late Cretaceous of Kazakhstan. Acta Palaeontologica Polonica 59: 537–542. Crossref
Bell, M.A. and Lloyd, G.T. 2015. Strap: an R package for plotting phylogenies against stratigraphy and assessing their stratigraphic congruence. Palaeontology 58: 379–389. Crossref
Benton, M.J., Wilf, P., and Sauquet, H. 2022. The Angiosperm Terrestrial Revolution and the origins of modern biodiversity. New Phytologist 233: 2017–2035. Crossref
Cifelli, R.L. 1990. Cretaceous mammals of southern Utah. IV. Eutherian mammals from the Wahweap (Aquilan) and Kaiparowits (Judithian) Formations. Journal of Vertebrate Paleontology 10: 346–360. Crossref
Cifelli, R.L. 1994. Therian mammals of the Terlingua local fauna (Judithian), Aguja Formation, Big Bend of the Rio Grande, Texas. Rocky Mountain Geology 30: 117–136.
Eberth, D.A. 2018. Stratigraphy and paleoenvironmental evolution of the dinosaur-rich Baruungoyot-Nemegt succession (Upper Cretaceous), Nemegt Basin, southern Mongolia. Palaeogeography, Palaeoclimatology, Palaeoecology 494: 29–50. Crossref
Fedorov, A., Beichel, R., Kalpathy-Cramer, J., Finet, J., Fillion-Robin, J.C., Pujol, S., Bauer, C., Jennings, D., Fennessy, F., Sonka, M., Buatti, J., Aylward, S., Miller, J.V., Pieper, S., and Kikinis, R. 2012. 3D Slicer as an image computing platform for the Quantitative Imaging Network. Magnetic Resonance Imaging 30: 1323–1341. Crossref
Fostowicz-Frelik, Ł. 2016. A new Zalambdalestid (Eutheria) from the Late Cretaceous of Mongolia and its implications for the origin of Glires. Palaeontologia Polonica 67: 127–136.
Fox, R.C. 1979. Mammals from the Upper Cretaceous Oldman Formation, Alberta. III. Eutheria. Canadian Journal of Earth Sciences 16: 114–125. Crossref
Gheerbrant, E. and Astibia, H. 1994. Un nouveau mammifère du Maastrichtien de Laño (Pays Basque espagnol). Comptes Rendus de l’Académie des Sciences, Paris, Série II, 318: 1125–1131.
Gheerbrant, E. and Teodori, D. 2021. An enigmatic specialized new eutherian mammal from the Late Cretaceous of Western Europe (Northern Pyrenees). Comptes Rendus Palevol 20: 207–223. Crossref
Giallombardo, A. 2007. New Cretaceous eutherian from the Gobi Desert. 67th Annual Meeting of the Society of Vertebrate Paleontology, Austin. Abstract Book, Journal of Vertebrate Paleontology (Supplement): 81A [available online, https://vertpaleo.org/wp-content/uploads/2021/03/JVPabstracts2007.pdf].
Giallombardo, A. and Novacek, M.J. 2006. A new eutherian mammal from the Cretaceous of Mongolia. 66th Annual Meeting of the Society of Vertebrate Paleontology, Ottawa. Abstract Book, Journal of Vertebrate Paleontology (Supplement): 67A [available online, https://vertpaleo.org/wp-content/uploads/2021/04/ABSTRACTS-OF-PAPERS_2006.pdf].
Gill, T. 1872. Arrangements of the families of mammals and synoptical table of the characters of the subdivisions of mammals. Smithsonian Miscellaneous Collections 11: 1–98.
Goloboff, P.A. and Catalano, S.A. 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32: 221–238. Crossref
Gregory, W.K. and Simpson, G.G. 1926. Cretaceous mammal skulls from Mongolia. American Museum Novitates 225: 1–20.
Grossnickle, D.M. and Newham, E. 2016. Therian mammals experience an ecomorphological radiation during the Late Cretaceous and selective extinction at the K-Pg boundary. Proceedings of the Royal Society B: Biological Sciences 283: 20160256. Crossref
Grossnickle, D.M., Weaver, L.N., Jäger, K.R.K., and Schultz, J.A. 2021. The evolution of anteriorly directed molar occlusion in mammals. Zoological Journal of the Linnean Society 194: 349–365. Crossref
Hurum, J.H. and Kielan-Jaworowska, Z. 2008. Postcranial skeleton of a Cretaceous multituberculate mammal Catopsbaatar. Acta Palaeontologica Polonica 53: 545–566. Crossref
Ishigaki, S., Tsogtbaatar, K., Tsujigiwa, H., Mainbayar, B., Takahashi, A., Aoki, K., Shogo, A., Buyantegsh, B., Byambaa, P., Bayardorj, C., Otgonbat, B., Saneyoshi, M., Hayashi, S., and Chiba, K. 2019. Report of the Okayama University of Science: Mongolian Institute of Paleontology Joint Expedition in 2019. The Bulletin of Research Institute of Frontier Science and Technology 1: 53–57.
Jerzykiewicz, T. and Russell, D.A. 1991. Late Mesozoic stratigraphy and vertebrates of the Gobi Basin. Cretaceous Research 12: 345–377. Crossref
Jerzykiewicz, T., Currie, P.J., Eberth, D.A., Johnston, P.A, Koster, E.H., and Zheng, J.J. 1993. Djadokhta Formation correlative strata in Chinese Inner Mongolia: an overview of the stratigraphy, sedimentary geology, and paleontology and comparisons with the type locality in the pre-Altai Gobi. Canadian Journal of Earth Sciences 30: 2180–2195. Crossref
Khand, Y., Badamgarav, D., Ariunchimeg, Y., and Barsbold, R. 2000. Cretaceous system in Mongolia and its depositional environments. Developments in Palaeontology and Stratigraphy 17: 49–79. Crossref
Kielan-Jaworowska, Z. 1969. Preliminary data on the Upper Cretaceous eutherian mammals from Bayn Dzak, Gobi desert. Palaeontologia Polonica 19: 171–191.
Kielan-Jaworowska, Z. 1970. New Upper Cretaceous multituberculate genera from Bayn Dzak, Gobi Desert. Palaeontologia Polonica 21: 35–49.
Kielan-Jaworowska, Z. 1975. Preliminary description of two new eutherian genera from the Late Cretaceous of Mongolia. In: Z. Kielan-Jaworowska (ed.), Results of the Polish-Mongolian Palaeontological Expeditions. Part Ⅳ. Palaeontologia Polonica 33: 5–15.
Kielan-Jaworowska, Z. 1992. Interrelationships of Mesozoic mammals. Historical Biology 6: 185–201. Crossref
Kielan-Jaworowska, Z. and Nessov, L.A. 1992. Multituberculate mammals from the Cretaceous of Uzbekistan. Acta Palaeontologica Polonica 37: 1–17.
Kielan-Jaworowska, Z. and Sochava, A.V. 1969. The first multituberculate from the uppermost Cretaceous of the Gobi Desert (Mongolia). Acta Palaeontologica Polonica 14: 355–371.
Kielan-Jaworowska, Z., Cifelli, R.C., and Luo, Z.-X. 2004. Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure. 630 pp. Columbia University Press, New York. Crossref
Kielan-Jaworowska, Z., Novacek, M.J., Trofimov, B.A., and Dashzeveg, D. 2000. Mammals from the Mesozoic of Mongolia. In: M.J. Benton, M.A. Shishkin, D.M. Unwin, and E.N. Kurochkin (eds.), The Age of Dinosaurs in Russia and Mongolia, 573–626. Cambridge University Press, Cambridge.
Ksepka, D.T. and Norell, M.A. 2006. Erketu ellisoni, a long-necked sauropod from Bor Guvé (Dornogov Aimag, Mongolia). American Museum Novitates 3508: 1–16. Crossref
Kurumada, Y., Aoki, S., Aoki, K., Kato, D., Saneyoshi, M., Tsogtbaatar, K., Windley, B.F., and Ishigaki, S. 2020. Calcite U-Pb age of the Cretaceous vertebrate-bearing Bayn Shire Formation in the Eastern Gobi Desert of Mongolia: Usefulness of caliche for age determination. Terra Nova 32: 246–252. Crossref
Lillegraven, J.A. 1976. A new genus of therian mammal from the Late Cretaceous “El Gallo Formation”, Baja California, Mexico. Journal of Paleontology 50: 437–443
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Vol. 1: Regnum animale. Editio decima, reformata. 824 pp. Laurentii Salvii, Stockholm. Crossref
Lloyd, G.T., Davis, K.E., Pisani, D., Tarver, J.E., Ruta, M., Sakamoto, M., Hone, D.W.E., Jennings, R., and Benton, M.J. 2008. Dinosaurs and the Cretaceous Terrestrial Revolution. Proceedings of the Royal Society B 275: 2483–2490. Crossref
Lopatin, A.V. 2020. A review of the Mesozoic and Cenozoic mammals of Mongolia. Paleontological Journal 54: 779–808. Crossref
Lopatin, A.V. and Averianov, A.O. 2023. A new eutherian mammal from the Upper Cretaceous Bayinshire Formation of Mongolia. Journal of Vertebrate Paleontology 43 (2): e2281478. Crossref
Martin, T. 2002. New stem-lineage representatives of Zatheria (Mammalia) from the Late Jurassic of Portugal. Journal of Vertebrate Paleontology 22: 332–348. Crossref
Martin, T. and Jäger, K.R.K., Plogschties, T., Schwermann, A.H., Brinkkötter, J.J., and Schultz, J.A. 2020. Molar diversity and functional adaptations in Mesozoic mammals. In: T. Martin and W.V. Koenigswald (eds.), Mammalian Teeth Form and Function, 187–214. Dr. Friedrich Pfeil, München.
Martinson, G.G. 1982. The Upper Cretaceous mollusks of Mongolia [in Russian]. In: L.A. Nevesskaya (ed.), Sovmestnaâ Sovetsko-Mongol’skaâ Paleontologičeskaâ Ekspediciâ 17: 5–76.
Nessov, L. A. 1982. The most ancient mammals of the USSR [in Russian]. Ežegodnik Vsesoûznogo Paleontologičeskogo Obŝestva 25: 228–242.
Nessov, L.A. 1985a. New mammals from the Cretaceous of Kyzylkum [in Russian]. Vestnik Leningradskogo Universiteta, Seriâ 7 17: 8–18.
Nessov, L.A. 1985b. Rare bony fishes, terrestrial lizards and mammals from the zone of estuaries and coastal plains ofthe Cretaceous of Kizylkum [in Russian]. Ežegodnik Vsesoûznogo Paleontologičeskogo Obŝestva 28: 199–219.
Nessov, L. A. 1993. New Mesozoic mammals of Middle Asia and Kazakhstan and comments about evolution of theriofaunas of Cretaceouscoastal plains of Asia [in Rusian]. Trudy Zoologičeskogo Instituta RAN 249: 105–133.
Nessov, L.A. 1997. Melovye nemorskie pozvonočnye Severnoj Evrazii (Posthumous edition by L.B. Golovneva and A.O. Averianov). 218 pp. Izdatel’stvo Sankt-Peterburgskogo Universiteta, St. Petersburg.
Nessov, L.A., Archibald, J.D., and Kielan-Jaworowska, Z. 1998. Ungulate-like mammals from the Late Cretaceous of Uzbekistan and a phylogenetic analysis of Ungulatomorpha. Bulletin of Carnegie Museum of Natural History 34: 40–88.
Novacek, M.J., Rougier, G.W., Wible, J.R., McKenna, M.C., Dashzeveg, D., and Horovitz, I. 1997. Epipubic bones in eutherian mammals from the Late Cretaceous of Mongolia. Nature 389: 483–486. Crossref
Paradis, E., Claude, J., and Strimmer, K. 2004. APE: analyses of phylogenetics and evolution in R language. Bioinformatics 20: 289–290. Crossref
Parker, T.J. and Haswell, W.A. 1897. A Text-book of Zoology, Volume 2. 683 pp. MacMillan and Company, London. Crossref
Popowics, T. and Mulimani, P. 2023. Mammalian dental diversity: an evolutionary template for regenerative dentistry. Frontiers in Dental Medicine 4: 1158482. Crossref
Redman, C.M. and Leighton, L.R. 2009. Multivariate faunal analyses of the Turonian Bissekty Formation. Palaios 24: 18–26. Crossref
Rose, K.D. 2006. The Beginning of the Age of Mammals. 428 pp. The Johns Hopkins University Press, Baltimore. Crossref
Rougier, G.W., Davis, B.M., and Novacek, M.J. 2015. A deltatheroidan mammal from the Upper Cretaceous Baynshiree Formation, eastern Mongolia. Cretaceous Research 52: 167–177. Crossref
Shuvalov, V.F. 2000. The Cretaceous stratigraphy and palaeobiogeography of Mongolia. In: M.J. Benton, M.A. Shishkin, D.M. Unwin, and E.N. Kurochkin (eds.), The Age of Dinosaurs in Russia and Mongolia, 256–278. Cambridge University Press, Cambridge.
Szalay, F.S. and Trofimov, B.A. 1996. The Mongolian Late Cretaceous Asiatherium, and the early phylogeny and paleobiogeography of Metatheria. Journal of Vertebrate Paleontology 16: 474–509. Crossref
Tabuce, R., Tortosa, T., Vianey-Liaud, M., Garcia, G., Lebrun, R., Godefroit, P., Dutour, Y., Berton, S., Valentin, X., and Cheylan, G. 2013. New eutherian mammals from the Late Cretaceous of Aix-en-Provence Basin, south-eastern France. Zoological Journal of the Linnean Society 169: 653–672. Crossref
Tabuce, R., Vianey-Liaud, M., and Garcia, G. 2004. A eutherian mammal in the latest Cretaceous of Vitrolles, southern France. Acta Palaeontologica Polonica 49: 347–356.
Trofimov, B.A. and Szalay, F.S. 1994. New Cretaceous marsupial from Mongolia and the early radiation of Metatheria. Proceedings of the National Academy of Sciences USA 91: 12569–12573. Crossref
Turnbull, W.D. 1970. Mammalian masticatory apparatus. Fieldiana Geology 18: 147–356.
Van Valen, L.M. 1993. Serial homology: The crests and cusps of mammalian teeth. Acta Palaeontologica Polonica 38: 145–158.
Weaver, L.N., Kelson, J.R., Holder, R.M., Niemi, N.A., and Badgley, C. 2024. On the role of tectonics in stimulating the Cretaceous diversification of mammals. Earth-Science Reviews 248: 104630. Crossref
Wessel, P., Luis, J.F., Scharroo, R., Wobbe, F., Smith, W.H.F., and Tian, D. 2019. The generic mapping tools version 6. Geochemistry, Geophysics, Geosystems 20: 5556–5564. Crossref
Wible, J.R., Rougier, G.W., Novacek, M.J., and Asher, R.J. 2007. Cretaceous eutherians and Laurasian origin for placental mammals near the K/T boundary. Nature 447: 1003–1006. Crossref
Wible, J.R., Rougier, G.W., Novacek, M.J., and Asher, R.J. 2009. The eutherian mammal Maelestes gobiensis from the Late Cretaceous of Mongolia and the phylogeny of Cretaceous Eutheria. Bulletin of the American Museum of Natural History 327: 1–123. Crossref
Wilson, G.P., Evans, A.R., Corfe, I.J., Smits, P.D., Fortelius, M., and Jernvall, J. 2012. Adaptive radiation of multituberculate mammals before the extinction of dinosaurs. Nature 483: 457–460. Crossref
Xu, L., Zhang, X., Pu, H., Jia, S., Zhang, J., Lü, J., and Meng, J. 2015. Largest known Mesozoic multituberculate from Eurasia and implications for multituberculate evolution and biology. Scientific Reports 5: 14950. Crossref
Yuan, C.-X., Ji, Q., Meng, Q.-J., Tabrum, A.R., and Luo, Z.-X. 2013. Earliest evolution of multituberculate mammals revealed by a new Jurassic fossil. Science 241: 779–783. Crossref
Character scores for Ravjaa ishiii gen. et sp. nov., based on the matrix of Averianov and Archibald (2016).
1(0), 2(0), 57(0), 58(0), 61(1), 62(0), 63(0), 106(0), 107(0), 108(0), 109(0), 110(0), 111(2), 112(1), 113(1), 114(1), 115(1), 116(2), 117(1), 118, (2), 119(2), 120(3), 121(1), 122(2), 123(0), 124(0), 125(0), 126(1), 127(0), 130(2), 131(0), 132(1), 135(1), 136(1), 138(0), 139(0), 154(1), 156(1)
Acta Palaeontol. Pol. 70 (1): 193–203, 2025
https://doi.org/10.4202/app.01213.2024