
Parasitism in Jurassic belemnites: examples from the upper Callovian of southern Poland
SREEPAT JAIN, WOJCIECH KRAWCZYŃSKI, MARIUSZ A. SALAMON, PIOTR DUDA, and SIMON F. MITCHELL
Jain, S., Krawczyński, W., Salamon, M.A., Duda, P., and Mitchell, S.F. 2025. Parasitism in Jurassic belemnites: examples from the upper Callovian of southern Poland. Acta Palaeontologica Polonica 70 (4): 755–764.
Deformation in specimens of Hibolithes hastatus and Rhopaloteuthis spp. are documented from the upper Callovian Lamberti Zone of the Ogrodzieniec quarry, southern Poland. A specimen with hook-shaped bending is assigned to forma hamata, whereas other specimens with local thickening of the rostrum are assigned to another paleopathy, forma bullata. The absence of any external injury corroborated by high-resolution computed tomography suggests parasitism as the most probably cause for shell deformation. Identification of a specific parasite taxon is, however, hampered by the fact that parasites are typically small-bodied (submillimeter) and generally lack biomineralized hard parts. We argue that different parasites can leave similar traces and taxonomically distant parasites can inflict similar symptoms on their hosts due to convergence in the evolution of host-exploitation strategies. and microtomography offers a non-destructive way to analyze such structures in belemnite rostra. We show that the scan-based evidence provides a clearer picture of the internal structure of the paleopathy and suggests that the Keupp’s classification of paleopathies based only on external features might lead to false inferences of their formational mechanisms.
Key words: Belemnitida, Hibolithes, Rhopaloteuthis, paleopathy, parasite, Callovian, Poland.
Sreepat Jain [sreepatjain@gmail.com; ORCID: https://orcid.org/0000-0002-7679-9248 ], Adama Science and Technology University, Department of Applied Geology, School of Applied Natural Sciences, P.O. Box 1888, Adama, Ethiopia.
Wojciech Krawczyński [wojciech.krawczynski@us.edu.pl; ORCID: https://orcid.org/0000-0002-0555-8420 ] and Mariusz A. Salamon [paleo.crinoids@poczta.fm; ORCID: https://orcid.org/0000-0001-9399-2798 ], University of Silesia in Katowice, Faculty of Natural Sciences, Institute of Earth Sciences, ul. Będzińska 60, 41-200 Sosnowiec, Poland.
Piotr Duda [piotr.duda@us.edu.pl; ORCID: https://orcid.org/0000-0003-0867-6315 ], University of Silesia in Katowice, Faculty of Science and Technology, ul. Będzińska 39, 41-200 Sosnowiec, Poland.
Simon F. Mitchell [simon.mitchell@uwimona.edu.jm, barrettia2000@yahoo.co.uk; ORCID: https://orcid.org/0000-0002-7069-0188 ], Department of Geography and Geology, The University of the West Indies, Mona, Kingston 7, Jamaica.
Received 7 July 2025, accepted 3 September 2025, published online 10 December 2025.
Copyright © 2025 S. Jain et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
Belemnites are the most common fossil coleoid cephalopods of the Mesozoic (Dera et al. 2016; Hoffmann and Stevens 2020), with a frequently preserved calcitic rostrum. Hence, they provide an excellent tool to document and infer the internal structure of any paleopathy and their probable formation mechanisms.
High-resolution computed tomography (CT) offers an innovative and non-destructive way to analyze the internal structure of a belemnite rostra and identify various paleopathies. CT-scan images not only reveal internal structures that allow for the examination of a variety of pathological phenomena but also enable differentiation between different types of paleopathy. More importantly, the CT-scan data also allows the inference of potential causes of such deformations (predator attack or parasite activity). Therefore, it allows us to go beyond the classification of pathologies based on surface expression of a specimen alone (sensu Keupp 2012). Table 1 provides a short description of the various types (forma) of paleopathies developed for the belemnite rostra based on such surface expressions alone (sensu Keupp 2012; see also Keupp et al. 2020 and references therein).
Table 1. Belemnite paleopathologies and their link with parasitism. The table provides a short description of the various types (forma) of paleopathies developed for the belemnite rostra (after Keupp et al. 2020 and references therein). Shaded rows are paleopathies reported in this contribution. Those marked in bold are paleopathies attributed to or tentatively attributed to parasitism (Keupp 2012; Keupp et al. 2020 and references therein).
|
Type |
Short description |
Reference |
|
Forma |
This paleopathy is marked by multiple fractures of the rostrum, where dislocated fragments are covered by post-traumatic growth layers, resulting in variable bends or a screw-like morphology in different planes. The condition can also involve rostral fragments oriented sub-parallel to each other or a reversal of growth direction towards the animal’s head. |
Duval-Jouve 1841; Kabanov 1967; Doyle 1990; Kraus 2000; Keupp 2002, 2012: fig. 397; Mietchen et al. 2005; Hüne and Hüne 2008; Moosleitner 2012; Keupp et al. 2020 |
|
Forma |
This paleopathy is characterized by the formation of ridges composed of organic material, resulting from proliferating scar tissue of the mantle sac epithelium, which in turn creates furrows on the surface of the rostrum that are oriented perpendicular to the regular longitudinal furrows. |
Keupp et al. 2020: fig. 6.2 |
|
Forma
|
This paleopathy features a hollow space, suggesting potential occupation by an earlier endoparasite. In rare instances, localized mineralization of the rostrum can lead to the development of a secondary rostrum paralleling the primary one, supporting a parasitic origin for the formation of this paleopathy. Herein we demonstrate parasitism as the probable causes for this paleopathy. |
Keupp 2012: fig. 405; Hoffmann et al. 2018; Keupp et al. 2020: fig. 6.3; this study |
|
Forma |
This paleopathy is characterized by a club-shaped rostrum that is thicker and shorter than normal, resulting from a broken juvenile rostrum that leads to deformation, often affecting the earliest parts of the phragmocone, including the initial chamber, and resulting in a distinctive drop-shaped morphology and significant shortening of the rostrum. |
Keupp 2012: top of fig. 381; Kabanov 1967; Keupp 2012; Keupp et al. 2020: fig. 6.4 |
|
Forma
|
This paleopathy is characterized by a flap-like outgrowth at the subapical rostrum area, occurring without any apparent injury, potentially resulting from a parasitic infestation that triggers the formation of a collar-like structure, indicative of inflammation of the mantle sac epithelium. |
Keupp 2012; Keupp et al. 2020: fig. 6.5 |
|
Forma |
This paleopathy is marked by disruptions in the ventral longitudinal furrows of the rostrum, potentially related to the central blood vessel system, resulting in S-shaped or laterally displaced furrows that suggest temporary dislocations of the soft body, possibly caused by traumatic events like predator attacks or intrinsic disturbances. |
Stevens 1965; Kabanov 1967; Keupp 2012; Keupp et al. 2020: fig. 6.6 |
|
Forma |
This paleopathy results from damage to the mantle epithelium, impairing its ability to secrete normal growth layers, similar to those seen in forma aegra manca. However, the mantle continues to secrete calcite grains rich in organic matter, leading to irregular deposits that create a rough, granular surface on the rostrum. In some cases, isolated fragments of injured epithelium can form spherulithic aggregates or, rarely, free pearls, when the epithelium encloses itself in a shell sac-like structure that temporarily survives in the surrounding cavity. |
Keupp 2012: figs. 382, 384; Keupp et al. 2020: fig. 6.7 |
|
Forma
|
This paleopathy is characterized by hook-shaped rostra, typically resulting from fractures during early developmental stages. Following the fracture, parts of the rostrum become dislocated, and subsequent growth occurs more prominently on one side, altering the direction of growth. In severe cases where the rostrum becomes significantly shortened, a transitional form between hook-shaped and club-shaped morphologies can occur. Notably, this condition has been attributed to injuries and growth defects rather than parasitism, and its association with trauma has been the prevailing interpretation. Herein we demonstrate parasitism as one of the probable causes for this paleopathy. |
Blainville 1827; Duval-Jouve 1841; Radwańska and Radwański 2004; Moosleitner 2006; Keupp 2012; Keupp et al. 2020: fig. 6.8; this study |
|
Forma
|
This paleopathy results from mechanical injury to the mantle sac epithelium, leading to a partial absence of growth layers due to impaired secretion function. The condition often affects the apical area, where the apex is incompletely developed, with the juvenile, pre-traumatic portion forming the tip. Additionally, the rostrum may become thinner on the side of the injury, giving it a scepter-like shape. |
Keupp 2012; Keupp et al. 2020: fig. 6.9 |
|
Forma
|
This paleopathy involves the duplication or multiplication (up to 5) of the rostral tip, potentially resulting from trauma or parasitic infestation. In this condition, the mantle epithelium responsible for apex formation becomes divided into separate, locally independent secretion centers, each forming its own apex. |
Finzel 1963; Schmid 1963; Miertzsch 1964; Keupp 2002, 2012; Mietchen et al. 2005; Hoffmann and Weissmüller 2018; Keupp et al. 2020: fig. 6.10 |
|
Forma |
This paleopathy features linear scratch marks on the surface of belemnite rostra, classified as bite traces. These bite traces can persist in post-traumatic growth layers due to scarring, resulting from the formation of scar tissue in the mantle sac epithelium. |
Riegraf 1973; Hölder 1973; Keupp et al. 2020: fig. 6.11 |
The usefulness and feasibility of the application of non-invasive imaging methods such as computed tomography to the field of paleopathology has recently been demonstrated through a series of studies (Wisshak et al. 2017; Hoffmann et al. 2014, 2018, 2020; Stevens et al. 2020; Jain et al. 2022b and references therein). The CT-scan data gained can be further used for the identification of various diagenetic alteration features in belemnite rostra or even bioerosion trace marks preserved in belemnite guards (Wisshak et al. 2017).
The significance of this contribution stems from the fact that most pathological specimens described so far (particularly those linked with parasitism) are from the Cretaceous (see review in Frerichs 2015) while Jurassic reports, such as this one, are relatively rare (see De Baets et al. 2021a, c). Additionally, on a much broader context of parasitism, recently, De Baets et al. (2021c) based on rigorous analyses (of marine parasite–host interactions among 10 host phyla and 13 parasite phyla across the Phanerozoic) noted that parasitism has increased through the Phanerozoic and that there is a positive correlation between diversity and parasitism (see also Baumiller and Gahn 2002), i.e., an evidence of increased parasitism with biodiversity (the amplification hypothesis). Thus, the distribution of paleopathies through time, space and phylogeny can also serve as a potential link with biodiversity (Baumiller and Gahn 2002; De Baets et al. 2021c). This study contributes to the basic framework for such rigorous analysis to be conducted in future.
Herein, we have used two approaches to study deformed belemnite rostra from the upper Callovian rocks of the Ogrodzieniec quarry, southern Poland (Figs. 1, 2): high-resolution computed-tomography, and external features (sensu Keupp 2012), to infer the type of paleopathy (see also Keupp et al. 2020). A high-resolution CT-scan is presented for a specimen of Hibolithes hastatus (Montfort, 1808) (Fig. 3), and from the same level, several specimens of Rhopaloteuthis spp. (Fig. 4) are documented to show malformations based on surface expressions (malformations) (sensu Keupp 2012). Additionally, a high-resolution CT-scan of one of the representative specimens of Rhopaloteuthis spp. was also done (Fig. 5).
Institutional abbreviations.—GIUS, Faculty of Natural Sciences of the University of Silesia in Katowice, Poland.
Historical background
Hoffmann et al. (2018) used high-resolution computed-tomography and analyzed a pathological belemnite rostrum belonging to Goniocamax sp. from the lower Santonian of the Bavnodde Greensand (Denmark). They attributed the malformation of the forma bullata type to the presence of an endogenic parasite during the lifetime of the belemnite (Hoffmann et al. 2018). Their CT-scan revealed an elongated blister partially filled with smaller carbonate components as noted by their irregular shapes (cement and/or sediment). They inferred that this infection (i.e., blister formation) was possibly caused by an endoparasitic polychaetid and/or flatworm and suggested that similar structures, described from other Jurassic and Cretaceous belemnites, might also be attributed to the same group of parasitic animals, suggesting a similar paleoecology for these belemnites (Hoffmann et al. 2018).
Hoffmann et al. (2020) used high-resolution computed tomography (CT) to infer the internal structures of several groups of deformed belemnites, and illustrated a variety of pathologies of Jurassic–Cretaceous belemnite rostra. The studied taxa included examples of Belemnitella sp. (Late Cretaceous, NW Germany), Gonioteuthis spp. (Campanian, NW Germany), Belemnellocamax spp. (Campanian, Sweden), Goniocamax sp. (Santonian, Denmark). Hibolithes jaculoides (Hauterivian, N Germany), Duvalia emerici (late Valanginian, France), Pseudobelus sp. (Valanginian, France), and ?Acrocoelites sp., and Neoclavibelus sp. (Toarcian, SW Germany). The authors used CT-scan to identify several malformed categories that were initially based only on external features (sensu Keupp 2012). The authors noted the following: apex malformation (forma saepia), blister malformation (forma bullata), blunt rostra (forma clavata), bent rostra (forma angulata), hook-shaped rostra (forma hamata), sceptre-shaped rostra (forma manca), and collar formation (forma collata) (Hoffmann et al. 2020). They further noted that most of the pathologies were a result of predator attacks (forma angulata), and only a few were tentatively ascribed to the activity of parasites (such as formas bullata, collata, and saepia) (Hoffmann et al. 2020). They concluded that although an endogenic parasite infection was the most likely trigger of the inferred paleopathies during the lifetime of the belemnite, attributing these deformations to a specific parasite taxon was difficult to impossible (Hoffmann et al. 2020).
Stevens et al. (2020) while studying a rostrum of the Early Cretaceous belemnite Duvalia emerici for blister pearl formation (i.e., a pearl that was fused to the rostrum surface after its initial formation inside the mantle tissue as a free pearl) emphasized that CT-scan-based analysis provides a clearer picture of the internal structure of the paleopathy and that the classification of paleopathologies based only on external features (sensu Keupp 2012) might lead to false inferences of their formation mechanisms.
Geological setting
The upper Callovian–lower Oxfordian sediments at the Ogrodzieniec quarry, southern Poland (Fig. 1) have been well-studied (Różycki 1953; Głowniak 2012; Jain and Mazur 2021; Jain et al. 2022a, b, 2023, 2024; Jain and Salamon 2024) (Fig. 2A). The upper Callovian unit is a belemnite battlefield (Doyle and Macdonald 2007) with weathered to well-preserved fossil fauna, dominated by ammonites and belemnites (Fig. 2B–E). The pathological samples of Hibolithes hastatus (Montfort, 1808) (Fig. 3) and Rhopaloteuthis spp. (Figs. 4, 5) came from bed γ of Głowniak (2012) which has also yielded the upper Callovian ammonite Erymnocerites argoviensis Jeannet, 1951 (Jain and Salamon 2024) (Fig. 2A).
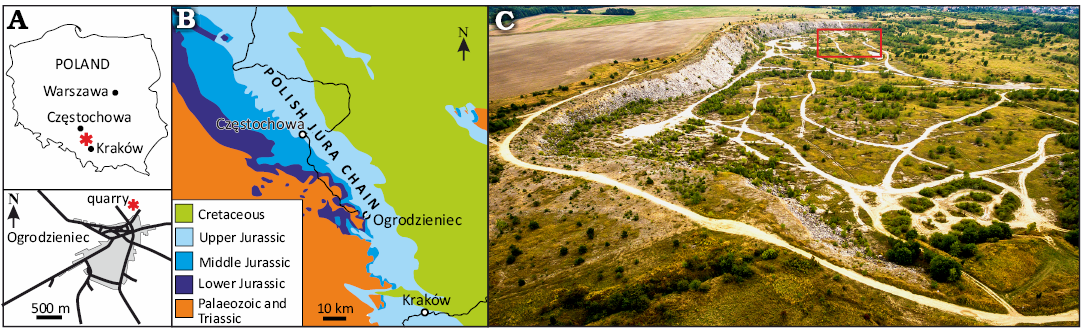
Fig. 1. A. Location of the studied area (asterisk) within Poland and Ogrodzieniec. B. Geological map of the Polish Jura Chain showing the location of Ogrodzieniec. C. Aerial view of the Ogrodzieniec quarry; red rectangle marks the location of the studied section. Modified after Jain and Mazur (2021).

Fig. 2. Biostratigraphy of the studied section (modified after Jain and Salamon 2024). A. Correlation of studied beds with those of Głowniak (2012). B. Geological column and the occurrence of specimens from the top of bed γ recorded in the present study. C. Field photo of the condensed bed where the studied specimens were recorded. D. Close-up of the block with Quenstedtoceras lamberti (Sowerby, 1819) and Hibolithes hastatus (Montfort, 1808).
Material and methods
Five specimens from upper Callovian rocks of the Ogrodzieniec quarry, southern Poland are investigated for paleopathy (malformation). One specimen (GIUS 8-3798/1) (Fig. 3) was analyzed using high-resolution computed-tomography, whereas paleopathologies in 4 other specimens (GIUS 8-3798/ 2‒5) (Figs. 4, 5) were identified based on the traditional concept of external features (sensu Keupp 2012). All specimens are deposited in the collection of the Faculty of Natural Sciences of the University of Silesia in Katowice, Poland. Virtual sections of Hibolithes hastatus were made in the Faculty X-ray Microtomography Laboratory at the Faculty of Computer Science and Material Science, University of Silesia in Katowice, Chorzów, Poland, using the General Electric Phoenix v|tome|x micro-CT equipment at 160 kV, 70μA and a scanning time of 20 min (Davies et al. 2017) (see movies provided in SOM 1–3, Supplementary Online Material available at http://app.pan.pl/SOM/app70-Jain_etal_SOM.pdf). The projection images (Figs. 3, 5) were captured using a 1000×2024 pxs scintillator/CCD with an exposure time of 250 ms and processed using Volume Graphics VGSTUDIO Max software and analyzed using Volume Graphics myVGL viewer. A total of 120 fragmentary belemnite specimens were collected; 5 were pathological (~4%).
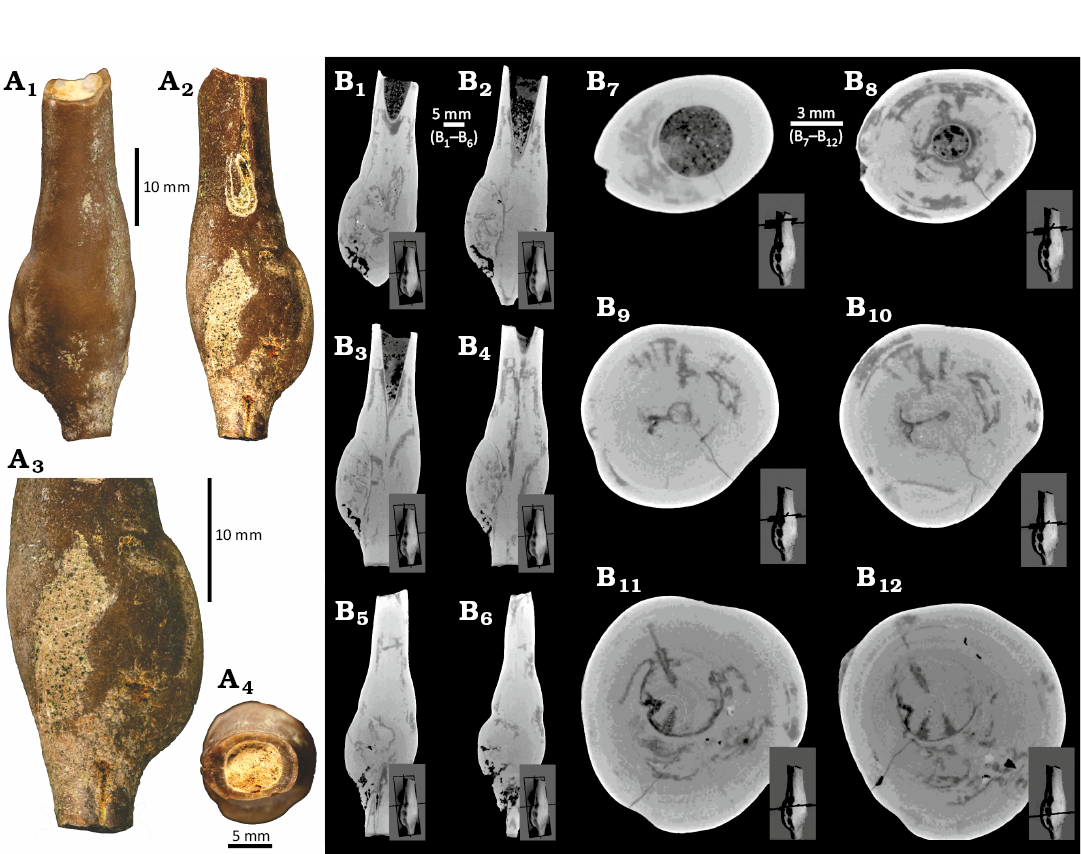
Fig. 3. Pathological specimen of the belemnite Hibolithes hastatus (Montfort, 1808), GIUS 8-3798/1, upper Callovian of Poland (see Jain and Salamon 2024 for details), Ogrodzieniec quarry, bed γ of Głowniak (2012). A. Plane light images, lateral (A1) and opposite lateral (A2) views, close-up showing the bulbous pathology (A3), circular cross-section (A4). B. CT-scans: longitudinal (B1‒B6) and transverse (B7‒B12) sections showing the relatively elongate groove and the bulbous pathological part.
All stack files and movies are uploaded in ZENODO repository: https://doi.org/10.5281/zenodo.16900861.
The belemnite GIUS 8-3798/1 (Fig. 3) has lost its apical portion and has a clearly pathological form. A well-developed groove is present in the alveolar region. The general form of this specimen (excluding its clearly pathological region) is of a relatively elongate subcyclindrical to subhastate morphology. By comparison with Polish belemnites (Pugaczewska 1961) from the upper Callovian, the best match is with Hibolithes hastatus. Four belemnite rostra with a generally different shape than GIUS 8-3798/2–5 have less strongly elongated club-shaped form, and a shorter apical region (Fig. 4). Again, by comparison with figured Polish belemnites from the upper Callovian, they are best assigned to the Rhopaloteuthis spp. As with most pathologically deformed belemnites (as opposed to many of those deformed by predation), an accurate identification a genus level is often tentative at best.

Fig. 4. Pathological specimens of the belemnite Rhopaloteuthis spp., upper Callovian of Poland (see Jain and Salamon 2024 for details), Ogrodzieniec quarry, bed γ of Głowniak (2012). A. GIUS 8-3798/2, forma hamata (Keupp 2012), in lateral view. B‒D. Forma bullata (Keupp 2012). B. GIUS 8-3798/3, in lateral (B1), opposite lateral (B2), and apertural (B3) views. C. GIUS 8-3798/4, in lateral (C1), opposite lateral (C2), and apertural (C3) views. D. GIUS 8-3798/5, in lateral (D1), opposite lateral (D2), and apertural (D3) views. Scale bars 5 mm.
Results
The specimen GIUS 8-3798/1 of Hibolithes hastatus (Montfort, 1808) shows both in its external features (sensu Keupp 2012) (Fig. 3A) and in high-resolution CT scans (Fig. 3B) a clear thickening of the rostrum with no signs of any external injury, and is summarily assigned to forma bullata Keupp (2012) paleopathy.
The specimens of Rhopaloteuthis spp. (Fig. 4) show two types of paleopathologies with no signs of any external injury. GIUS 8-3798/2 in which the rostrum is bent (i.e., hook-shaped bending of Keupp 2012) called forma hamata (Fig. 4A), and the third GIUS 8-3798/2, in which there is swelling (thickening of the rostrum; Keupp 2012) called forma bullata (Fig. 4B‒D). A high-resolution CT-scan of Rhopaloteuthis spp. (i.e., GIUS 8-3798/2) was also performed and reveled no injury that would have otherwise resulted in a hook-shaped bending (Fig. 5).
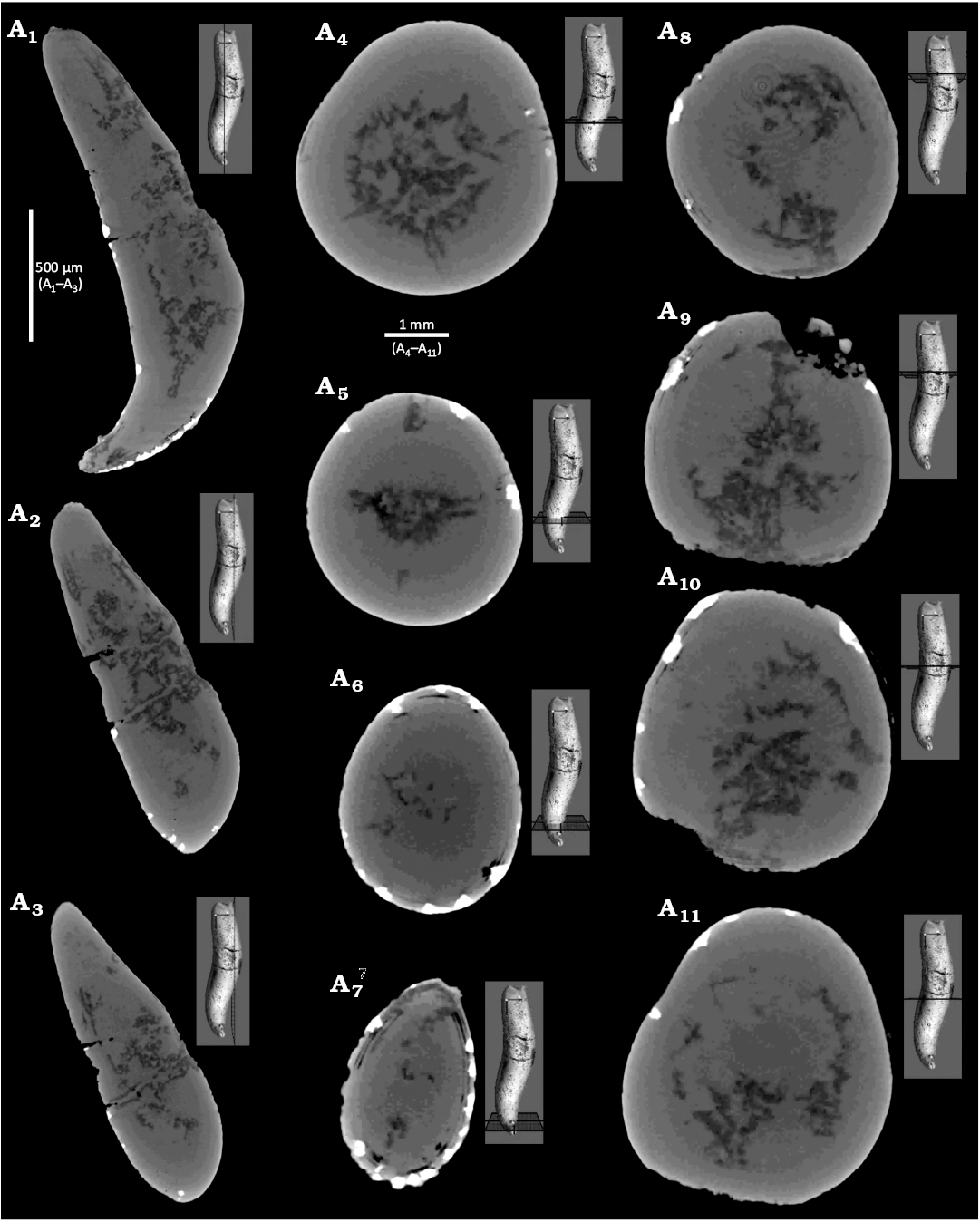
Fig. 5. CT-scans of pathological specimen (GIUS 8-3798/2) of the belemnite Rhopaloteuthis spp., upper Callovian of Poland (see Jain and Salamon 2024 for details), Ogrodzieniec quarry, bed γ of Głowniak (2012). Forma hamata (Keupp 2012), longitudinal (A1‒A3) and transverse (A4‒A11) sections.
Discussion
Computed-tomography on Hibolithes hastatus (GIUS 8-3798/1, Fig. 3) and Rhopaloteuthis spp. (GIUS 8-3798/2, Fig. 5) revealed no external injury. In both H. hastatus and Rhopaloteuthis spp., the CT-images show slight differences in grey scale values (Figs. 3B and 5) suggesting the presence of calcite or sediment (Figs. 3B7‒B12 and 5). The growth increments of the rostrum, are visible largely due to the varying amount of organic matter (Figs. 3B7‒B12 and 5), which has significant lower absorption properties as compared to the calcite. This implies that organic-rich growth increments are darker compared to those composed of calcite (Figs. 3B7‒B12, 5). These darker cavities were probably left behind by the voids created by the infesting parasite during the lifespan of the belemnite. However, the identification of a specific parasite taxon is greatly hampered by the fact that parasites are typically small-bodied (submillimetre) and usually lack biomineralized hardparts. Although, in modern cephalopods, parasites are known from many species from all major oceans ranging from coastal, shelf, to oceanic and deep-sea species (Hochberg 1990; Pascual et al. 1996, 2007; López-González et al. 2000; Gestal et al. 2019), belemnites likely lived in relatively shallow-water shelf seas (Mitchell 2005). Hochberg (1990) documented 200 species of endoparasites in cephalopods with macroparasites such as, nematodes, copepods, and isopods. These predominantly affected non-mineralized tissues such as the digestive tract and gills. The infestation of mainly non-mineralized tissues limits the fossil preservation potential of parasite activity and the resulting host reaction.
The paleopathyforma hamata, as previously documented was predominantly the result of rostrum fracture (Keupp 2012; Radwańska and Radwański 2004, Moosleitner 2006; Keupp et al. 2020) rather than due to parasitism, as noted here (Figs. 4A, 5). This hook-shaped bending compares well with the specimen illustrated by Keupp (2012: fig. 392, top left) as Neoclavibelus sp. from the upper Toarcian of Mistelgau, Upper Franconia (Germany). The present specimen GIUS 8-3798/1 is irregularly swollen throughout and less evenly bent (Fig. 4A) as compared to the Toarcian specimen of Keupp (2012). The other two specimens illustrated by Keupp (2012: fig. 392 top panel; ?Hibolithes sp., Callovian/Oxfordian boundary from the Ogrodzieniec, Poland and ?Hibolithes sp., lower Oxfordian from the Zalas near Kraków, Poland) either have a pronounced J-shaped bent (rostra with bent tips as proposed by Blainville 1827) or caused by rostrum fracture (Radwańska and Radwański 2004; Moosleitner 2006) or display a clear injury-related bending (Keupp 2012: fig. 392; Gonioteuthis sp., lower Campanian of Höver/Lower Saxony). These are distinctly different from the present specimen GIUS 8-3798/2 (Figs. 4A, 5). However, having said that it must also be kept in mind that a swelling or growth defect could potentially also partially cover/erase a previous injury (Kenneth De Baets, personal communication August 2025). However, CT-scan images do not reveal any injury (Fig. 5). Admittedly, although lack of external injury is an important requisite for the inferred paleopathy, but on its own is difficult to assign paleopathologies to parasitism with certainty. It could also reflect other types of diseases; localized swellings and other aspects may suggest that (see also Keupp 2012; De Baets et al. 2021a). However, the fracture of the rostrum or the displaced broken-off rostral fragment that are attached together distinctly shows new calcite lamellae which is absent in the present specimen (Figs. 4A, 5). It must also be kept in mind that the shape of the hook, whether the change in direction occurs distally or proximally, depends largely on the position and ontogenetic time of the injury.
In belemnites, most reported pathologies are due to survival following the attack of predators (e.g., forma angulata), but a few malformations are also assigned to the activity of parasites like forma bullata, collata, and saepia (Keupp 2012); forma hamata is also added herein (Figs. 4A, 5). Although we can identify a parasite infection as the most likely trigger of these pathologies, attributing the deformations to a specific parasite taxon is difficult to impossible. While some parasites leave characteristic patterns on their host, this is not necessarily definitive evidence of their presence. Different parasites can leave similar traces and taxonomically distant parasites can inflict similar symptoms on their hosts because of convergence in the evolution of host-exploitation strategies (Poulin 2011; Leung 2017). Differential diagnosis for paleopathological phenomena may lead to the conclusion that two or more alternative conditions remain as potential triggers (Buikstra et al. 2017; Hoffmann et al. 2018).
Additionally, the lack of soft-tissue remains partly explains the difficulties in identifying a particular culprit or parasite resulting in generalization or an oversimplification (De Baets et al. 2021a). In some instances, clear signs of parasitic remains are present such as organic tube-like structure in ammonoid blister pearls (De Baets et al. 2011), but belong to an unidentifiable (likely extinct) lineage of parasites (De Baets et al. 2021b). In other cases, it is unlikely to expect to find parasitic remains within swellings such as parasitic isopods which cause characteristic swellings in decapods. They are the only group known to cause such characteristic swellings in decapods but they seem to decay faster than their hosts so it would be difficult to find them or rule their presence out just based on the absence of their fossil remains (Klompmaker et al. 2014, 2017; Kenneth De Baets, personal communication August 2025). These swellings as well as pathologies in bivalves caused by trematodes (Huntley et al. 2021) are characteristic enough to be identified as they have modern analogues.
Thus, for now, and pending more collection of similar specimens and their subsequent CT-scans, the bending in the present specimen is assigned to parasitism wherein injury as the causal factor is ruled out (Figs. 4A, 5).
Conclusions
The absence of any signs of external injury in the studied specimens is suggestive of paleopathologies due to parasite infestation. Data based on external features and high-resolution CT-scans on Hibolithes hastatus shows a clear thickening of the rostrum (forma bullata sensu Keupp 2012); the CT-images show differences in grey scale values with darker cavities that were probably left behind by the voids created by the infesting parasite during the lifespan of the belemnite. The specimens of Rhopaloteuthis spp. show two types of paleopathologies with no signs of any external injury; one in which the rostrum is bent (i.e., hook-shaped bending in Keupp 2012) and assigned to forma hamata, and the others in which there is swelling (thickening of the rostrum of Keupp 2012) assigned to forma bullata. The CT-scan of the latter, Rhopaloteuthis spp., also shows no signs of injury. However, attributing the documented deformations to a specific parasitic taxon is difficult to impossible.
Authors’ contributions
SJ carried out the study, participated in the conceptualization and drafted the manuscript. WK and MAS carried out the fieldwork, geological data collection, participated in the fossil preparation and measurements. PD prepared microtomography study. SM participated in drafting and reviewing the manuscript. All authors read and approved the final manuscript.
Acknowledgements
The authors are grateful to the two reviewers, Kenneth De Baets (University of Warsaw, Poland) and Peter Doyle (London South Bank University, UK) for their constructive comments and suggestions that greatly improved the manuscript. The authors are also grateful to Damian Kuźma (Sosnowiec, Poland) for his help during field work.
Editor: Krzysztof Hryniewicz
References
Baumiller, T.K. and Gahn, F.J. 2002. Fossil record of parasitism on marine invertebrates with special emphasis on the platyceratid-crinoid interaction. The Paleontological Society Papers 8: 195–210. Crossref
Blainville, M.H.D. 1827. Mémoire sur les Belemnites. 136 pp. Levrault F.G., Paris.
Buikstra J.E., Cook, D.C., and Bolhofner, K.L. 2017. Introduction: Scientific rigor in paleopathology. International Journal of Paleopathology 19: 80–87. Crossref
Davies, T.G., Rahman, I.A., Lautenschlager, S., Cunningham, J.A., Asher, R.J., Barrett, P.M., Bates, K.T., Bengtson, S., Benson, R.B.J., Boyer, D.M., Braga, J., Bright, J.A., Claessens, L.P.A.M., Cox, P.G., Dong, X.-P., Evans, A.R., Falkingham, P.L., Friedman, M., Garwood, R.J., Goswami, A., Hutchinson, J.R., Jeffery, N.S., Johanson, Z., Lebrun, R., Martínez-Pérez, C., Marugán-Lobón, J., O’Higgins, P.M., Metscher, B., Orliac, M., Rowe, T.B., Rücklin, M., Sánchez-Villagra, M.R., Shubin, N.H., Smith, S.Y., Starck, J.M., Stringer, C., Summers, A.P., Sutton, M.D., Walsh, S.A., Weisbecker, V., Witmer, L.M., Wroe, S., Yin, Z., Rayfield, E.J., and Donoghue, P.C.J. 2017. Open data and digital morphology. Proceedings of the Royal Society B: Biological Sciences 284: 20170194. Crossref
De Baets, K., Hoffmann, R., and Mironenko, A. 2021a. Evolutionary history of cephalopod pathologies linked with parasitism. In: K. De Baets and J.W. Huntley (eds.), The Evolution and Fossil Record of Parasitism: Coevolution and Paleoparasitological Techniques, 203–249. Springer International Publishing, Cham. Crossref
De Baets, K., Huntley, J.W., Klompmaker, A.A., Schiffbauer, J.D., and Muscente, A. D. 2021b. The fossil record of parasitism: its extent and taphonomic constraints. In: K. De Baets and J.W. Huntley (eds.), The Evolution and Fossil Record of Parasitism: Coevolution and Paleoparasitological Techniques, 1–50. Springer International Publishing, Cham. Crossref
De Baets, K., Huntley, J.W., Scarponi, D., Klompmaker, A.A., and Skawina, A. 2021c. Phanerozoic parasitism and marine metazoan diversity: dilution versus amplification. Philosophical Transactions of the Royal Society B 376: 20200366. Crossref
De Baets K., Klug, C., and Korn, D. 2011. Devonian pearls and ammonoid-endoparasite co-evolution. Acta Palaeontologica Polonica 56: 159–180. Crossref
Dera, G., Toumoulin, A., and De Baets, K. 2016. Diversity and morphological evolution of Jurassic belemnites from South Germany. Palaeogeography, Palaeoclimatology, Palaeoecology 457: 80–97. Crossref
Doyle, P. 1990. The British Toarcian (Lower Jurassic) Belemnites. Monograph of the Palaeontological Society London 144: 1–49. Crossref
Doyle, P. and Macdonald, D. 2007. Belemnite Battlefields. Lethaia 26: 65‒80. Crossref
Duval-Jouve, J. 1841. Bélemnites des terrains Crétacés inférieures des environs de Castellane (Basses-Alpes), considérées géologiquement et zoologiquement, avec la description de ces terrains. Académie des Sciences Naturelles de Paris 1841: 1–80.
Finzel, E. 1963. Der Zweispitz-Belemnit von Misburg – ein Unikum. Der Aufschluss 14: 47.
Frerichs, U. 2015. Dokumentation über Belemniten mit Missbildungen aus dem Campan von H. ver und Misburg. Arbeitskreis Paläontologie, Hannover 43: 99–133
Gestal, C., Pascual, S., Guerra, Á., Fiorito, G., and Vieites, J.M. 2019. Handbook of Pathogens and Diseases in Cephalopods. 230 pp. Springer, Cham. Crossref
Głowniak, E. 2012. The perisphinctid genus Prososphinctes Schindewolf (Ammonoidea, subfamily Prososphinctinae nov.): an indicator of palaeoecological changes in the Early Oxfordian Submediterranean sea of southern Poland. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 264: 117–179. Crossref
Hochberg, F.G. 1990. Diseases of Mollusca: Cephalopoda. In: O. Kinne (Ed.), Diseases of Marine Animáis, Vol. 3. Cephalopoda to Urochordata, 47–227. Biologisches Anstalt Helgoland, Hamburg.
Hoffmann, R. and Stevens, K. 2020. The palaeobiology of belemnites—foundation for the interpretation of rostrum geochemistry. Biological Reviews 95: 94–123. Crossref
Hoffmann, R. and Weissmüller, M. 2018. Ein Belemnitenrostrum mit zwei Spitzen aus dem Toarcium (Unterjura) von Dörlbach bei Altdorf. Der Steinkern 34: 38–43.
Hoffmann, R., Ansorge, J., Wesendonk, H., and Stevens, K. 2018. A Late Cretaceous pathological belemnite rostrum with evidence of infection by an endoparasite. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 287: 335–349. Crossref
Hoffmann, R., Schultz, J.A., Schellhorn, R., Rybacki, E., Keupp, H., Gerden, S.R., Lemanis, R., and Zachow, S. 2014. Non-invasive imaging methods applied to neo-and paleo-ontological cephalopod research. Biogeosciences 11: 2721–2739. Crossref
Hoffmann, R., Stevens, K., Picollier, M.-C., Mutterlose, J., and Klug, C. 2020. Non-destructive analysis of pathological belemnite rostra by micro-CT techniques. Acta Palaeontologica Polonica 65: 11‒27. Crossref
Hölder, H. 1973. Auf Fluchtversuch weisende Narben eines Belemniten-Rostrums. Münsterische Forschung zur Geologie und Paläontologie 29: 59–62.
Hüne, L. and Hüne, P. 2008. Des phénomènes paléopathologiques chez une faune de Bélemnites du Callovien supérieur de Bénerville-sur-Mer Calvados, France, Calvados, France. L’Echo des Falaises 12: 67–70.
Hüne, L. and Peter, H. 2008. Des phénomènes paléopathologiques chez une faune de Bélemnites du Callovien supérieur de Bénerville-sur-Mer (Calvados, France). L´Echo des Falaises 12: 67–70.
Huntley, J.W., De Baets, K., Scarponi, D., Linehan, L.C., Epa, Y.R., Jacobs, G.S., and Todd, J.A. 2021. Bivalve mollusks as hosts in the fossil record. In: K. De Baets and J.W. Huntley (eds.), The Evolution and Fossil Record of Parasitism: Coevolution and Paleoparasitological Techniques, 251–287. Springer International Publishing, Cham. Crossref
Jain, S. and Mazur, M. 2021. New dimorphic species of the genus Rollierites Jeannet from southern Poland. Annales Societatis Geologorum Poloniae 91: 405‒418. Crossref
Jain, S. and Salamon, M.A. 2024. First occurrence of the rare middle Jurassic ammonite genus Erymnocerites Jeannet from southern Poland. Historical Biology 36 (4): 742–746. Crossref
Jain, S., Salamon, M.A., and Brachaniec, T. 2022a. On the occurrence of the ammonite aptychi Lamellaptychus cf. rectecostatus (Peters, 1854) from the Ogrodzieniec quarry (southern Poland). Annales de Paléontologie 108: 102534. Crossref
Jain, S., Salamon, M.A., Schweigert, G., and Paszcza, K. 2024. Ammonite-calibrated nautiloid occurrences from the Callovian–Oxfordian (Middle–Upper Jurassic) deposits of southern Poland, Historical Biology 36: 1496–1528. Crossref
Jain, S., Salamon, M.A., Schweigert, G., and Płachno, B.J. 2023. Morphological variations and geographic distribution of the rare Middle Jurassic ammonite Oecoptychius refractus. Acta Palaeontologica Polonica 68: 321–335. Crossref
Jain, S., Salamon, M.A., Schweigert, G., Kuźma, D., and Duda, P. 2022b. Genetic abnormality, parasitism and a new forma type of paleopathy from the upper Callovian ammonite genus Peltoceras Waagen. Lethaia 55: 1–17. Crossref
Jeannet, A., von 1951. Stratigraphie und Palaeontologie des oolithischen Eisenerzlagers von Herznach und seiner Umgebung. Beiträge zur Geologieder Schweiz, Geotechnische Serie 13: 1–240.
Kabanov, G.K. 1967. Belemnite skeleton: morphology and biological analysis [in Russian]. Trudy Paleontologičeskogo Instituta Akademii Nauk SSSR 114: 1–117.
Keupp, H. 2002. Pathologische Belemniten Schein und Wirklichkeit. Fossilien 2002 (2): 85–92.
Keupp, H. 2012. Atlas zur Paläopathologie der Cephalopoden. Berliner paläobiologische Abhandlungen 12: 1–390.
Keupp, H., René, H., and Dirk, F. 2020. Part M, Chapter 20: Pathology of fossil and extant coleoid shells. Treatise Online 135: 1–16. Crossref
Klompmaker, A.A., Artal, P., van Bakel, B.W., Fraaije, R.H., and Jagt, J.W. 2014. Parasites in the fossil record: a Cretaceous fauna with isopod-infested decapod crustaceans, infestation patterns through time, and a new ichnotaxon. PLOS ONE 9 (3), e92551. Crossref
Klompmaker, A.A., Kowalewski, M., Huntley, J.W., and Finnegan, S., 2017. Increase in predator-prey size ratios throughout the Phanerozoic history of marine ecosystems. Science 356: 1178–1180. Crossref
Kraus, W. 2000. Morphologieänderungen von Belemnitenrostren: Phänomene, Ursachen und Bewertung. Mitteilungen zur Ingenieurgeologie und Hydrogeologie 76: 119–126.
Leung, T.L.F. 2017. Fossils of parasites: what can the fossil record tell us about the evolution of parasitism? Biological Reviews of the Cambridge Philosophical Society 92: 410–430. Crossref
López-González, P.J., Bresciani, J., Huys, R., Af, G., Guerra, A., and Pascual, S. 2000. Description of Genesis vulcanoctopusi gen. et sp. nov. (Copepoda: Tisbidae) parasitic on a hydrothermal vent octopod and a reinterpretation of the life cycle of cholidyinid harpacticoids. Cahiers de Biologie Marine 41: 241–253.
Miertzsch, E. 1964. Ein Belemnit mit fünf Spitzen. Der Aufschluss 15: 74.
Mietchen, D., Helmut, K., Bertram, M., and Frank, V. 2005. Non-invasive diagnostics in fossils: Magnetic Resonance Imaging of pathological belemnites. Biogeosciences 2: 133–140. Crossref
Mitchell, S.F. 2005. Eight belemnite biohorizons in the Cenomanian of northwest Europe and their importance. Geological Journal 40: 363–382. Crossref
Montfort, D. de. 1808. Conchyliologie systematique et classification methodique des coquilles. Vol. 1: 408. Paris, F. Schoell.
Moosleitner, G. 2006. Meeresleben in der Unteren Kreide. Die Valanginiummergel im Raum Salérans: Col d´Araud. Fossilien 2006(2): 79–86.
Moosleitner, G. 2012. Fossilien sammeln in der Provence: Die Unterkreide des Plan du Peyron. Fossilien 2012(3): 176–184.
Pascual, S., Gestal, C., Estévez, J.M., Rodríguez, H., Soto, M. Abollo, E., and Arias, C. 1996. Parasites in commercially-exploited cephalopods (Mollusca, Cephalopoda) in Spain: an updated perspective. Aquaculture 142: 1–10. Crossref
Pascual, S., Gonzáez, A., and Guerra, A. 2007. Parasites and cephalopod fisheries uncertainty: Towards a waterfall understanding. Reviews in Fish Biology and Fisheries 17: 139–144. Crossref
Poulin, R. 2011. The many roads to parasitism: a tale of convergence. Advances in Parasitology 74: 1–40. Crossref
Pugaczewska, H. 1961. Belemnoids from the Jurassic of Poland. Acta Palaeontologica Polonica 6: 105‒236.
Radwańska, U. and Radwański, A. 2004. Disease and trauma in Jurassic invertebrate animals of Poland: An updated review. Volumina Jurassica 2: 99–111.
Riegraf, W. 1973. Bißspuren auf jurassischen Belemnitenrostren. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1973 (8): 494–500.
Różycki, S.Z. 1953. Górny dogger i dolny malm Jury Krakowsko-Częstochowskiej. Prace Instytutu Geologicznego 17: 1–412.
Schmid, F. 1963. Ein Nachtrag zum “Zweispitz-Belemnit” aus dem Untercampan von Misburg bei Hannover, pathologische Ausbildung an Gonioteuthis quadrata (Blainville). Der Aufschluss 14: 294–296.
Stevens, G.R. 1965. The Jurassic and Cretaceous belemnites of New Zealand and a review of the Jurassic and Cretaceous belemnites of Indo-Pacific region. New Zealand Geological Survey. Palaeontology Bulletin 36: 1–283.
Stevens, K., Hoffmann, R., Picollier, M.-C., and Mutterlose, J. 2020. Pearl formation in an Early Cretaceous belemnite. Paläontologische Zeitschrift 94: 481–486. Crossref
Wisshak, M., Titschack, J., Kahl, W.-A., and Girod, P. 2017. Classical and new bioerosion trace fossils in Cretaceous belemnite guards characterised via micro-CT. Fossil Record 20: 173‒199. Crossref
Acta Palaeontol. Pol. 70 (4): 755–764, 2025
https://doi.org/10.4202/app.01273.2025