
A new saurodontid fish from the Late Cretaceous of the Western Desert, Egypt
GEBELY A. ABU EL-KHEIR, AHMED MARZOUK, LUIGI CAPASSO, ESSAM ZAHRAN, ATEF M. KASEM, ASMAA KAMEL, MOHAMED K. MOUSA, and ABDELHAMID ELBSEBESHI
Abu El-Kheir, G.A., Marzouk, A., Capasso, L., Zahran, E., Kasem, A.M., Kamel, A., Mousa, M.K., and Elbsebeshi, A. 2026. A new saurodontid fish from the Late Cretaceous of the Western Desert, Egypt. Acta Palaeontologica Polonica 71 (1): 85–94.
A new genus and species of the extinct family Saurodontidae (Ichthyodectiformes) is described from the Upper Cretaceous deposits of Dakhla Formation, Western Desert, Egypt. The specimen is identified as Wadiichthys anbaawyi gen. et sp. nov. The holotype comprises complete skull with maxillae, premaxillae, dentaries and predentary. The specimen was embedded in a compacted mudstone layer of the lower part of Dakhla Formation, Western Desert, Egypt. The new taxon Wadiichthys anbaawyi differs from the others saurodontid genera by (i) the dorsal margin of the predentary is longer than the ventral margin. (ii) the longitudinal foramina below each alveolus of the dentary, premaxilla and maxilla; (iii) a small diastema between the teeth; (iv) a convex contact between the rostrodermethmoid and the frontal; (v) the posterior margin of the maxilla is triangular shaped; (vi) the teeth are more deeply embedded in the premaxilla, maxilla and dentary. Wadiichthys anbaawyi represents the first formal description of a saurodontid from Africa that contributes to the increase of the knowledge of the diversity and the paleogeographic distribution of this family. The new taxon sheds light on the taxonomy, morphology and evolutionary history of the Cretaceous marine vertebrates of the Tethys Sea. To preserve and digitally document the fossil, a terrestrial laser scanner Trimble TX6 was employed. Each of the five fossil parts was scanned separately and processed with Trimble RealWorks software. This created a high-resolution 3D point cloud in true color, grayscale, and intensity mode. This method protects the fossil from possible damage or loss. It also allows for remote collaboration and morphometric analysis without physical handling.
Key words: Actinopterygii, Ichthyodectiformes, Saurodontidae, Dakhla Formation, Cretaceous, Western Desert, Egypt.
Gebely A. Abu El-Kheir [gebely2006@sci.nvu.edu.eg, ORCID: https://orcid.org/0000-0001-5905-8790] and Mohamed K. Mousa [Mohamedkamel.sci@sci.nvu.edu.eg, ORCID: https://orcid.org/ 0000-0002-9951-1905], New Valley Vertebrate Palaeontology Centre, New Valley University, 10th kilometer, Kharga-Assuit road, Kharga, New Valley, Egypt.
Ahmed Marzouk [ahmed.marzouk@sci.dmu.edu.eg, ORCID: https://orcid.org/0009-0006-1111-5163], Essam Zahran [ezahran@sci.dmu.edu.eg], Atef M. Kasem [Kasematef@sci.dmu.edu.eg], and Asmaa Kamel [asmaa.kamel@sci.dmu.edu.eg], Earth Science Department, Faculty of Science, Damanhur University, El Gumhuriya street, Damanhur, Egypt.
Luigi Capasso [l.capasso@unich.it, ORCID: https://orcid.org/0000-0001-6045-3827], University Museum, ‘G. d’Annunzio’ University of Chieti–Pescara, Via dei Vestini 31, 66100 Chieti (CH), Italy.
Abdelhamid Elbsebeshi [abdelhamid_mohammed@nriag.sci.eg, ORCID: https://orcid.org/0009-0004-3616-1516], National Research Institute of Astronomy and Geophysics, Elmarsad street, Helwan, Cairo, Egypt.
Received 2 August 2025, accepted 12 December 2025, published online 18 March 2026.
Copyright © 2026 G.A. Abu El-Kheir et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Ichthyodectiformes an extinct order of actinopterigians, ranging from the Middle Jurassic to the early Paleocene (Boles et al. 2024), and contains species found on nearly all continents (Bardack and Sprinkle 1969). The family Saurodontidae Cope, 1870, are extinct teleosts, first recorded from the Upper Cretaceous, occurring in various deposits and attaining a wide geographical distribution. Within almost 20 genera of the order Ichthyodectiformes, Saurodontidae is more closely related to Gillicus, Xiphactinus, and Ichthyodectes (Maisey 1991). Currently Saurodontidae includes the three genera Saurodon, Saurocephalus, and Prosaurodon, and are all Late Cretaceous in age, ranging from the Cenomanian to the Maastrichtian (Leidy 1860; Cope 1870; Davies 1878; Stewart A. 1898; Bardack and Sprinkle 1969; Stewart J.D. 1999; Taverne and Bronzi 1999; Stewart and Friedman 2001; Alvarado-Ortega et al. 2006).
The Saurodontidae were recorded in the Upper Cretaceous deposits of Western Interior Sea of North America (Fielitz and Irwin 2007), Europe (Taverne and Bronzi 1999), Asia (Kaddumi 2009) and North Africa (Youssef et al. 2011; Sallam et al. 2016). These genera occurred at different time intervals: Saurodon Hays, 1830, was restricted to the Late Coniacian, Saurocephalus Harlan, 1824, first occurred during the early Campanian and continued until the early Paleocene (Boles et al. 2024), and Prosaurodon Stewart, 1999, occurred through Santonian and Campanian (Stewart 1999). The most characteristics feature of this type of fish is the presence of unpaired, edentulous predentary bone.
Cope (1870) established the family Saurodontidae for the two genera, Saurodon and Saurocephalus. Zittel (1888) created the family Saurocephalidae to contain these two genera. Berg (1940), and Bardack and Sprinkle (1969) indicated that Saurodon and Saurocephalus should be placed in the later family, given the priority of Saurocephalus over Saurodon, although the name Saurodontidae given by Cope (1870) is currently used. Currently, Stewart (1999), described Prosaurodon as a more primitive saurodontid fish than both Saurodon and Saurocephalus.
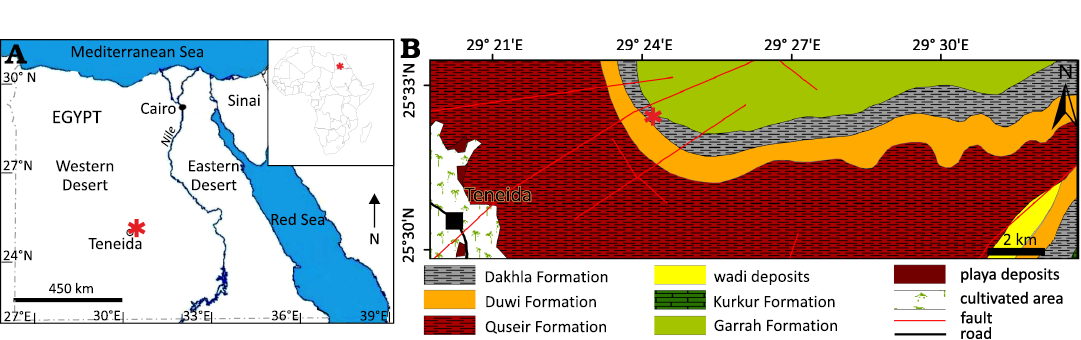
Fig. 1. A. Map showing location of the studied area, northeast of Teneida, Egypt. B. Geological map of Dakhla Oasis. Asterisks indicate fossil locality.
The specimen described in this paper was collected from the lower Maastrichtian deposits of Dakhla Formation, Western Desert of Egypt (Fig. 1). It is represented by a skull, including maxillae, premaxillae, dentaries, and predentary. It is associated with many other remnants of marine vertebrate fossils, such as mosasaurs, turtles and bony fish (GAEK personal observation). The specimen represents a new genus and new species, that is the first genus and species of Saurodontidae described from the African continent. The discovery of a new taxon belonging to the Saurodontidae from the Western Desert of Egypt increases the knowledge of the taxonomy, habitats and the distribution of this family in the Cretaceous period.
Recent advances in three-dimensional imaging and digital documentation are changing vertebrate paleontology. It allows for precise capture, preservation, and worldwide sharing of morphological data from fragile or historically important specimens. High-resolution scanning techniques, like terrestrial laser scanning, create detailed records that protect fossils from damage or loss. Also, promote remote collaboration among researchers. This cuts down on the need for risky transport or repeated handling of delicate materials (Elbshbeshi et al. 2023). For Wadiichthys anbaawyi gen. et sp. nov., these methods have been crucial for detailed documentation. They enable full morphological description, measurement, and phylogenetic analysis without direct physical contact. This is a key advantage for preserving the integrity of such rare and important material.
Institutional abbreviations.—NVP, New Valley Vertebrate Paleontology Center, Kharga city, New Valley Governorate, Egypt.
Nomenclatural acts.— This published work and the nomenclatural acts it contains have been registered in ZooBank: urn:lsid:zoobank.org:pub:6C2B4069-4E06-431F-8446-F75982E17D23.
Geological setting
The stratigraphic section of the study area is represented by the Dakhla Formation (Maastrichtian to lower Paleocene), which is conformably based on the upper most part of the Duwi Formation (Campanian in age), that is formed by the phosphatic fossiliferous layers of limestone and shale (El Azabi and El Arabi 2000). The Dakhla Formation is conformably overlaid by the Tarawan Chalky limestone Formation (Abbass and Habib 1971; Barthel and Herramann-Degen 1981; Tantawy et al. 2001) (Fig. 2). It is formed of about 200 m thickness of intercalation of laminated shale with calcareous fossiliferous siltstone, sandstone and limestone layers. The Dakhla shale facies were deposited on the inner shelf to lagoonal environments. This formation is divided into four members as mentioned in previous studies: Mawhoob, Beris, Lower Kharga, and Upper Kharga (Awad and Ghobrial 1965; Luger 1988; Hewaidy 1997; El-Azabi and El-Araby 2000; Tantawy et al. 2001).
The Mawhoob Member is formed of yellow fossiliferous siltstone with intercalation of laminated shale (Tantawy et al. 2001). The siltstone layer contains elements of vertebrate fossil remains, such as fish bones, mosasaur vertebrae, marine turtles and some shark teeth (Abu El-Kheir et al. 2021).
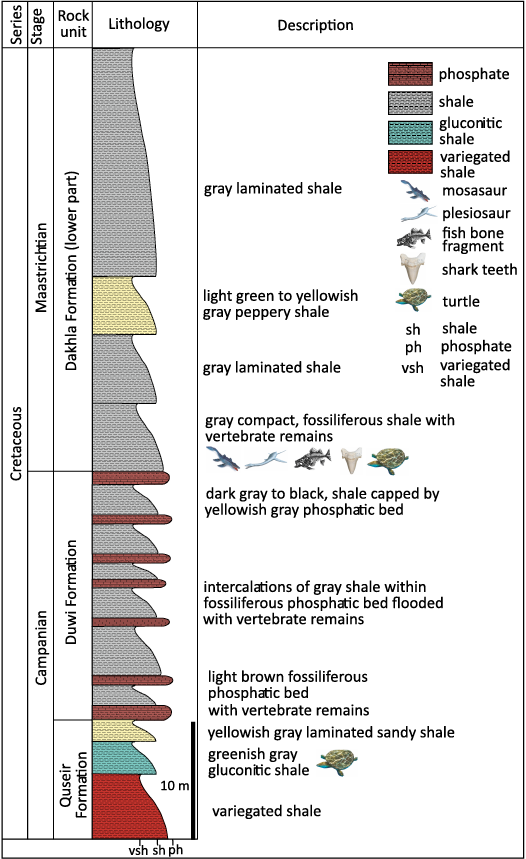
Fig. 2. Stratigraphic column of Teneida section, showing the Quseir, Duwi, and Dakhla formations and location of fossils.
The Beris Member is formed of dark laminated shale with intercalated fossiliferous siltstone layers, this member represents the middle part of the Dakhla Formation. The siltstone layers of this member are known by the abundance of the oyster Exogyra overwegi. The Beris Member contains a variety of marine vertebrate fossil remains, such as partial elements of mosasaurs, plesiosaurs teeth and vertebrae, turtle remains, fishbones, and shark teeth.
The Lower and Upper Kharga members are formed of dark laminated shale with intercalation of calcareous sandstone and limestone layers. These two members are separated by the K/T boundary, which is represented by an erosional surface of the fossiliferous limestone (Abbass and Habib 1971; Barthel and Herramann-Degen 1981; Tantawy et al. 2001). They were deposited in middle to outer continental shelf environment (El Azabi and El Arabi 2000). The present material was collected by the authors during a field trip of the New Valley University Vertebrate Paleontology Centre in February 2024. It was collected from the Mawhoob Member of the Dakhla Formation, north of Teneida village.
Material and methods
The materials described in this paper are housed in New Valley Vertebrate Paleontology center of New Valley University, Kharga Oasis, New Valley governorate, Egypt as NVP028. The specimen is represented by complete left dentary, well preserved robust predentary, two separated left and right maxillae, two separated left and right premaxillae, and highly compacted and squeezed skull.
The described elements were photographed using a digital camera (Af- S Nikkors 24-120 mm 1:4 G ED VR (Nikon) and lense (AF-S Nikkors 70-00 mm 1:28E FL ED VR) in normal light.
To ensure the long-term digital preservation and enable detailed, non-invasive analysis of the Wadiichthys anbaawyi gen et sp. nov. holotype (NVP028), a uniquely complete yet fragile saurodontid specimen, the five main cranial parts (maxillae, premaxillae, dentaries, predentary, and the compressed skull) were each scanned using a TX6 Trimble terrestrial laser scanner. This precise instrument was chosen to capture complex surface shapes and fine details, including tooth rows, foramina, and sutural contacts, which are crucial for systematic and evolutionary studies (Soliman et al. 2025). Each fossil part was scanned from multiple overlapping angles to ensure full coverage and reduce data gaps caused by the occlusion or specimen curvature, a common issue in vertebrate paleontology. The resulting high-density point clouds preserve not only the three-dimensional shape of each element but also surface color and intensity data, allowing for better visualization of anatomical features that may be hidden in traditional photographs.
All raw scan data were imported into
Trimble RealWorks software for processing 3D point clouds, for initial
registration, cleaning, and noise reduction. During registration,
individual scans were aligned precisely using both automated reference
systems, ensuring submillimeter accuracy and a fully
integrated digital model (Elbshbeshi et al. 2022).
Next, automated and manual cleaning tools were used to eliminate
false points caused by scanner artifacts, matrix debris,
or lighting issues, producing refined, high-resolution point clouds for
each specimen part. The processed datasets were exported in various
formats, including true-color and grayscale point clouds, as well as
intensity-based renders, to meet different analytical and
visualization needs. This digital workflow not only creates a strong
archival record that protects the specimen from future degradation or
loss but also allows for global access and collaborative research
without the risks of physical transport or handling. Furthermore, these
3D models enable accurate morphological measurements, virtual
dissection, and comparative anatomical studies entirely in the digital
space, making them valuable tools for both current research and future
discoveries in the systematics and paleobiogeography of Cretaceous
marine vertebrates.
Source data available from the online repository Zenodo
(https://doi.org/10.5281/zenodo.18784034)
Systematic paleontology
Class Osteichthyes Huxley, 1880
Subclass Actinopterygii Klein, 1885 (sensu Goodrich, 1930)
Superorder Teleosteomorpha Arratia et al., 2004
Order Ichthyodectiformes Bardack & Sprinkle, 1969
Family Saurodontidae Cope, 1870
Genus Wadiichthys nov.
Zoobank LSID: lsid:zoobank.org:act:2E1FEF12-EE7A-4C4A-A1AA-B65C2BD058CA.
Type species: Wadiichthys anbaawyi gen. et sp. nov., see below.
Etymology: A combination of wadi, valley; referring to New Valley Governorate, the locality of the studied specimen and Greek ichthys, fish.
Diagnosis.—As for the monotypic type species.
Stratigraphic and geographic range.—As for the monotypic type species.
Wadiichthys anbaawyi gen. et sp. nov.
Figs. 3–6.
Zoobank LSID: lsid:zoobank.org:act:BD27B6D7-45F5-4360-8E47-96EA14E2AFD5.
Etymology: In honor of Mohammed Ibraheem El Anbaawy, the professor and the spiritual father of the first author.
Holotype: NVP028, complete left dentary, well preserved robust predentary, two separated left and right maxillae, two separated left and right premaxillae, and highly compacted and squeezed skull.
Type locality: North of Teneida village, northeast of Mut city, Dakhla Oasis, New Valley Governorate, Egypt. GPS coordinates deposited at NVP and available to authorized researchers.
Type horizon: Biozone CF7 (Hewaidy et al. 2017), base Mawhoob Member, Dakhla Formation, Maastrichtian.
Material.—Type material only.
Diagnosis.—Wadiichthys anbaawyi gen. et sp. nov. is diagnosed by the following combination of characters: (i) the dorsal margin of the predentary is longer than the ventral margin, (ii) the longitudinal foramina below each alveoli of tooth of the dentary, premaxilla and maxilla, (iii) presence of a small diastema between the teeth, (iv) the contact between the rostrodermethmoid and frontal is convex, (v) the maxilla has a triangular posterior margin, (vi) the teeth are distinctly embedded in the maxilla and dentary.
Description.—Skull: The skull is nearly complete, but highly compacted and squeezed due to the compaction of the hosting mudstone layer (Fig. 3). The rostrodermethmoid is a laterally compressed relatively small bone, and represents the most anterior bone of the skull distinguished by an anterolaterally thick, narrow and short projected process. It extends anteriorly and may slightly cleft in the median plane. Posterior to this process the rostrodermethmoid forms a pair of laterally projecting processes varying in size. The rostrodermethmoid is articulated posteriorly with the frontal through a curved contact, vomer ventrolaterally, and ethmopalatines dorsolaterally.
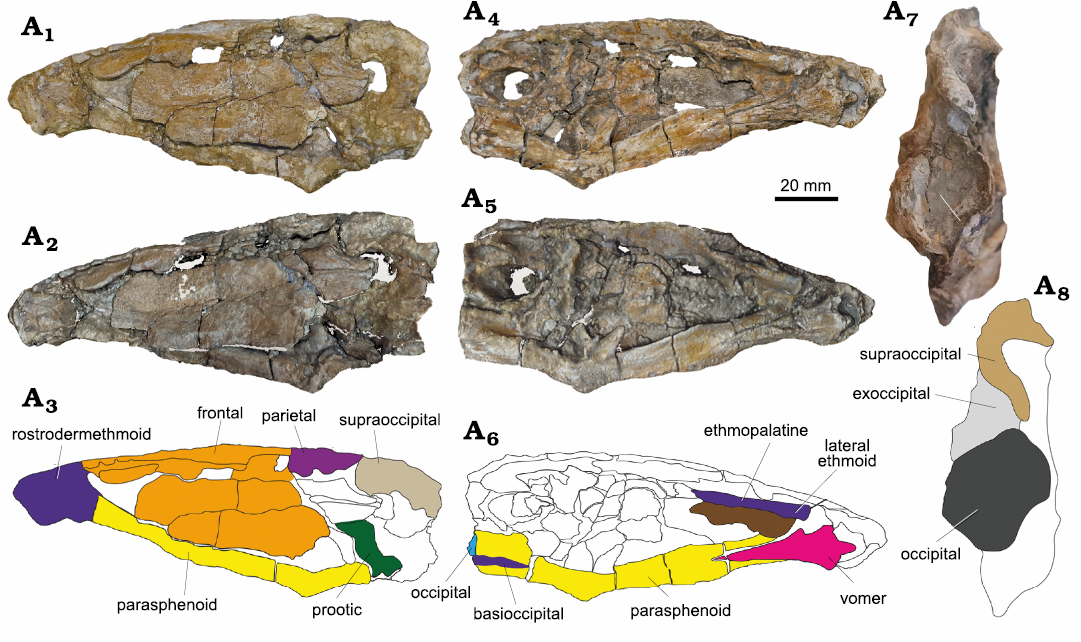
Fig. 3. Saurodontid fish Wadiichthys anbaawyi gen. et sp. nov. (NVP028), north Teneida village, Northeast Mut city, Dakhla Oasis, Egypt, Maastrichtian. Skull in left (A1–A3) and right (A4–A6) lateral views and posterior view of the skull (A7, A8), showing the occipital, exoccipital and supraocipital bones. Photograph (A1, A4), laser scanning (A2, A5, A7), and explanatory drawing (A3, A6, A8).
The ethmopalatine is characterized by a lateral stout elongate thick process that extends from the posterior end of the rostrodermethmoid with a slightly curved stout head to the anterior end of the orbit region. The ethmopalatine bone contacts the rostrodermethmoid anteriorly, frontal dorsally, vomer and parasphenoid ventrally.
The lateral ethmoid is a semi triangular robust bone, contacts the ethmopalatine dorsally, rostrodermethmoid anteriorly and vomer and parasphenoid ventrally
The frontal is a long broad bone above the orbital region of the skull. It is highly affected by the compaction of the hosting mudstone layer and moved laterally closing the orbital region. It contacts with the rostrodermethmoid anteriorly, ethmopalatine anterolaterally, parietals posteriorly through a suture. The frontal is dorsally thick, split by a median line of the skull, and laterally extended above the orbital region with thick shield-like bone.
The parietal is a short thick bone, it contacts the frontal anteriorly, supraoccipital posteriorly. The lateral parts of the parietal are completely removed.
The supraoccipital is a thin compact bone. Its crest is well preserved and extended ventrally to the occipital. It contacts the parietal anteriorly, and the rest of the exoccipital posteroventrally. The occipital bone has a rounded to an oval shallow groove for the articulation with the first vertebra. It is bounded by exoccipital laterally, supraoccipital dorsally, and basioccipital ventrally. The epioccipital bone is broken.
The exoccipital form the lateral margin of the occipital. It meets the supraoccipital dorsally, basioccipital ventrally, and the rest of the prootic anteriorly.
The basioccipital is a rounded and concave bone. It is laterally compressed, forming the posteroventral section of the skull. It contacts the parasphenoid ventrally, and the rest of the exoccipital dorsally.
The parasphenoid is a longitudinal flattened bar shaped bone, covering the most of the ventral side of the skull. It contacts the basioccipital posteriorly, and the lateral ethmoid anterodorsally. It is bifurcated anteriorly into two divergent arms by the posterior tapered bar extending of the vomer. Also, it bifurcated again below the basioccipital, and the two arms extended almost to the posterior end of the basioccipital. The parasphenoid is bent at the posteroventral corner of the orbit, and forms an obtuse angle. The parasphenoid has a triangular-shaped at the posteroventral corner of the orbit, and extends anterolaterally to form the ventral border of the orbit, but has a flat-shaped below the otic region, and extends posterolaterally to complete the ventral border of the otic region.
The vomer is an elongated dagger shaped bone. It is wide anteriorly and tapered posteriorly. It contacts the lateral ethmoid anteriorly, and posteriorly and laterally the parasphenoid. Two well preserved left and right maxillae, containing damaged and preserved small sharp teeth. The maxilla is a robust bone element, labiolingually compressed and dorsoventerally extended (Fig. 4). The maxilla has a slightly convex oral margin, containing a single original teeth row alternated with remnants of replacement teeth beneath the marginal teeth in the lingual side.
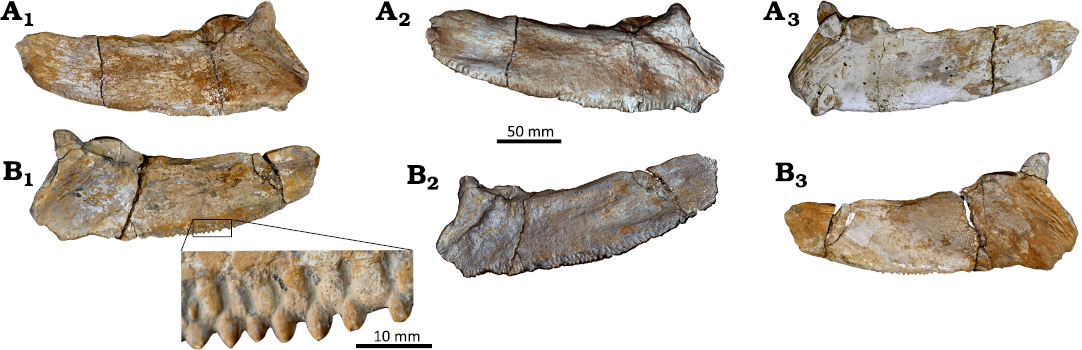
Fig. 4. Saurodontid fish Wadiichthys anbaawyi gen. et sp. nov. (NVP028), north Teneida village, Northeast Mut city, DakhlaOasis, Egypt, Maastrichtian. A. Left maxilla in lingual (A1, A2) and labial (A3) views. B. Right maxilla in labial (B3) and lingual (B1, B2) views, showing the longitudinal foraminae (enlargement), located directly above the row of the teeth. Photograph (A1, A3, B1, B3), laser scanning (A2, B2).
The anterior margin exhibits oblique triangular shape, it is anterolingually directed with rough labial face serving the articular with the premaxilla. The anterior margin is higher than the posterior margin, and both exhibit convex surfaces. The tips of the original teeth of the maxilla are almost broken and missed except some of them which appeared as lanceolate shaped. The teeth are labiolingually compressed and extended vertically. They have lingual and labial biconvex surfaces with sharp and smooth bicarinate and sharp apex. The teeth are circular in cross section and tightly packed. The replacement teeth are originated aside below the original teeth, keeping the same morphological characters as the original teeth. Distinctive series of longitudinal foramina, located directly above the row of the teeth. The foramina are parallel to each other exhibiting comb shaped.
The premaxilla is a semi rectangular bone with flat dorsal margin (Fig. 5) The oral margin is strongly convex, while the symphyseal margin is slightly straight to concave. The oral and symphyseal margins intersect, forming a narrow anterior end. The symphyseal margin has a medial bend that produces a convex labial face. The labial surface is slightly convex, but lingual surface is flat. The anterior one third of the premaxilla is thicker than the posterior part, containing one row of densely packed teeth. The teeth are similar to those of the maxilla, they are embedded inside the premaxilla, with longitudinal foramina below the row of the teeth .The posterior two third of the premaxilla in the lingual surface is thin and have rough surface for the articulation with the anterior end of the maxilla.

Fig. 5. Saurodontid fish Wadiichthys anbaawyi gen. et sp. nov. (NVP028), north Teneida village, Northeast Mut city, Dakhla Oasis, Egypt, Maastrichtian. . A. Left premaxilla in lingual (A1) and labial (A2) views. B. Right premaxilla in lingual (B1) and labial (B2) views.
The dentary is a straight robust, triangular shaped bone (Fig. 6A). It extends anteroposteriorly and labiolingually compressed. It has straight oral and ventral margins that converge anteriorly forming a narrow and straight symphyseal margin. The symphyseal margin meets the oral margin at a nearly right angle. The oral margin has a single row of densely embedded packed teeth. Most of the teeth have broken and the preserved teeth exhibit lanceolate shape. The teeth extended vertically, and have anterior and posterior smooth carinae and sharp apex. They are triangular shaped, and gradually increase in size from the anterior to the posterior end of the dentary. A series of distinctive faint longitudinal foramina is located directly below the alveoli of the teeth in the lingual side. The lingual side has a deep elongated medial triangular depression that intersects with an anteriorly deep oval depression.
The predentary is a semi triangular shaped, and slender ventrally. The dorsal and ventral margins are straight and unequal (Fig. 6B1). The dorsal margin is longer, wider and more robust than the ventral margin. The dorsal and ventral margins meet medially, forming an anterior tapered point. The length of the dorsal margin is roughly one and half the length of the posterior margin, while the length of the ventral margin is 1.4 the length of the posterior margin.
The posterior margin is an oval shaped (Fig. 6B2), with blunt dorsal and pointed ventral edges. About four articular facets and mid keel in the posterior margin for articulation with the dentaries.

Fig. 6. Saurodontid fish Wadiichthys anbaawyi gen. et sp. nov. (NVP028), north Teneida village, Northeast Mut city, Dakhla Oasis, Egypt, Maastrichtian. A. Left dentary in lingual (A1, A2) and labial (A3, A4) views. Photograph (A1, A3), laser scanning (A2, A4). B. Predentary in lateral (B1) and posterior (B2) views.
Remarks.—The described specimen was embedded in hard and compact mudstone of the basal part of Mawhoob Member, Dakhla Formation. The specimen elements are highly affected by compaction and by numerous cracks caused by cycles of wetting and drying of the mudstone. Some elements are fragmented and abraded, affecting by the wind action and the weathering factors. The skull is highly compressed and squeezed due to the compaction of the host mudstone layer. The left side of the frontal is missing and the right side is compressed laterally closed the orbital opening. The other bones of the skull are highly lateral compressed.
Stratigraphic and geographic range.—Type locality and horizon only.
Discussion
The specimen NVP028 may be referred to the family Saurodontidae, based on the dentary that extends beyond the upper jaw and tipped by unpaired edentulous predentary bone (Bardack and Sprinkle 1969), blade like teeth arranged in a single row along the dentary and maxilla, elongated jaws, the structure of the mandibular symphysis at the front meet and that contact with the predentary (Bardack 1965; Stewart 1999). The specimen is characterized by diagnostic features that allow it to be partially differentiated from Saurocephalus lanciformis Harlan, 1834, and Saurodon leanus Hays, 1830, as following:
(i) the dorsal and ventral margins of the predentary are unequal in length as in Saurocephalus lanciformis, but in Saurodon leanus they are equal in length. The dorsal margin is longer than the ventral margin, but in Saurocephalus lanciformis the ventral margin is longer than the dorsal margin (Fig. 7);

Fig. 7. Comparison of predentaries in saurodontid fishes. A. Saurodon leanus (Hays, 1830) EPC 1993-23, Gove County, Kanzas, USA; Santonian. B. Saurocephalus lanciformis (Harlan, 1824) RMDRC 10-033, Logan County, Kanzas, USA; Early Campanian. C. Wadiichthys anbaawyi gen. et sp. nov. (NVP028), north Teneida village, Northeast Mut city, Dakhla Oasis, Egypt, Maastrichtian.
(ii) a distinctive line of shallow vertical longitudinal groove just below the alveoli in the lingual border of the maxilla and the dentary;
(iii) the teeth are more embedded in the maxilla and dentary than in Saurocephalus lanciformis and Saurodon leanus;
(iv) the teeth are vertically directed as in Saurocephalus lanciformis, differing than those of Saurodon leanus, which are directed anteriorly;
(v) there is a short diastema between each tooth, which is differentiated than in Saurocephalus lanciformis, where the teeth are located very close to each other;
(vi) the anterior margin of the maxilla is triangular shaped as in Saurocephalus lanciformis, but in Saurodon leanus is rounded;
(vii) the antero-dorsal flange of the anterior margin is straight as in Saurodon leanus, more differentiated than in Saurocephalus lanciformis, which is shallow concave;
(viii) the dorsal margin of the maxilla as in Saurocephalus lanciformis is more concave than in Saurodon leanus;
(ix) the premaxilla is subrectangular as in Saurocephalus lanciformis, differing than in Saurodon leanus, which rhomboidal;
(x) the dorsal margin of the premaxilla is rounded as in Saurodon leanus, differing in that respect from Saurocephalus lanciformis, where premaxilla is straight;
(xi) the symphyseal margin of the premaxilla is shallow concave, differing than in Saurocephalus lanciformis, which is convex, and Saurodon leanus, which is straight;
(xii) the oral margin of the premaxilla is straight, while in Saurocephalus lanciformis and Saurodon leanus it is convex;
(xiii) the contact between the rostrodermethmoid and the frontal is curved similar to the Egyptian specimen (mentioned in the published poster by Youssef et al. 2011), unlike both Saurodon leanus, which has a zigzag suture, and Saurocephalus lanciformis, which is zigzag and curved, Saurocephalus longicorpus Kaddumi, 2009, which is more pointed and extended, resulted in bifurcating of the rostrodermethmoid. Also, the rostrodermethmoid is relatively larger than in the Egyptian specimen (Youssef et al. 2011), but smaller than in Saurodon leanus, Saurocephalus lanciformis, and Saurocephalus longicorpus (Fig. 8).

Fig. 8. Comparison of skull in saurodontid fishes. A. Saurocephalus
longicropus (Kaddumi, 2009). B. Saurocephalus lanciformis (Harlan, 1824).
C. Saurodon
leanus (Hays, 1830). D.
Egyptian specimen (Youssef et al. 2011). E.
Wadiichthys anbaawyi sp. nov. Note
different types of contact between rostrodermethmoid and frontal.
Dorsal (A–C), lateral (D, E) views.
The application of high-resolution terrestrial laser scanning (TX6) to the Wadiichthys anbaawyi holotype provided precise three-dimensional models that were fundamental to the anatomical interpretation presented in this study. The resulting point cloud data captured subtle morphological details, including foramina distribution, tooth-row configuration, and complex bone sutures, which are difficult to document reliably using conventional photography or manual illustration. These high-fidelity models enabled detailed measurement, virtual restoration, and morphometric comparison without physical handling of the fragile specimen, thereby minimizing the risk of damage.
Importantly, the TLS-derived models supported accurate visualization of features affected by breakage and sediment compaction, allowing clearer assessment of diagnostic characters relevant to saurodontid taxonomy. In addition, the creation of permanent digital models ensures long-term preservation of the holotype and facilitates reproducibility, comparative studies, and future re-evaluation.
Biogeography.—The Saurodontidae is an extinct family of the order Ichthyodectiformes. They are known from marine deposits of North America, Asia, Europe and North Africa, indicating their wide biogeographic distribution through the Cretaceous epicontinental seas.
The genera Saurodon and Saurocephalus were recorded in the Western Interior Seaway of the North America, Europe and Asia, showing their cosmopolitan nature.
The saurodontid Wadiicthys anbawyii gen. et sp. nov. from the Maastrichtian Dakhla Formation, Western Desert, Egypt reported herein, represents the first formal description of a saurodontid genus and species from Africa.
The new discovery not only provides the wide distribution of the saurodontid, but also their abilities for inhibiting the southern margin of the Tethyan Sea.
The record of a saurodontid fish in Egypt reflects the high significance of Dakhla Formation as well Late Cretaceous vertebrate bearing sites in Egypt. Moreover, it also documents the routes of faunal migrations between the present day North America, Eurasia and Africa during this period.
Generally, the wide distribution of the saurodontid reflects their abilities for adaptation within different Late Cretaceous marine ecosystems.
Evolutionary implications.—The diagnostic cranial and dental features of Wadiichthys such as the proportions of the predentary margins, the observed diastema between the teeth and the shape of the foramina below the teeth, illustrate new morphological characteristics that are not previously recognized in this family. This provides that the saurodontid were more evolutionary flexible and could adapt to different environments.
The record of Wadiichthys in Africa, far from the North American and Eurasian occurrences, shows that saurodontid achieved wide geographical distribution, allowed by extensive marine connections of the Cretaceous Tethys Sea. This wide distribution supports the faunal exchange between the continents that allowed evolutionary adaptations to the variety of the marine environments. It also fills the gap in the geographic distribution of the family and helps in reconstructing its phylogeny.
Conclusions
Many vertebrate fossils remains, such as marine reptiles and fish bones, have been recorded in the Late Cretaceous deposits of the Dakhla Formation, Western Desert, Egypt. Saurodontid fish remains have been previously recorded in the Late Cretaceous deposits of the Late Campanian Duwi Formation, including the Egyptian specimen (Youssef et al. 2011).
The newly described specimen posesses diagnostic features, which allow to describe a new genus and species of Wadiichthys anbaawyi within the family Saurodontidae. Summarizing, the characteristic features of the new taxon are: (i) the contact between the rostrodermethmoid and the frontal is convex; (ii) the ratio between the length of dorsal margin to the ventral margin of the predentary, where the dorsal margin is longer than the ventral margin; (iii) the presence of diastema between the teeth; (iv) the shape of the foramina of premaxillae, maxillae, and dentaries are longitudinal; (v) the posterior margin of the maxilla is triangular shaped; (vi) the teeth are more deeply embedded in the premaxilla, maxilla and dentary.
Acknowledgments
The authors thank very much the New Valley University (Kharga Oasis, Egypt), especially Abdel Aziz Tantaawy for funding the field trips as well as preparation of the specimens in their laboratory. Additionally, the authors would like to thank Salama Ali Mohammed (the Head of Balat city, Egypt) and his team for their hospitality and great support during the field trips. Great thanks are extended to Essam Yousef (New Valley University, Kharga Oasis, Egypt) for his great efforts in collecting fossils and Ali Mohamed Radwan (Geodynamic Department, National Research Institute of Astronomy and Geophysics, Cairo, Egypt) for appreciated efforts Laser Scan documentation. Many thanks go to Heba Farahat Mohammed, the restoration technician of the New Valley Vertebrate paleontology center for her great efforts in the restoration of the studied specimen. We deeply thank the reviewers Diogo de Mayrinck (Rio de Janeiro State University, Brazil) and Jesús Alvarado-Ortega (Departamento de Paleontología, Instituto de Geología, National Autonomous University of Mexico, Mexico City, Mexico) for their valuable and constructive insights, which significantly improved the manuscript’s quality. Furthermore, a great gratitude to Editor Camila Cupello for the professional guidance and dedication throughout the revision process.
Editor: Camila Cupello
References
Abbass, H.L. and Habib, M.M. 1971. Stratigraphy of the west Mawhoob area, south Western Desert, Egypt. Bulletin Institute Desert Egypt 19: 47–107.
Abu El-Kheir, G.-M., Abdel Gawad, M.K., and Kassab, W.G. 2021. First known gigantic sea turtle from the Maastrichtian deposits in Egypt. Acta Palaeontologica Polonica 66: 349–355. Crossref
Alvarado-Ortega, J., Blanco-Piñón, A., and Porras Múzquiz, H. 2006. Primer registro de Saurodon (Teleostei: Ichthyodectiformes) en la cantera La Mula, Formación Eagle Ford (Cretácico Superior: Turoniano), Múzquiz, Estado de Coahuila, México. Revista Mexicana de Ciencias Geológicas 23: 107–112.
Arratia, G., Scasso, R.A., and Kiessling, W. 2004. Late Jurassic fishes from Longing Gap, Antarctic Peninsula. Journal of Vertebrate Paleontology 24 (1): 41–55. Crossref
Awad, G.H and Ghobrial, M.G. 1965. Zonal stratigraphy of the Kharga Oasis. Geological Survey Egypt 34: 1–77.
Bardack, D. 1965. Anatomy and evolution of chirocentrid fishes. University of Kansas, Paleontological Contributions, Vertebrata 10: 1–87.
Bardack, D. and Sprinkle, G. 1969. Morphology and relationships of saurocephalid fishes. Fieldiana Geology 16: 297–340. Crossref
Barthel, K.W. and Herrmann-Degen, W. 1981. Late Cretaceous and Early Tertiary stratigraphy in the Great Sand Sea and its south-eastern margins (Farafra and Dakhla Oases), south Western Desert, Egypt. Mitteilungen Bayerische Staatssammlung Palaontologie Historische Geologie 21: 141–182.
Berg, L.S. 1940. Classification of fishes both recent and fossil. Trudy Zoologicheskogo Instituta Akademia Nauk SSSR 5: 87–517. [In Russian with English translation].
Boles, Z.M., Ullmann, P.V., Putnam, I., Ford, M., and Deckhut, J.T. 2024. New vertebrate microfossils expand the diversity of the chondrichthyan and actinopterygian fauna of the Maastrichtian–Danian Hornerstown Formation in New Jersey. Acta Palaeontologica Polonica 69: 173–198. Crossref
Cope, E.D. 1870. On the Saurodontidae. Proceedings of the American Philosophical Society 11: 529–538.
Cuvier, G. 1817. Le régne animal; 2. Les reptiles, les poissons, les mol lusques, et les annélides. 523 pp. Chez Déterville, Paris.
Cuvier, G. and Valenciennes, A. 1846. Histoire Naturelle des Poissons, Vol. 19, 150–168. Pitois-Levrault, Paris.
Davies, W. 1878. On the nomenclature of Saurocephalus lanciformis of the British Cretaceous deposits: with description of a new species (S. woodwardi). Geological Magazine 5: 254–261. Crossref
El-Azabi, M.H. and El-Araby, A. 2000. Depositional cycles: an approach to the sequence stratigraphy of Dakhla Formation, west Dakhla-Farafra stretch, Western Desert, Egypt. Journal of African Earth Science 30: 971–996. Crossref
Elbshbeshi, A., Gomaa, A., Mohamed, A., Othman, A., and Ghazala, H. 2022. Seismic hazard evaluation by employing microtremor measurements for Abu Simbel area, Aswan, Egypt. Journal of African Earth Sciences 196: 104734 Crossref
Elbshbeshi, A., Gomaa, A., Mohamed, A., Othman, A., Ibraheem, I.M., and Ghazala, H. Applying geomatics techniques for documenting heritage buildings in Aswan Region, Egypt: A case study of the Temple of Abu Simbel. Heritage 2023: 742–761. Crossref
Fielitz, C. and Irwin, K. 2007. Records of ichthyodectiform fishes from the Upper Cretaceous (Campanian) of Arkansas. Journal of Vertebrate Paleontology 27 (3, supplement): 74A.
Goodrich, E.S. 1930. Studies on the Structure and Development of Vertebrates. 837 pp. Macmillan, London. Crossref
Harlan, R. 1824. On a new fossil of genus of the order Enalio Sauri (of Conybeare). Journal of the Academy of Natural Sciences of Philadelphia 3: 331–337.
Hay, O.P. 1898. Notes on species of Ichthyodectes, including the new species I. cruentus, and on the related and herein established genus Gillicus. American Journal of Science, Series 4 6: 225–232. Crossref
Hays, I. 1830. Description of a fragment of the head of a new fossil animal, discovered in a marl pit, near Moorestown, New Jersey. Transactions of the American Philosophical Society, Series 2 3: 471–477. Crossref
Hewaidy, A.A. 1997. A proposed palaeoecologic scheme for the Upper Cretaceous–Lower Tertiary sequences in Egypt. Middle East Research Centre, Ain Shams University, Earth Science Series 11: 159–168.
Hewaidy, A.A., Farouk, S., and Bazeen, Y.S. 2017. Sequence stratigraphy of the Maastrichtian–Paleocene succession at the Dakhla Oasis, Western Desert, Egypt. African Earth Sciences 136: 22–43. Crossref
Huxley, T.H. 1880. On the application of the laws of evolution to the arrangement of the Vertebrata, and more particularly of the Mammalia. Proceedings of the Zoological Society of London 1880: 649–662.
Kaddumi, H.F. 2009. Saurodontids (Ichthyodectiformes: Saurocephalus lanciformis) of Harrana with a description of a new species from the late Maastrichtian Muwaqqar Chalk Marl Formation. In: H.F. Kaddumi, Fossils of the Harrana Fauna and the Adjacent Areas, 215–231. Eternal River Museum of Natural History, Amman.
Leidy, J. 1860. Remarks on Saurocephalus and its allies. Transactions of the American Philosophical Society (new series) 11: 91–95. Crossref
Leidy, J. 1870. Remarks on ichthyodorulites and on certain fossil Mammalia. Proceeding of Academy of Natural Sciences of Philadelphia 22: 12–13.
Luger, P. 1988. Campanian to Palaeocene agglutinated foraminifera from freshwater influenced marginal marine (deltaic) sediments of southern Egypt. Geologische Bundesanstalt 41: 255–263.
Maisey, J.G. 1991. Santana Fossils: An Illustrated Atlas. 459 pp. TFH Publications, Neptune City.
Newton, E.T. 1878. Remarks on Saurocephalus lanciformis, and on the species which have been referred to this genus. Geological Society of London, Quarterly Journal 34: 786–796. Crossref
Sallam, H.M., O’Connor, P.M., Kora, M., Sertich, J.J.W., Seiffert, E.R., Faris, M., Ouda, K., El-Dawoudi, I., Saber, S., and El-Sayed, S. 2016. Vertebrate paleontological exploration of the Upper Cretaceous succession in the Dakhla and Kharga Oases, Western Desert, Egypt. Journal of African Earth Sciences 117: 223–234. Crossref
Soliman, M., Ali, D., Elbshbeshi, A., and Ibrahim, M. 2025. Digitization of Qaitbay Fort in Alexandria (884AH/1479CE). Simplification of modelling techniques to safeguard vulnerable cultural heritage. Art Research Journal 25: 109–121.
Stewart, A. 1898. Some notes on the genus Saurodon and allied species. Kansas University Quarterly 7: 177–186.
Stewart, J.D. 1999. A new genus of Saurodontidae (Teleostei: Ichthyodectiformes) from Upper Cretaceous rocks of the Western Interior of North America. In: A. Arratia and H.-P. Schultze (eds.), Mesozoic Fishes. Systematics and Fossil Record, 335–360. Dr Friedrich Pfeil, München.
Stewart, J.D. and Friedman, V. 2001. Oldest North American record of Saurodontidae (Teleostei: Ichthyodectiformes). Journal of Vertebrate Paleontology 21 (3, supplement): 104A.
Tantawy, A.A., Keller, G., Addatte, T., Stinnesbeck, W., Kassab, A., and Schulte, P. 2001. Maastrichtian to Paleocene depositional environment of the Dakhla Formation, Western Desert, Egypt: sedimentology, mineralogy, and integrated micro-and macrofossil biostratigraphies. Cretaceous Research 22: 795–827. Crossref
Taverne, L. and Bronzi, P. 1999. Les poissons crétacés de Nardò. Note complémentaire sur le Saurodontinae (Teleostei, Ichthyodectiformes): Saurodon elongatus, sp. nov. Studie Ricerchesui Giacimenti Terziari di Bolca 8: 105–116.
Youssef, M., Sallam, H., Friedman, M., O’Connor, P., and Sertich, J. 2011. A saurodontid fish from the Late Cretaceous of Dakhla Oasis, Western Desert, Egypt. Journal of Vertebrate Paleontology 31 (Conference Supplement): 218.
Zittel, K.A. 1888. Handbuch der Paläontologie, Palaeozologie, Band III, Vertebrata (Pisces, Amphibians, Reptilia, Aves). 924 pp. R. Oldenbourg, München.
Acta Palaeontol. Pol. 71 (1): 85–94, 2026
http://dx.doi.org/10.4202/app.01277.2025