
A terrestrial snake from the lower Eocene of the mid-Atlantic region (Nanjemoy Formation, Virginia) of North America
ADAM C. PRITCHARD, JACOB A. MCCARTNEY, GEORGIOS L. GEORGALIS, and KRISTER T. SMITH
Pritchard, A.C., McCartney, J.A., Georgalis, G.L., and Smith, K.T. 2025. A terrestrial snake from the lower Eocene of the mid-Atlantic region (Nanjemoy Formation, Virginia) of North America. Acta Palaeontologica Polonica 70 (4): 817–826.
We report on the oldest vertebra of a terrestrial snake from the Paleogene of eastern North America. The nearly complete trunk vertebra was recovered from the Eocene Nanjemoy Formation of Virginia and is referred to Constrictores, the clade including booids and pythonoids, as it bears a relatively broad centrum, high neural spine, and a relatively massive zygosphene compared to most other snake taxa. Although a combination of features of the specimen, including a dorsoventrally tall and transversely narrow neural canal and a relatively high neural spine mostly developed in the posterior half of the neural arch, are distinct from most other described Paleogene Constrictores, we refrain from naming a new taxon based on a single element. The discovery of this early Eocene snake in the north of the Paleogene Atlantic coast strengthens similarities with contemporaneous vertebrate assemblages in western North America, the Paleogene Gulf Coast of North America, and western Europe. It also extends the broad biogeographic range of the rich Paleogene radiation of Constrictores to the Atlantic coast of North America. The specimen also exhibits interesting taphonomic signatures (e.g., eroded outer layers of cortical and articular bone, specific damages of the zygantrum) indicating that it may have been digested prior to fossilization.
Key words: Squamata, Serpentes, Constrictores, Eocene, Paleogene, Virginia, USA.
Adam Pritchard [adam.pritchard@vmnh.virginia.gov; ORCID: https://orcid.org/0000-0001-8722-0011 ], Virginia Museum of Natural History, 21 Starling Avenue, Martinsville, Virginia, USA.
Jacob McCartney [jmccart61@naz.edu; ORCID: https://orcid.org/0000-0003-3661-8518 ], Biology Department, Nazareth University, 4245 East Avenue, Rochester, NY 14618.
Georgios Georgalis [georgalis@isez.pan.krakow.pl; ORCID: https://orcid.org/0000-0001-7759-6146 ], Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Sławkowska 17, 31-016, Kraków, Poland.
Krister Smith [krister.smith@senckenberg.de; ORCID: https://orcid.org/0000-0003-1442-2944 ], Department of Messel Research and Paleobiology, Senckenberg Research Institute and Natural History Museum Frankfurt, Senckenberganlage 25, 60325, Frankfurt am Main, Germany.
Received 19 May 2025, accepted 7 October 2025, published online 19 December 2025.
Copyright © 2025 A. Pritchard et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
During the early Paleogene, the Laurasian continents saw a geographically widespread diversification of Constrictores: the clade of alethinophidian snakes including boas and pythons as defined by Georgalis and Smith (2020; also see Gilmore 1938; Rage 1984; Holman 2000; Scanferla and Smith 2020; Georgalis et al. 2021; Smith and Georgalis 2022; Zaher et al. 2023). Reptile assemblages from western Europe and western North America have produced rich records of constrictors, including a wide range of body sizes and disparate vertebral morphologies (Smith and Georgalis 2022). It is unclear to what extent this broad biogeographic range can be attributed to vicariance or to dispersal from a center of origin, partly due to unclear phylogenetic relationships among extinct Constrictores (Smith and Georgalis 2022) but also to the almost complete absence of terrestrial snake fossils from the Paleogene of eastern North America.
A small number of isolated snake vertebrae referrable to Constrictores have been recognized from the far southeast of North America along the Paleogene Gulf Coast (e.g., Holman 1977, 2001; Holman and Case 1988; Smith and Georgalis 2022). Thus far, the absence of Paleogene terrestrial snake records from more northerly localities on the Atlantic coast (e.g., Weems 1999b; Holman 2001; Smith and Georgalis 2022) leaves open questions about the relationships and distribution of snakes across the northern continents:
Are Constrictores present in Atlantic coastal localities, at the intersection between western North America, the Gulf Coast, and western Europe, in the lower Paleogene, supporting a continuous distribution of the clade across these regions?
Do terrestrial snakes in the northern and/or mid-Atlantic sites show closer taxonomic affinities to known taxa in any of the aforementioned regions?
Here, we describe a new record of Constrictores from the Fisher/Sullivan site in Stafford County, Virginia, USA. Representing a locality dated to 54.5 million years ago from the lower Eocene Potapaco B Member of the Nanjemoy Formation (Weems and Grimsley 1999), the Fisher/Sullivan site is a well-known marine microvertebrate locality (e.g., Kent 1999a, b; Weems 1999b; Hilton and Carpenter 2020; Weems 2020), but it has also produced the best record of terrestrial vertebrates from the Paleogene of the mid-Atlantic (e.g., Olson 1999; Rose 1999; Weems 1999a; Rose et al. 2021; Mayr et al. 2022). The new specimen has implications for the biogeography of Constrictores and the biogeographic relationships of the mid-Atlantic Fisher/Sullivan reptile assemblage to contiguous regions in North America and Europe, particularly when taken into consideration that certain congeneric species inhabited both North America and Europe during the Eocene (xd; Georgalis and Mennecart 2025).
Institutional abbreviations.—CM, Carnegie Museum of Natural History, Pittsburgh, USA; GMH, Geiseltalsammlung at Zentralmagazin Naturwissenschaftlicher Sammlungen, Halle, Germany; MCZ, Museum of Comparative Zoology, Cambridge, USA; NHMUK, Natural History Museum, London, UK; MNHN, Museum national d’Histoire naturelle, Paris, France; SMF-PH, Senckenberg Research Institute, paleoherpetological collections, Frankfurt am Main, Germany; Tü, Universität Tübingen, comparative osteological collection, Tübingen, Germany; UMA, University of Massachusetts Natural History Collections, Amherst, USA; USNM, National Museum of Natural History, Washington, DC, USA; VMNH, Virginia Museum of Natural History, Martinsville, USA; YPM, Yale Peabody Museum, New Haven, USA.
Geological setting
The Fisher/Sullivan site outcrops in Stafford County, Virginia, on a tributary of Muddy Creek, itself a tributary of the Rappahannock River. Weems and Grimsley (1999) extensively reviewed the stratigraphy of the site, reporting that the main bonebed of the Fisher/Sullivan locality occurred in “zone 11” of the Potapaco B Member of the Nanjemoy Formation, dating it to the early Eocene (~54.5 Ma) (most recent stratigraphic section illustrated in Rose et al. 2021: fig. 2). Terrestrial vertebrates reported from the site include the anguimorph Parophisaurus mccloskeyi Weems, 1999b, and a mammalian fauna including an omomyid possibly referrable to the genus Teilhardina (see Rose et al. 2021).
Collecting efforts at the site by palaeontological teams from the 1990s to the present have produced tens of thousands of specimens of vertebrates and plants, providing the most complete window into terrestrial and marine ecosystems from the Eocene of the mid-Atlantic of North America.
Material and methods
VMNH 129332 was collected by palaeontologist Michael Folmer (Stafford County, USA) and donated to VMNH in the summer of 2021.
The photographs presented in Fig. 1 were acquired using a Canon EOS 90D camera. Focal stacking was performed in Helicon Focus v. 8.2.2 (Helicon Soft, Ltd.) to create the final images. The specimen was microCT scanned on a Nikon X-Tek scanner at Virginia Polytechnic Institute (Blacksburg, Virginia) at a resolution of 16.21 microns (kV = 205, uA = 209). 3D surface files of the specimen were generated in Dragonfly 3D v 2025 using the “export to a mesh” function on the segmented vertebra. The raw CT slices and surface files are available on Morphosource as Project 000795639 at https://morphosource.org.
Systematic palaeontology
Serpentes Linnaeus, 1758
Alethinophidia Nopcsa, 1923
Constrictores Oppel, 1811
(sensu Georgalis & Smith
2020)
Constrictores indet.
Figs. 1, 2.
Material.—VMNH 129332, a nearly complete mid-trunk vertebra missing a small part of the ventral margin of the cotyle and the dorsolateral margins of the zygantrum. The tip of the neural spine, the lateral edges of the prezygapophyses and synapophyses, part of the zygantral roof, and the condyle are eroded from the lower Eocene of Fisher/Sullivan site, Virginia, USA.
Description.—VMNH 129332 is a nearly complete vertebra that we identify as belonging to the mid-trunk region (Fig. 1), lacking only the posterodorsal portion of the zygantral roof and the ventralmost margin of the cotyle. The surface of the vertebra is somewhat eroded, with thin layers of cortical bone remaining on the neural arch, lateral surface, paracotylar fossae, and neural canal. The synapophyseal facets and condyle are pitted and irregular, suggestive of heavy erosion as well. This erosion may be due to digestive processes (see below). The neural canal and zygantrum remain filled with compacted sand to support the delicate margins of these cavities.

Fig. 1. Photographs of the terrestrial snake Constrictores indet. (VMNH 129332) from the lower Eocene of Fisher/Sullivan site, Virginia, USA. Mid-trunk vertebra in anterior (A1), posterior (A2), dorsal (A3), ventral (A4), right (A5) and left (A6) lateral views.
CT scan data show that the internal architecture of the bone conforms to the general pattern in snakes described by Houssaye et al. (2013), with a prominent anteroventral cavity in the centrum framed posteriorly by a series of anteroposteriorly oriented trabeculae supporting the condyle. The neural spine is supported internally by thin, dorsoventrally running trabeculae (Fig. 2).
Anterior view (Fig. 1A1): The original shape of the cotyle is slightly obscured, as the ventral margin is cracked off, but the structure was likely slightly wider transversely than dorsoventrally tall. There are deep and ventrally tapering paracotylar fossae that bear paracotylar foramina. The ventromedial border of the fossae alongside the lateral cotylar margin is sharply defined. Dorsomedially, the fossae grade smoothly into the ventrolateral edge of the neural canal.
The eroded ventral margins of the synapophyses are positioned directly lateral to the cotyle, with the synapophyseal facets in line with the ventral cotylar margin. The more weathered left synapophyseal surface extends further ventromedially than the right. Neither side shows a demarcation into dorsal and ventral diapophyseal and parapophyseal regions. Although the synapophyseal articular surfaces are eroded, the structures do not appear to have extended far distally from the lateral margin of the centrum. The neural canal is dorsoventrally taller than transversely wide. At its ventrolateral corners, the canal bears two small lateral sinuses. The dorsoventral height of the neural canal is greater than that of the zygosphene and similar to that of the cotyle.
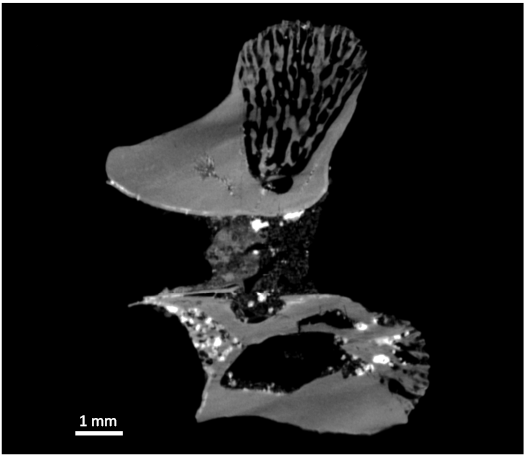
Fig. 2. Median sagittal section of CT scan of vertebra of the terrestrial snake Constrictores indet. (VMNH 129332) from the early Eocene of Fisher/Sullivan site, Virginia, USA.
The prezygapophyses extend farther laterally than the preserved edges of the eroded synapophyses. The prezygapophyseal articular facets are positioned slightly ventral to the level of the dorsoventral midpoint of the neural canal. They are oriented dorsolaterally at an angle between 3–5° from horizontal. No prezygapophyseal accessory processes are preserved, although the cortical bone is eroded at the tips on both sides. If accessory processes were present, they were likely quite small. The better-preserved right prezygapophysis bears a smooth, ventrolaterally inclined margin ventral to the facet. There is a subtle, convex lamina extending from the prezygapophyseal articular facet to the anterodorsal edge of the synapophysis.
The zygosphene is moderately thick, being transversely wider than dorsoventrally tall. The facets are oriented ventrolaterally at roughly 38° relative to the dorsoventral axis. The dorsal edges of the facets are angled dorsolaterally, and the dorsal margin of the zygosphene is convex. The neural spine is dorsoventrally high, forming roughly one-quarter the total dorsoventral height of the vertebra.
Posterior view (Fig. 1A2): The haemal keel is triangular in cross section with a tapered, sharp ventral edge that extends ventral to the level of the condyle. The roughly hemispherical condyle is distinctly eroded, with extensive pitting. The roughly triangular posterior opening of the neural canal is dorsoventrally taller than transversely wide, similar in breadth to the anterior opening and relatively larger than the condyle.
The postzygapophyses are situated at the level of the dorsoventral midpoint of the neural canal. The postzygapophyseal articular facets face ventrolaterally at roughly 80° relative to the dorsoventral axis. The roof of the neural arch dorsal to the postzygapophyseal facets is straight and angled strongly dorsomedially.
The zygantrum is damaged, with prominent portions of the roof broken off at a level just medial to both the left and right postzygapophyseal facets. We estimate the vaulting ratio (sensu Georgalis et al. 2021) at approximately 0.58, although the damage to the zygantral roof obscures the measure.
The posterodorsal margin of the zygantral opening is eroded, with the outer layers of the cortical bone of the dorsal margin being clearly weathered away; however, the overall shape of the roof of the zygantrum appears to remain intact. The zygantral facets are angled strongly dorsomedially at 35° relative to the dorsoventral axis. The opening itself is wider than tall, and its dorsal margin is subtly convex. Although the cavity remains filled with matrix, endozygantral foramina are present based on the CT scan data (see Morphosource Project 000795639, https://doi.org/10.17602/M2/M796201).
Dorsal view (Fig. 1A3): The zygosphene is anteriorly concave, with two lateral lobes and no median lobe. The neural arch is roughly equivalent in maximum anteroposterior length to its maximum transverse width (measured across the postzygapophyses). The prezygapophyses are angled anterolaterally at 48° relative to the midline of the vertebra. The prezygapophyseal articular facets have a wide posteromedial margin and a strongly tapering anterolateral tip (obovate sensu Auffenberg 1963). Both right and left prezygapophyseal facets preserve rings that radiate outwards from their posteromedial edges, with those on the left side being better preserved (probable annulations per Warren 1963, but see Petermann and Gauthier 2018).
The interzygapophyseal constriction is relatively deep near the anteroposterior midpoint of the arch. The neural spine arises as a thin lamina posterior to the zygosphene, becoming prominently developed in the posterior half of the neural arch, overhanging the dorsal margin of the zygantrum. Erosion of the neural spine tip has exposed the interior of trabeculae. However, the bone seems complete enough to support the inference that a transverse expansion of the dorsal tip of the structure was absent. The posterior median notch of the neural arch appears deep, although a thin layer of bone is eroded away in this region.
Ventral view (Fig. 1A4): The haemal keel is broken at its anterior tip. It is transversely narrow throughout its preserved anteroposterior length (the gladiate shape of Auffenberg 1963). The centrum itself is triangular, with the prominent subcentral grooves being bounded by weakly rounded subcentral ridges. A single subcentral foramen is present on the right and the left sides of the centrum, positioned at about one-third of the length of the centrum. The right foramen is twice as broad and positioned slightly farther posteriorly than the left.
The postzygapophyses are strongly posterolaterally angled at 48° relative to the midline. The postzygapophyseal articular facets themselves are also obovate with a strongly tapered posterolateral tip. Each facet bears radiating rings, the probable growth lines, similar to those on the prezygapophyses. The posterolateral tip of the right postzygapophyseal articular facet is broken off, and there is a prominent, subcircular crack running across the surface of the left facet.
Lateral view (Fig. 1A5, A6): The haemal keel is ventrolaterally projected and convex in the posterior half of the centrum. The subcentral grooves are visible throughout the anteroposterior length of the centrum, bounded laterally by rounded subcentral ridges. The synapophyses are roughly reniform in shape (sensu Auffenberg 1963).
The condyle is slightly upturned relative to the long axis of the centrum. The lateral surfaces of both prezygapophyses and postzygapophyses on both sides of the vertebra have the outermost layer of cortical bone eroded at their distal tips. The prezygapophyses extend slightly anteriorly relative to the anterior margin of the zygosphene, whereas the postzygapophyses do not extend posterior to the level of the condyle.
There are lateral foramina on both sides of the vertebra posteroventral to the prezygapophyses and ventral to the well-developed interzygapophyseal ridge. The ridge arcs posterodorsally between the lateral margin of the prezygapophyses and the anterodorsal margin of the postzygapophyses. The left ridge is broken away completely leaving a prominent cleft in the bone.
The zygosphenal facets are well exposed, as is the dorsal convexity of the zygosphene. The posterior margins of the neural arch, dorsal to the postzygapophyses, are flat. The missing dorsolateral edges of the zygantrum are clearly visible in lateral view.
The prominently developed posterior half of the neural spine is roughly rhomboid in shape and angled posterodorsally dorsal to the zygantrum. The dorsal margin of the neural spine is flat with subtly convex anterior and posterior margins. The dorsal tip of the neural arch lacks the outer layer of bone on both sides, as evidenced by dorsally convex cracks.
Results
Comparative anatomy.—We identify the specimen as a mid-trunk vertebra by the presence of deep subcentral grooves, the relatively narrow and sharp haemal keel, the little-projecting synapophyses, and the absence of a hypapophysis. It is unclear if paracotylar notches were present due to damage to the cotyle and synapophyses (sensu La Duke 1991). The specimen is relatively anteroposteriorly shorter and transversely narrower than mid-trunk vertebrae from many contemporaries of VMNH 129332 among Constrictores (linear measurements presented in Table 1). Using the two body length estimations by McCartney et al. (2018), one based on cotylar breadth and the other based on trans-prezygapophyseal width, we estimate the total body length of the snake as between 194.1 and 208.0 cm.
Table 1. Linear measurements (in mm) of key anatomical structures preserved on VMNH 129332. Zygosphene height, measured in from transverse plane between ventral corners of facets; zygosphene width, measured from dorsolateral corners of facets; neural canal height, measured in midline; * indicates an measurement likely obscured by erosion or damage to the specimen.
|
Centrum length |
5.2* |
|
Trans-prezygapophyseal width |
9.1 |
|
Trans-postzygapophyseal width |
9.1 |
|
Neural arch width |
6.0 |
|
Neural spine height |
2.9* |
|
Neural spine length |
2.7 |
|
Cotylar width |
3.8 |
|
Zygosphene height |
1.7 |
|
Zygosphene width |
3.9 |
|
Neural canal height |
2.1 |
Taxonomic identification and referral to Constrictores.—We identify VMNH 129332 as a member of Constrictores based on several anatomical characters. These were recently defined as being present in most Constrictores by Georgalis and Smith (2020) and refined by Szyndlar and Georgalis (2023). These features include:
– The low ratio of centrum length to neural arch width (0.87 in VMNH 129332).
– A high neural spine, in this case occupying one-quarter the dorsoventral height of the vertebra.
– A relatively thick zygosphene, more massive (height/width ratio of 0.40 in VMNH 129332) than is typical for other non-caenophidian snakes, such as scolecophidians, aniliids, cylindrophiids, and uropeltids (see Szyndlar and Georgalis 2023), as well as most caenophidians (e.g., Zaher et al. 2019).
None of these characters are unique synapomorphies (Georgalis and Smith 2020). These features are also present in the non-Constrictores clade Tropidophiidae. However, as a diagnostic feature of tropidophiids is lacking the in Nanjemoy vertebra—the presence of a broad hypapophysis, usually with a distinct anteroventral corner (Smith and Georgalis 2022: appendix 4.S1; Szyndlar and Georgalis 2023; Zaher et al. 2024)—we exclude affinities with Tropidophiidae.
We consider it unlikely that the specimen belongs to Bolyeriidae, Candoiidae, or Ungaliophiidae, as the vertebral morphology of these groups deviates from that of other Constrictores (see Szyndlar and Georgalis 2023: figs. 57–62, 96–102, 118–127). The Nanjemoy specimen lacks the hypapophyses that are persistent throughout the column in bolyeriids or candoiids (Szyndlar and Georgalis 2023) and the relative anteroposterior elongation (Smith 2013) and gracility present in vertebrae of ungaliophiids (see Szyndlar and Georgalis 2023). Below, we compare VMNH 129332 to the mid-trunk vertebrae of Paleogene and extant members of Constrictores, restricting our sample due to the referral of the specimen to this broader clade.
Overall proportions.—The subequal centrum length and maximum neural arch width most resemble the condition in Casarea dussumieri and Exiliboa placata (see Szyndlar and Georgalis 2023: figs. 58, 59, 118–120), contrasting with most other Constrictores that possess greater arch-width/centrum-length ratios (e.g., Sanzinia madagascariensis [YPM R 12241], Boa constrictor [YPM R 12323]). Extinct taxa with similar length-width ratios to VMNH 129332 include Eoconstrictor barnesi (see Palci et al. 2024: fig. 10), Huberophis georgiensis (see Holman 1977), Totlandophis thomasae (see Holman and Harrison 1998a), and Paraplatyspondylia batesi (see Holman and Harrison 1998b). Examples of Paleogene Constrictores with ratios slightly greater than 1.0 include Boavus occidentalis (USNM PAL 427693 and see Onary et al. 2021: fig. 10D), Paleryx rhombifer, and Palaeopython spp. (see Georgalis et al. 2021: table 1), and Eoconstrictor fischeri (see Schaal 2004; Georgalis et al. 2021: table 1).
Centrum.—The morphology of the haemal keel is difficult to directly compare to that of other taxa due to intracolumnar variation, even within the mid-trunk region. In nearly all Constrictores (excluding those extant forms with persistent hypapophyses noted above), the posteroventral portion of the keel is dorsoventrally low and ventrally convex, as in VMNH 129332. The transversely narrow “gladiate” shape of the keel resembles mid-trunk vertebrae in many Paleogene Constrictores, such as Palaeopython spp., Phosphoroboa filholii (e.g., Georgalis et al. 2021: figs. 8–17, 23, 30–32, 79–84), Boavus spp. (Onary et al. 2021: figs. 1D–F, 2), and Cadurcoboa insolita (see Rage 1978: fig 6), as well as extant species of Boa (YPM R 12323; see Szyndlar and Georgalis 2023: figs. 80–85) and Eunectes murinus (CM 145331). Roughly flatter, spatulate keels that broaden posteriorly (sensu Auffenberg 1963) occur in Totlandophis thomasae (see Holman and Harrison 1998a). Relatively broad and flat keels with weak transverse constrictions at the anteroposterior midpoint occur in several enigmatic extinct forms (Huberophis georgiensis, Ogmophis voorhiesi, and Tallahattaophis dunni; see Holman 1977: figs. 2A, 3A; Holman and Case 1988).
The well-defined, dorsally arched subcentral ridges in VMNH 129332 compare favorably to most extant Constrictores, such as Sanzinia madagascariensis and Python sebae (see Szyndlar and Georgalis 2023: figs. 74–78, 146–148). Among fossil taxa, similar well-defined ridges occur in Boavus occidentalis (USNM PAL 437693), Huberophis georgiensis (Holman 1977), Palaeopython spp. (see Georgalis et al. 2021; Smith and Scanferla 2022), and Totlandophis thomasae (Holman and Harrison 1998a). In Huberophis georgiensis and Totlandophis thomasae, the ridges are similarly dorsally arched in lateral view, whereas they are straighter in Morelia viridis, Python spp. (see Szyndlar and Georgalis 2023: figs. 142, 147, 159), and Palaeopython spp. (GMH XXIV-274-1964 and see Georgalis et al. 2021).
Cotyles and condyles.—In VMNH 129332, both the cotylar and condylar shapes are obscured by damage to the margins and heavy erosion. Although the cotyle was transversely wider than tall, it was relatively less dorsoventrally compressed than in scolecophidians (see Szyndlar and Georgalis 2023) and the enigmatic taxon Tallahattaophis dunni (see Holman and Case 1988). The maximum cotylar width can be assessed based on the preserved margins as being equivalent to the breadth of the neural canal and that of the zygosphene. Subequal widths of the cotyle and neural canal occur in Casarea dussumieri (MCZ 49135), Eryx spp. (E. colubrinus SMF-PH 18; E. johnii SMF-PH 20; see Szyndlar and Georgalis 2023); and extinct taxa such as Totlandophis spp. (Holman and Harrison 1998b, 2001), Phosphoroboa filholii (see Georgalis et al. 2021: figs. 48–59, 78–81, 83–86), and Boavus brevis (see Onary et al. 2021: fig. 2). In examples of Palaeopython cadurcensis (see Georgalis et al. 2021: figs. 10–13, 17) and Palaeopython ceciliensis (see Georgalis et al. 2021: figs. 18–20), the ventral margin of the cotyle is subequal in width to the canal, which becomes extremely transversely wide at its ventral extent.
Zygapophyses.—The anterolateral angling of the prezygapophyses in VMNH 129332 of 48° is similar to that in some extant Constrictores, such as Casarea dussumieri (MCZ 49135), and ungaliophiids (see Szyndlar and Georgalis 2023: 118–120, 124, 125). In extinct taxa, similar angles occur in Totlandophis thomasae (see Holman and Harrison 1998b), Huberophis georgiensis (see Holman 1977: fig. 2A), Tallahattaophis dunni (see Holman and Case 1988: fig. 3A), and Boavus brevis (see Onary et al. 2021). A substantially wider angle occurs in species of Palaeopython (see Georgalis et al. 2021), Rageryx schmidi (see Smith and Scanferla 2022: fig. 9), Eoconstrictor fischeri, Phosphoroboa filholii (see Georgalis et al. 2021), Boavus occidentalis (USNM PAL 427693 and see Onary et al. 2021: figs. 10E, G), and Cadurcoboa insolita (see Rage 1978: fig. 6d, v).
The probable presence of extremely small prezygapophyseal accessory processes and a strongly dorsolaterally oriented ventrolateral margin to the prezygapophysis itself is similar to the ungaliophiine Exiliboa placata (see Szyndlar and Georgalis 2023: figs. 120, 121) and the candoiid Candoia carinata (CM 118570, and see Szyndlar and Georgalis 2023: figs. 96–102). A dorsolaterally oriented or ventrally rounded ventral margin to the prezygapophysis occurs in Totlandophis thomasae (see Holman and Harrison 1998b: fig 1A) and Paleryx rhombifer (see Georgalis et al. 2021: figs. 35A, 36A, 38A).
Zygosphene/zygantrum.—Zygosphenes of modest robusticity (height/width ratio about 0.40) in the mid-trunk vertebrae, as in VMNH 129332, is similar to that in extant taxa such as Epicrates cenchria (YPM R 13810), Boa imperator (CM S 9717), Candoia carinata (YPM R 13199), and Python molurus (YPM R 12545). Among extinct taxa, similarly proportioned zygosphenes occur in Boavus occidentalis (USNM PAL 427693), Rageryx schmidi (see Smith and Scanferla 2021: figs. 11B), and Totlandophis thomasae (see Holman and Harrison 1998b: figs. 1A, 2A).
An anteriorly concave zygosphene similar to VMNH 129332 occurs in the extinct Tallahattaophis dunni (see Holman and Case 1988), some vertebrae of Ogmophis compactus (see Smith 2013), and some vertebrae of Boavus occidentalis (see Onary et al. 2021: fig. 10B, F, G), and extant constrictors including Boa spp. (CM S 9717; see Szyndlar and Georgalis 2023: figs. 83, 84), and some Eryx spp. (e.g., E. johnii SMF-PH 20, E. jayakari NHMUK 1909.10.15.8, E. jaculus Tü-VI.1935). A concave zygosphene interrupted by a midline lobe occurs in Boavus brevis (see Onary et al. 2021), other Ogmophis compactus (Smith 2013), Eoconstrictor spp. (see Georgalis et al. 2021), and some extant species of booids and pythonoids. A similar dorsally convex zygosphene to that in VMNH 129332 occurs broadly in extant and extinct Constrictores (see Holman 1977; Smith 2013; Georgalis et al. 2021; Onary et al. 2021; Szyndlar and Georgalis 2023).
Neural canal.—A dorsoventrally tall and large neural canal relative to the zygosphene and preserved intervertebral articulations in VMNH 129332 resembles the bolyeriid Casarea dussumieri (MCZ 49135), Calabaria reinhardtii (see Szyndlar and Georgalis 2023: figs. 63–67), and Loxocemus bicolor (Szyndlar and Georgalis 2023: figs. 139–140). Most other extant Constrictores have a proportionally shorter and narrower canal. Among extinct taxa, Boavus brevis (see Onary et al. 2021: fig. 2) and Totlandophis spp. (see Holman and Harrison 1998a: figs. 1A, 2A; Holman and Harrison 2001: figs. 1A, 2A) bear similarly tall and proportionally broad canals.
Neural spine.—Relatively anteroposteriorly short and moderately tall neural spines occur in many Constrictores, including Calabaria reinhardtii (SMF-PH 68), sanziniids (e.g., YPM R 11921, 11241 and see Szyndlar and Georgalis 2023: figs. 71–75), and Eunectes murinus (CM 145331; Szyndlar and Georgalis 2023). The neural spine being mostly developed on the posterior half of the neural arch, as in VMNH 129332, is rarer; a similar condition occurs in distantly related members of Constrictores such as Eunectes murinus (CM 145331), Xenopeltis unicolor (Szyndlar and Georgalis 2023: fig. 129).
Extinct taxa, with similar mostly posteriorly developed spines occur in Huberophis georgiensis (see Holman 1977: fig. 2C), Totlandophis thomasae (see Holman and Harrison 1998b: figs. 1C, 2C), Eoconstrictor fischeri (see Schaal 2004; Georgalis et al. 2021; Onary et al. 2021: fig. 5C), and Eoconstrictor barnesi (Palci et al. 2024: figs. 10C, H). Anteroposteriorly longer neural spines that occupy much of the anteroposterior lengths of the spine are more common, as in Boavus spp. (see Onary et al. 2021: figs. 1C–H, 2), Palaeopython spp. and Phosphoroboa filholii (Georgalis et al. 2021), and Tallahattaophis dunni (see Holman and Case 1988: fig. 3A, E).
Discussion
Biogeography.—VMNH 129332 is supported as a member of Constrictores by a number of morphological characters, although no character or combination of characters supports it belonging to a specific lineage of Paleogene snake. It joins the pre-existing high taxonomic diversity of non-caenophidian snake lineages known from the Paleogene of western Europe and western North America.
Three constrictor taxa have been named from the eastern USA previously, viz. Huberophis georgiensis and Ogmophis voorhiesi from the Twiggs Clay of Georgia, USA and Tallahattaophis dunni from the Tallahatta Formation of Alabama, USA. Both H. georgiensis and T. dunni were named based on isolated vertebrae and are considered Alethinophidia incertae sedis by Smith and Georgalis (2022). None of the described specimens resemble VMNH 129332, indicating that the latter may be a new taxon for eastern North America.
VMNH 129332 also represents a first as the northernmost known record of a Paleogene snake from the eastern USA and the first from the Atlantic Coast. Palaeogeographic reconstructions indicate that the localities from which the Georgian and Alabaman snakes were recovered existed along the ancient Gulf Coast of North America. Thus, the Virginian constrictor is geographically closest to the land connections between Europe and North America that existed during the Paleogene.
Distributions for squamate lineages across North America and western Europe are common, supporting dispersal from the former to the latter facilitated by land corridors around the time of the Palaeocene–Eocene Thermal Maximum (PETM). These distributions of squamate lineages, as well as those of early Eocene mammals (see Rose 1999; Rose et al. 2012), have been used to support a common Euro-American terrestrial vertebrate fauna during this interval (Savage 1971; Smith 2017; Smith and Scanferla 2021; Georgalis and Mennecart 2025). Well-supported examples include corytophanids (Smith 2009; Conrad 2015; Smith and Scanferla 2016), polychrotids (Smith and Gauthier 2013; Smith et al. 2018), agamids, glyptosaurids (Smith and Gauthier 2013; Čerňanský et al. 2023, 2024), shinisaurids (Conrad 2006; Smith 2017), varanids (Augé 2005; Augé et al. 2022; Smith 2011; Smith et al. 2018; Čerňanský et al. 2024), and the snakes Dunnophis (Hecht 1959; Rage 1973, 1974) and Cheilophis (Georgalis and Mennecart 2025). A phylogenetic analysis also recovered the recently described “erycine” booid Rageryx schmidi from the Messel Pit of Germany (Smith and Scanferla 2021) on the stem of the otherwise North American clade Charinainae. The presence of a member of Constrictores in the mid-Atlantic region geographically intermediate between those in western Europe and western North America supports this narrative. Future discoveries of terrestrial snake fossils from the Nanjemoy Formation, especially those referrable to known lineages, could be key to supporting the hypothesis of a continuous squamate fauna across Euro-America during the early Paleogene.
Taphonomic hypothesis.—The extensive erosion of the surface bone in VMNH 129332, and especially the bilateral opening of the zygantral roof, is noteworthy. The bilateral opening of the zygantral roof, in particular, has been described only twice before, as far as we are aware: in some vertebrae attributed to Palaeopython ceciliensis from the Eocene of Geiseltal, Germany (Georgalis et al. 2021: fig. 28) and in the holotype and paratypes of Coprophis dakotaensis, which were recovered from a coprolite (Parris and Holman 1978; Georgalis et al. 2021). The pattern of damage to the cortical bone surface (Fig. 3) suggests corrosion in the gut tract of a predator, such as a crocodylian. Rounding of projecting structures, such as zygapophyses and neural spine, can result from both abrasion and digestion (e.g., Andrews 1990; Fernández Jalvo and Andrews 2016). While these projecting structures are eroded in VMNH 129332, erosion of the cortical surface is also present under the zygosphene and under the postzygapophyses, which are not projecting structures. Furthermore, all surfaces of synovial joints (the paradiapophyses and condyle) are eroded. Lev et al. (2020) found that the “regularity” of the holes in the surfaces of synovial joints, as in VMNH 129332, was characteristic of digestion rather than abrasion. Finally, there are cracks associated with many of the areas of damaged cortical bone, particularly on the lateral surfaces of the neural arch. While cracking is associated with both weathering and corrosion (Andrews 1990; Fernández Jalvo and Andrews 2016; Fernández Jalvo et al. 2014), the extensive unaltered surfaces on the lateral surface of the neural arch in VMNH 129332 argue against weathering. Finally, the paired opening of the zygantral roof is particularly noteworthy, because that roof, even when zygantral mounds (Hecht 1953; “arcual ridges” of Scanferla and Canale 2007) are present, are not nearly as prominent as the neural spine. However, the zygantral roof is very thin, often without cancellous bone (Martin Ivanov, personal communication 2025), in these areas to accommodate the projecting corners of the zygosphene, so that very little cortical bone surface need be removed to open the zygantral roof. The similar damage to the holotype and paratypes of Coprophis dakotaensis, all recovered from within a coprolite, is additional evidence that corrosion in a predator’s digestive tract is sufficient to produce similar damage. Whether hydrochloric acid or proteases or a combination thereof (both contribute to erosion of bone, e.g., Denys et al. 1995; Fernández Jalvo et al. 2014; Smith et al. 2021) are responsible for the modifications in VMNH 129332 cannot be determined at present.
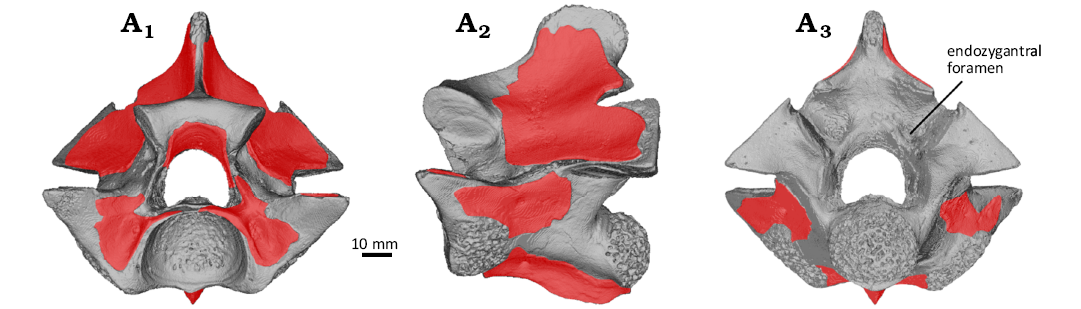
Fig. 3. 3D surface rendering of vertebra of the terrestrial snake Constrictores indet. (VMNH 129332) from the lower Eocene of Fisher/Sullivan site, Virginia, USA. Anterior (A1), left lateral (A2), and posterior (A3) views. Red overlays indicate the extent of preservation of the outermost layer of cortical bone.
Conclusions
The discovery of VMNH 129332 supports the presence of Constrictores on the Paleogene Atlantic Coast during the early Eocene, supporting a continuous distribution of the clade across western Europe and North America. However, in the absence of apomorphies or a combination of characters comparable to any known lineages, VMNH 129332 does not illuminate specific taxonomic connection to any specific snake assemblage in North America or in Europe. Nevertheless, this taxon, and any other records of terrestrial reptiles from Nanjemoy Formation localities, represent crucial mid-Atlantic records at the intersection between the terrestrial reptile assemblages in the American west, the Gulf Coast region, and western Europe. Future analysis and discovery of terrestrial reptiles in the Nanjemoy could illuminate routes of dispersal, regions of endemism, or centers of origin in Paleogene reptile clades. Erosion of outer layers of bone and specific points of damage support the hypothesis that the vertebra was digested prior to fossilization.
Acknowledgements
We thank Jeff Carpenter (Fairfax, USA), Mike Folmer (Millersville, USA), Gary Grimsley (Annandale, USA), and Marco Gulotta (King George, USA) for their ongoing work in the Nanjemoy Formation of northern Virginia and their contributions to the collections of the Virginia Museum of Natural History. Special thanks go to Mike Folmer for his contribution of VMNH 129332 to the VMNH collections and to Gary Grimsley for his laser-focused copy-editing. We thank Liberty Hightower, Jackson Means, and Kal Ivanov (all VMNH) for use of and training on the VMNH entomology photography setup. We thank Michelle Stocker and Sterling Nesbitt (Virginia Tech, Blacksburg, USA) for assistance with microCT scanning. We also thank Martin Ivanov (Masaryk University, Brno, Czech Republic) and an anonymous reviewer for comments that greatly improved this manuscript. We thank The Objects for making Dragonfly 3D licenses available free for non-profit research usage. Support to AP for this work was provided by the Virginia Museum of Natural History Foundation. GG acknowledges funding from the research project no. 2023/49/B/ST10/02631 financed by the National Science Center (Narodowe Centrum Nauki) of Poland.
Editor: Daniel Barta
References
Andrews, P. 1990. Owls, Caves and Fossils. 231 pp. University of Chicago Press, Chicago.
Auffenberg, W. 1963. The fossil snakes of Florida. Tulane Studies in Zoology 10: 131–216. Crossref
Augé, M. 2005. Evolution des lézards du Paléogène en Europe. Mémoires du Muséum national d’Histoire naturelle, Paris 192: 1–369.
Augé, M.L., Folie, A., Smith, R., Phélizon, A., Gigase, P., and Smith, T. 2022. Revision of the oldest varanid, Saniwa orsmaelensis Dollo, 1923, from the earliest Eocene of northwest Europe. Comptes Rendus Palevol 21 (25): 511–529. Crossref
Čerňanský, A., Smith, R., Smith, T., and Folie, A. 2023. Iguanian lizards (Acrodonta and Pleurodonta) from the earliest Eocene (MP 7) of Dormaal, Belgium: the first stages of these iconic reptiles in Europe. Journal of Vertebrate Paleontology 42: e2184696. Crossref
Čerňanský, A., Smith., R., Smith, T., and Folie, A. 2024. Timing of intercontinental faunal migrations: Anguimorph lizards from the earliest Eocene (MP 7) of Dormaal, Belgium. Zoological Journal of the Linnean Society 201: zlae082. Crossref
Conrad, J.L. 2006 An Eocene shinisaurid (Reptilia, Squamata) from Wyoming, U.S.A. Journal of Vertebrate Paleontology 26: 113–126. Crossref
Conrad, J.L. 2015 A New Eocene Casquehead Lizard (Reptilia, Corytophanidae) from North America. PLOS ONE 10: e0127900. Crossref
Denys, C., Fernández-Jalvo, Y., and Dauphin, Y. 1995. Experimental taphonomy: preliminary results of the digestion of micromammal bones in the laboratory. Comptes Rendus de l’Académie des Sciences de Paris, Série IIA 321: 803–809.
Fernández-Jalvo, Y. and Andrews, P. 2016. Atlas of Taphonomic Identifications: 1001+ Images of Fossil and Recent Mammal Bone Modification. 359 pp. Springer, Dordrecht. Crossref
Fernández-Jalvo, Y., Andrews, P., Sevilla, P., and Requejo, V. 2014. Digestion versus abrasion features in rodent bones. Lethaia 47: 323–336. Crossref
Georgalis, G.L. and Mennecart, B. 2025. Another wanderer from the Paleocene–Eocene Thermal Maximum? A new species of the North American snake genus Cheilophis from the early Eocene of France. In: G.L. Georgalis, H. Zaher, and M. Laurin (eds.), Snakes from the Cenozoic of Europe—Towards a Macroevolutionary and Palaeobiogeographic Synthesis. Comptes Rendus Palevol 24: 317–324. Crossref
Georgalis, G.L. and Smith, K.T. 2020. Constrictores Oppel, 1811—the available name for the taxonomic group uniting boas and pythons. Vertebrate Zoology 70: 291–304.
Georgalis, G.L., Rabi, M., and Smith, K.T. 2021. Taxonomic revision of the snakes of the genera Palaeopython and Paleryx (Serpentes, Constrictores) from the Paleogene of Europe. Swiss Journal of Palaeontology 140: 1–140. Crossref
Gilmore, C.W. 1938. Fossil snakes of North America. Geological Society of America, Special Papers 9: 1–96. Crossref
Hecht, M.K. 1959. Amphibians and reptiles. In: P.O. McGrew, J.E. Berman, M.K. Hecht, J.M. Hummel, G.G. Simpson, and A.E. Wood (eds.), The Geology and Paleontology of the Elk Mountain and Tabernacle Butte Area, Wyoming. Bulletin of American Museum of Natural History 117: 130–146.
Hilton, E.J. and Carpenter, J. 2020. Bony-tongue fishes (Teleostei: Osteoglossomorpha from the Eocene Nanjemoy Formation, Virginia. Northeastern Naturalist 27: 1–34. Crossref
Holman, J.A. 1977. Upper Eocene snakes (Reptilia, Serpentes) from Georgia. Journal of Herpetology 11: 141–145. Crossref
Holman, J.A. 2000. Fossil Snakes of North America: Origin, Evolution, Distribution, Paleoecology. 357 pp. Indiana University Press, Bloomington.
Holman, J.A. and Case, G.R. 1988. Reptiles from the Eocene Tallahatta Formation of Alabama. Journal of Vertebrate Paleontology 8: 328–333. Crossref
Holman, J.A. and Harrison, D.L. 1998a. A new genus of small boid snake from the Upper Eocene of Hordle Cliff, Hampshire, England. Acta Zoologica Cracoviensia 41: 29–33.
Holman, J.A. and Harrison, D.L. 1998b. A new genus of snake (Serpentes: Boidae) from the Upper Eocene of Hordle Cliff, Hampshire, England. Acta Zoologica Cracoviensia 41: 23–27.
Holman, J.A. and Harrison, D.L. 2001. Early Oligocene (Whitneyan) snakes from Florida (USA): remaining boids, indeterminate colubroids, summary and discussion of the I-75 Local Fauna snakes. Acta Zoologica Cracoviensia 44: 25–36.
Houssaye, A., Boistel, R., Böhme, W., and Herrel, A. 2013. Jack-of-all-trades master of all? Snake vertebrae have a generalist inner organization. Naturwissenschaften 100: 997–1006. Crossref
Kent, B.W. 1999a. Rays from the Fisher/Sullivan Site. In: R.E. Weems and G.J. Grimsley (eds.), Early Eocene Vertebrates and Plants from the Fisher/Sullivan Site (Nanjemoy Formation) Stafford County, Virginia, 39–52. Commonwealth of Virginia Department of Mines, Minerals and Energy, Charlottesville.
Kent, B.W. 1999b. Sharks from the Fisher/Sullivan Site. In: R.E. Weems and G.J. Grimsley (eds.), Early Eocene Vertebrates and Plants from the Fisher/Sullivan Site (Nanjemoy Formation) Stafford County, Virginia, 11–38. Commonwealth of Virginia Department of Mines, Minerals and Energy, Charlottesville.
La Duke, T.C. 1991. The fossil snakes of Pit 91, Rancho La Brea, California. Contributions in Science 424: 1–28. Crossref
Lev, M., Weinstein-Evron, M., and Yeshurun, R. 2020. Squamate bone taphonomy: a new experimental framework and its application to the Natufian zooarchaeological record. Scientific Reports 10: 9373. Crossref
Mayr, G., De Pietri, V., and Scofield, P. 2022. New bird remains from the early Eocene Nanjemoy Formation of Virginia (USA), including the first records of the Messelasturidae, Psittacopedidae, and Zygodactylidae from the Fisher/Sullivan site. Historical Biology 34: 322–334. Crossref
McCartney, J., Roberts, E., Tapanila, L., and O’Leary, M. 2018. Large palaeophiid and nigerophiid snakes from Paleogene Trans-Saharan Seaway deposits of Mali. Acta Palaeontologica Polonica 63: 207–220. Crossref
Olson, S.L. 1999. Early Eocene birds from eastern North America: a faunule from the Nanjemoy Formation of Virginia. In: R.E. Weems and G.J. Grimsley (eds.), Early Eocene Vertebrates and Plants from the Fisher/Sullivan Site (Nanjemoy Formation) Stafford County, Virginia, 123–132. Commonwealth of Virginia Department of Mines, Minerals and Energy, Charlottesville.
Onary, S., Hsiou, A.S., Lee, M.S.Y., and Palci, A. 2021. Redescription, taxonomy and phylogenetic relationships of Boavus Marsh, 1871 (Serpentes: Booidea) from the early–middle Eocene of the USA. Journal of Systematic Palaeontology 19: 1601–1622. Crossref
Palci, A., Onary, S., Lee, M.S.Y., Smith, K.T., Wings, O., Rabit, M., and Georgalis, G. 2024. A new booid snake from the Eocene (Lutetian) Konservat-Lagerstätte of Geiseltal, Germany, and a new phylogenetic analysis of Booidea. Zoological Journal of the Linnean Society 202: zlad179. Crossref
Parris, D.C. and Holman, J.A. 1978. An Oligocene snake from a coprolite. Herpetologica 34: 258–264.
Petermann, H. and Gauthier, J.A. 2018. Fingerprinting snakes: paleontological and paleoecological implications of zygantral growth rings in Serpentes. PeerJ 6: e4819. Crossref
Rage, J.-C. 1973. Présence de Dunnophis (Reptilia, Serpentes) dans l’Eocène et l’Oligocène europeens. Compte rendu sommaire des séances de la Société Géologique de France 1973 (3): 76–78.
Rage, J.-C. 1974. Les serpents des phosphorites du Quercy. Palaeovertebrata 6: 274–303.
Rage, J.-C. 1978. La poche à phosphate de Ste. Néboule (Lot) et sa faune de vertébrés du Ludien Supérieur. Oiseaux. 5. Squamates. Palaeovertebrata 8: 201–215.
Rage, J.-C. 1984. Serpentes. Part 11. Handbuch Der Paläoherpetologie. 80 pp. Gustav Fisher Verlag, Stuttgart.
Rose, K.D. 1999. Fossil mammals from the early Eocene Fisher/Sullivan site. In: R.E. Weems and G.J. Grimsley (eds.), Early Eocene Vertebrates and Plants from the Fisher/Sullivan Site (Nanjemoy Formation) Stafford County, Virginia, 133–138. Commonwealth of Virginia Department of Mines, Minerals and Energy, Charlottesville.
Rose, K.D., Chew, A.E., Dunn, R.H., Kraus, M.J., Fricke, H.C., and Zack, S.P. 2012. Earliest Eocene mammalian fauna from the Paleocene–Eocene Thermal Maximum at Sand Creek Divide, southern Bighorn Basin, Wyoming. Contributions from the Museum of Paleontology, the University of Michigan 36: 1–122.
Rose, K.D., Perry, J.M.G., Prufrock, K.A., and Weems, R.E. 2021. Early Eocene omomyid from the Nanjemoy Formation of Virginia: first fossil primate from the Atlantic Coastal Plain. Journal of Vertebrate Paleontology 41: e1923340. Crossref
Savage, D.E. 1971. The Sparnacian–Wasatchian mammalian fauna, Early Eocene, of Europe and North America. Abhandlungen des Hessischen Landesamtes für Bodenforschung 60: 154–158.
Scanferla, C.A. and Canale, J. I. 2007. The youngest record of the Cretaceous snake genus Dinilysia (Squamata, Serpentes). South American Journal of Herpetology 2: 76–81. Crossref
Scanferla, C.A. and Smith, K.T. 2020. Exquisitely preserved fossil snakes of Messel: Insight into the evolution, biogeography, habitat preferences and sensory ecology of early boas. Diversity 12: 1–16. Crossref
Schaal, S.F.K. 2004. Palaeopython fischeri n. sp. (Serpentes: Boidae), eine Riesenschlange aus dem Eozän (MP11) von Messel. Courier Foschunginstitut Senckenberg 252: 35–100.
Smith, K.T. 2009. A new lizard assemblage from the earliest Eocene (Zone Wa0) of the Bighorn Basin, Wyoming, USA: Biogeography during the warmest interval of the Cenozoic. Journal of Systematic Palaeontology 7: 299–358. Crossref
Smith, K.T. 2011. On the phylogenetic affinity of the extinct acrodontan lizard Tinosaurus. Bonner Zoologische Monographien 57: 9–28.
Smith, K.T. 2013. New constraints on the evolution of the snake clades Ungaliophiinae, Loxocemidae and Colubridae (Serpentes), with comments on the fossil history of erycine boids in North America. Zoologischer Anzeiger 252: 157–182. Crossref
Smith, K.T. 2017. First crocodile-tailed lizard (Squamata: Pan-Shinisaurus) from the Paleogene of Europe. Journal of Vertebrate Paleontology 37: e1313743. Crossref
Smith, K.T. and Gauthier, J.A. 2013. Early Eocene lizards of the Wasatch Formation near Bitter Creek, Wyoming: Diversity and paleoenvironment during an interval of global warming. Bulletin of the Peabody Museum of Natural History 54: 135–230. Crossref
Smith, K.T. and Georgalis, G.L. 2022. The diversity and distribution of Palaeogene snakes: a review with comments on vertebral sufficiency. In: D.J. Gower and H. Zaher (eds.), The Origin and Early Evolution of Snakes, 55–84. Cambridge University Press, Cambridge. Crossref
Smith, K.T. and Scanferla, A. 2016. Fossil snake preserving three trophic levels and evidence for an ontogenetic dietary shift. Palaeobiodiversity and Palaeoenvironments 96: 589–599. Crossref
Smith, K.T. and Scanferla, A. 2021. A nearly complete skeleton of the oldest definitive erycine boid (Messel, Germany). Geodiversitas 43: 1–24. Crossref
Smith, K.T. and Scanferla, A. 2022. More than one large constrictor snake lurked around paleolake Messel. Palaeontographica A 323: 1–3. Crossref
Smith, K.T., Bhullar, B.-A.S., Köhler, G., and Habersetzer, J. 2018. The only known jawed vertebrate with four eyes and the bauplan of the pineal complex. Current Biology 28: 1101–1107. Crossref
Smith, K.T., Comay, O., Maul, L., Wegmüller, F., Le Tensorer, J.-M., and Dayan, T. 2021. A model of digestive tooth corrosion in lizards: experimental tests and taphonomic implications. Scientific Reports 11: 12877. Crossref
Szyndlar, Z. and Georgalis, G.L. 2023 An illustrated atlas of the vertebral morphology of extant non-caenophidian snakes, with special emphasis on the cloacal and caudal portions of the column. Vertebrate Zoology 73: 717–886. Crossref
Warren, J.W. 1963. Growth Zones in the Skeleton of Recent and Fossil Vertebrates. 142 pp. Ph.D. Dissertation, University of California, Los Angeles.
Weems, R.E. 1999a. Actinopterygian fishes from the Fisher/Sullivan Site. In: R.E. Weems and G.J. Grimsley (eds.), Early Eocene Vertebrates and Plants from the Fisher/Sullivan Site (Nanjemoy Formation) Stafford County, Virginia, 53–100. Commonwealth of Virginia Department of Mines, Minerals and Energy, Charlottesville.
Weems, R.E. 1999b. Reptile remains from the Fisher/Sullivan site. In: R.E. Weems and G.J. Grimsley (eds.), Early Eocene Vertebrates and Plants from the Fisher/Sullivan Site (Nanjemoy Formation) Stafford County, Virginia, 101–122. Commonwealth of Virginia Department of Mines, Minerals and Energy, Charlottesville.
Weems, R.E. 2020. Additions to the bony fish fauna from the early Eocene Nanjemoy Formation of Maryland and Virginia (U.S.A.). The Mosasaur 9: 117–152.
Weems, R.E. and Grimsley, G.J. 1999. Introduction, geology, and paleogeographic setting. In: R.E. Weems and G.J. Grimsley (eds.), Early Eocene Vertebrates and Plants from the Fisher/Sullivan Site (Nanjemoy Formation) Stafford County, Virginia, 1–10. Commonwealth of Virginia Department of Mines, Minerals and Energy, Charlottesville.
Zaher, H., Mohabey, D.M., Grazziotin, F.G., and Wilson Mantilla, J.A. 2023. The skull of Sanajeh indicus, a Cretaceous snake with an upper temporal bar, and the origin of ophidian wide-gaped feeding. Zoological Journal of the Linnean Society 197: 656–697. Crossref
Zaher, H., Murphy, R.W., Arredondo, J.C., Graboski, R., Machado-Filho, P.R., Mahlow, K., Montingelli, G.-G., Bottallo Quadros, A.B., Orlov, N.L., Wilkinson, M., Zhang Y.-P., and Grazziotin, F.G. 2019. Large-scale molecular phylogeny, morphology, divergence-time estimation, and the fossil record of advanced caenophidian snakes (Squamata: Serpentes). PLOS ONE 14: e0216148. Crossref
Zaher, H., Trusz, C., Koch, C., Entiauspe-Neto, O.M., Battilana, J., and Grazziotin, F.G. 2024. Molecular phylogeny and biogeography of the dwarf boas of the family Tropidophiidae (Serpentes: Alethinophidia. Systematics and Biology 22: 2319289. Crossref
Acta Palaeontol. Pol. 70 (4): 817–826, 2025
https://doi.org/10.4202/app.01265.2025