
A Paleogene chondrichthyan assemblage from central Chile supports a latitudinal and temporal boundary of the Weddellian Province along the southeastern Pacific
RODRIGO A. OTERO, SERGIO SOTO-ACUÑA, RAÚL UGALDE P., and HÉCTOR ORTIZ
Otero, R.A. Soto-Acuña, S., Ugalde P., R., and Ortiz, H. 2026. A Paleogene chondrichthyan assemblage from central Chile supports a latitudinal and temporal boundary of the Weddellian Province along the southeastern Pacific. Acta Palaeontologica Polonica 71 (2): 255–266.
The locality of Algarrobo in central Chile, has yielded a rich Late Cretaceous fossil assemblage that enriched the knowledge of the vertebrate diversity in the context of the Weddellian Biogeographic Province, by providing mid-latitude records comparable to coeval taxa found in subantarctic paleolatitudes. In this sense, Algarrobo represents the northernmost Late Cretaceous available locality that can be included as part of the Weddellian Biogeographic Province. Before this research, post-Cretaceous vertebrate records from Algarrobo were limited to the mention of chondrichthyan teeth which remained undescribed, without detailed stratigraphic provenance, nor additional context. Unusual high-energy tide conditions during 2024 caused the removal of most algae that permanently cover the fossiliferous sediments exposed in the coastal platform. This allowed the recovery of the first Paleogene chondrichthyan assemblage from Algarrobo. Available material includes Abdounia beaugei, aff. Carcharhinus sp., Cretolamna sp., Xiphodolamia sp., Striatolamia macrota, Carcharias sp., Rhinoptera sp., and Sulcidens sulcidens. The combined biochron of these taxa supports the occurrence of Ypresian levels in the studied unit, extending the age interval of the fossil-bearing informal unit (Algarrobo Beds) to the lower–upper Eocene, thus, implying a hiatus of ca. 12–14 Ma with respect to the Upper Cretaceous underlying levels. The paleogeographic distribution of the recognized taxa shows affinities with Europe and north Africa. This interchange reinforces the hypothesis of a late Paleogene chondrichthyan turnover along the southeastern Pacific, based on upper Paleocene–lower Oligocene assemblages recognized in southern localities of the Pacific margin. The studied fauna complements an emerging picture for the temporal and geographical declination of the Weddellian Province, likely beginning during the late Paleocene, and acquiring a more marked Atlantic influence during the Eocene–Oligocene.
Key words: Chondrichthyes, biogeography, Paleogene, Weddellian Province, Chile, South America.
Rodrigo A. Otero [otero2112@gmail.com; ORCID: https://orcid.org/0000-0002-3046-2973 ], Red Paleontológica U-Chile, Laboratorio de Ontogenia y Filogenia, Departamento de Biología, Facultad de Ciencias, Universidad de Chile, Las Palmeras 3425, Ñuñoa, Santiago, Chile; Nucleus Early Evolutionary Transitions of Mammals, ANID, Chile; Museo de Historia Natural y Cultural del Desierto de Atacama. Interior Parque El Loa s/n, Calama, Chile.
Sergio Soto-Acuña [sesotacu@ug.uchile.cl; ORCID: https://orcid.org/0000-0002-9311-9355 ], Paleontológica Red U-Chile, Laboratorio de Ontogenia y Filogenia, Departamento de Biología, Facultad de Ciencias, Universidad de Chile, Las Palmeras 3425, Ñuñoa, Santiago, Chile; Nucleus Early Evolutionary Transitions of Mammals, ANID Chile; Escuela de Medicina Veterinaria, Pontificia Universidad Católica de Chile. Benito Rebolledo 2054, Macul, Santiago, Chile; Kaytreng Consultores en Paleontología SpA. Ñuñoa, Santiago, Chile.
Raúl Ugalde P. [raul.ugalde@umayor.cl; ORCID: https://orcid.org/0000-0002-8085-8909 ], Nucleus Early Evolutionary Transitions of Mammals, ANID Chile; Kaytreng Consultores en Paleontología SpA. Ñuñoa, Santiago, Chile; Escuela de Geología, Facultad de Ciencias, Ingeniería y Tecnología, Universidad Mayor, Camino La Pirámide 5750, Huechuraba, Santiago, Chile; PEDECIBA Geociencias, Facultad de Ciencias, Universidad de la República, Iguá 4225, Montevideo, Uruguay.
Héctor Ortiz [hctros@gmail.com; ORCID: https://orcid.org/0000-0001-6010-3990 ], Red Paleontológica U-Chile, Laboratorio de Ontogenia y Filogenia, Departamento de Biología, Facultad de Ciencias, Universidad de Chile; Las Palmeras 3425, Ñuñoa, Santiago, Chile; Nucleus Early Evolutionary Transitions of Mammals, ANID, Chile.
Received 11 October 2025, accepted 21 March 2026, published online 3 June 2026.
Copyright © 2026 R.A. Otero et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
The Weddellian Biogeographic Province (WBP) is a concept that reunites several austral coeval localities as part of an equivalent biogeographic realm extended along mid latitudes (33–65°S) of the southern hemisphere, including Antarctica, southern South America and New Zealand, in a time span between the Late Cretaceous–Eocene (Zinsmeister 1979). The WBP was initially based on the taxonomical affinities noted among invertebrate assemblages, although, this was later shown to occur also among marine vertebrates (Reguero et al. 2012).
Within this provincial context, chondrichthyan assemblages from the southeastern Pacific have been mostly documented in Upper Cretaceous units (Philippi 1887; Wilckens 1904; Wetzel 1930; Oliver-Schneider 1936; Gasparini 1979; Suárez et al. 2003; Suárez 2015; Otero 2024). In the last decades, growing evidence of Paleogene chondrichthyans has been documented in several localities of central Chile (Muñoz-Ramírez et al. 2007; Groz and Palma-Heldt 2013; Fernández-Jiménez et al. 2018; Rodríguez et al. 2023), allowing a more complete picture on the local evolution of the group along this realm.
In particular, the locality of Algarrobo (33°21’S; 71°40’W) has yielded the richest Late Cretaceous (early Maastrichtian) chondrichthyan diversity known in the southeastern Pacific (Philippi 1887; Brüggen 1915; Tavera 1980; Suárez et al. 2003; Suárez 2015; Bogan et al. 2016; Otero 2019b, 2024). On the contrary, the Paleogene chondrichthyan record from Algarrobo remained poorly known, comprising to date the sole mention of material referred to as indeterminate myliobatids from putative middle–upper Eocene levels (Suárez and Marquardt 2003; Suárez 2015).
This contribution presents new local chondrichthyan remains recently recovered during 2024, which were naturally exposed due to unusual tide conditions that washed up the fossil-bearing levels. The studied diversity helps to fill a chronostratigraphic and latitudinal gap in the austral chondrichthyan fossil record. The implications of the new studied diversity studied are commented, pointing to a moment and latitudinal segment for the declination of the WBP.
Institutional abbreviations.—MUSA.P.VERT., Colección Paleontológica, Museo de Historia Natural e Histórico de San Antonio, Valparaíso Region, Chile.
Geological setting
Algarrobo is a coastal touristic town settled 120 km W from Santiago, in central Chile (Fig. 1A). Along its shoreline, two marine units crop out, mostly exposed during low tides. These units are the Estratos de la Quebrada Municipalidad (Quebrada Municipalidad Beds; Gana et al. 1996), of early Maastrichtian age based on biostratigraphy (Brüggen 1915; Tavera 1980) and a single Sr/Sr stable isotope data (Suárez and Marquardt 2003). The second unit is the Estratos de Algarrobo (“Algarrobo Beds”; Gana et al. 1996). The existence of post-Cretaceous levels in Algarrobo was noted by Brüggen (1915) who distinguished two different sedimentary units in the coast of Algarrobo, one of them of “Tertiary” age based on its invertebrate fauna. The latter author also provided the first approach to the spatial distribution of fossiliferous beds in a sketched but detailed geological map with sections Later, Tavera (1980) studied the invertebrate assemblage, constraining the unit to the middle–upper Eocene, and moreover, correlating it to the Millongue Formation (Tavera 1942). Gana et al. (1996) referred to these levels as the Algarrobo Beds, describing them as variably coarse, very fossiliferous sandstones with abundant concretions, reaching ca. 95 m thickness, and overlying through an erosive discordant contact to the lower Maastrichtian older strata. The age of the Algarrobo Beds was mostly based on the combined biochron of the invertebrate assemblage studied by Tavera (1980). Their outcrops are intermittently exposed during low tides, but these are mostly covered by brown and green algae rendering the fossil sampling difficult.
The unit (Fig. 1B) represents a shallow marine to coastal environments, even reaching the backshore, as evidenced by subordinated coquinas in the middle portion of the analysed section. The direct supply of continental and terrigenous materials, as tree trunks or organic matter, suggest a proximity to a drainage mouth.
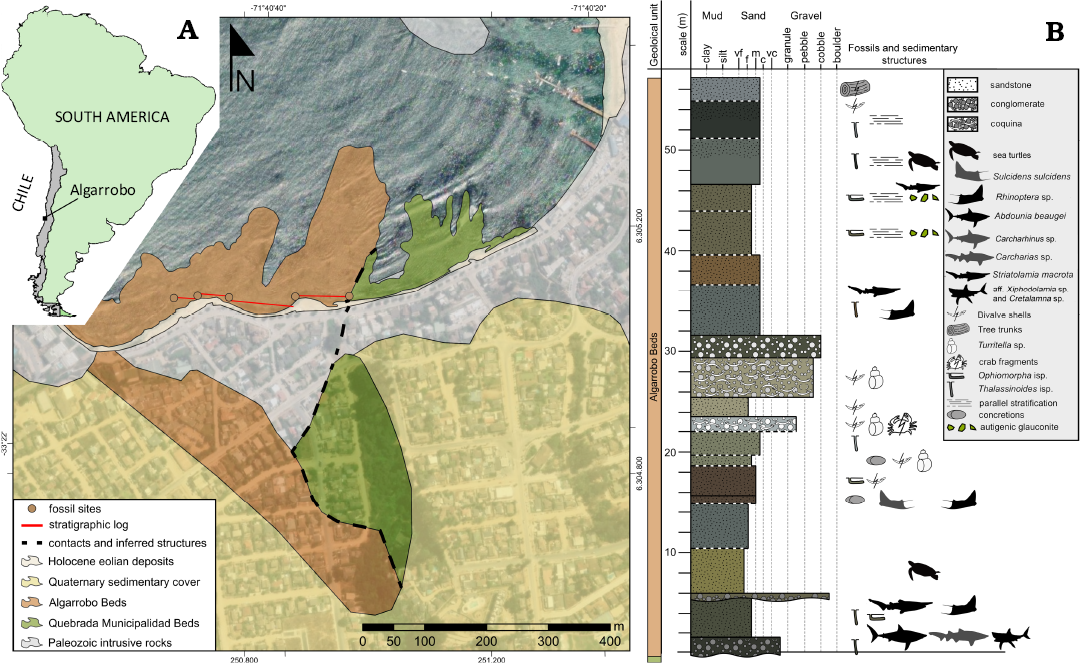
Fig. 1. A. Map of Algarrobo, central Chile, with detail of the sedimentary units in its coast. There are exposed the Quebrada Municipalidad Beds and the Algarrobo Beds, with the main trace of the sedimentary log. Base satellite image from ArcGIS Pro 3.4 repository. The cartography composes previous large scale geological maps, satellite photointerpretation and field data. B. Scheme of the studied section, complemented with the previous collected vertebrate materials. The total thickness reach ~57 m, comprising the lower two thirds of the unit.
Material and methods
The studied material was collected in a single day. During austral winter 2024, exceptional high-energy tides affected the coast of Algarrobo. These events caused high erosion in the sedimentary outcrops but also eroded the dense covering of algae (Ulva spp.). This event allowed the recognition of an unusually high number of in situ chondrichthyan teeth, as well as remains of other vertebrates currently under study. The early detection of this situation was notified by inhabitants of Algarrobo, motivating a quick action for recovering these samples under imminent risk of loss due to the ongoing high tides. In a single day of field work during July 2024, the samples studied were recovered together with their stratigraphic context. The preparation of the specimens considered sharp dental tools for mechanical removal of the hard matrix and cleaning with isopropyl alcohol for removing humidity.
Despite the limited sampling time, the collected specimens were recovered from several different horizons, representing an interesting diversity with only few taxonomically redundant specimens. However, the reduced sampling time should be considered limited and poorly representative of the expected diversity present in the studied section. Due to this, the collected specimens represent a first approach to chondrichthyans and they allow the first comparisons between the known subantarctic assemblages and those recovered in lower latitudes of the Pacific Ocean.
Assessing reworked or transported material considered preservation aspects as worn surfaces, loss of lateral cusplets, loss of sharp cutting edges, and consistent abrasion over the crown and root of each tooth. Among the recovered samples, three teeth show abraded crowns and roots, however, these also show a mineralization, color and general aspect identical to those observed in well-preserved teeth. Moreover, the chronostratigraphic distribution of the taxa represented by abraded teeth does not conflict with that of well-preserved samples (see Discussion). Under these criteria, coeval transportation is interpreted as a primary driver for the abraded teeth instead of reworking from older strata.
The stratigraphic section was measured during June 2025, being complemented with the registered occurrences of the fossil material studied here. A measure tape was used to directly record the apparent thickness. After several and progressive measurements of the dip and strike of the levels with a Brunton Compass, the effective dimension of each stratum in the unit was calculated. The exposure is also cut by the marine ravine surface, in assessing its disposition. For each level, the fresh rock colour was taken using a Munsell Rock Color chart. By visual inspection and magnifiers, the grain sizes were noted, as well as sedimentary structures. Later, the sedimentary log was digitalized using Sedlog v3.1 and InkScape v1.2.
Systematic palaeontology
Chondrichthyes Huxley, 1880
Elasmobranchii Bonaparte, 1838
Neoselachii Compagno, 1977
Lamniformes Berg, 1958
Mitsukurinidae Jordan, 1898
Genus Striatolamia Glikman, 1964
Type species: Otodus macrotus Agassiz, 1843; Eocene, Paris Basin, France.
Striatolamia macrota (Agassiz, 1843)
Fig. 2A–C.
Material.—MUSA.P.VERT.1752, anterior tooth from lowest part of the Algarrobo Beds basal conglomerate, briefly above the inferred K/Pg erosive conformable contact; MUSA.P.VERT.1764, large lateral tooth from upper levels of the Algarrobo Beds, middle–upper Eocene; MUSA.P.VERT.1756, small lateral tooth from sandstone level overlying the basal conglomerate, Ypresian.
Description.—Anterior tooth with high and slender cusp, slightly sigmoidal in profile. A single, small, sharp and medially recurved cusplet is present on each flank of the crown. The cusplet has a smooth labial face and a lingual face with many strong striations. The root lobes are elongated and divergent. Lateral teeth possess high triangular cups, with a single, crenulated cusplet on each side. There are striations over the crown lingual face, but these are comparatively softer than those present in anterior teeth. The root lobes are divergent but less extended than in anterior teeth.

Fig. 2. Lamniformes and Carcharhiniformes from the Ypresian (lower Eocene) of Algarrobo Beds, Algarrobo, central Chile. A–C. Striatolamia macrota (Agassiz, 1843). A. MUSA.P.VERT.1752, anterior tooth in labial (A1), profile (A2), and lingual (A3) views. B. MUSA.P.VERT.1764, lateral tooth in labial (B1) and lingual (B2) views. C. MUSA.P.VERT.1756, lateral tooth in labial (C1) and lingual (C2) views. D. Carcharias sp., MUSA.P.VERT.1753, lateral tooth in labial (D1 and lingual (D2) views. E. aff. Xiphodolamia sp. MUSA.P.VERT.1754, lateral tooth in labial (E1) and lingual (E2) views. F. Cretolamna sp. MUSA.P.VERT.1757, lateral tooth in labial (F1) and lingual (F2) views. G. aff. Carcharhinus sp., MUSA.P.VERT.1765, lateral tooth in labial (G1) and lingual (G2) views. H–J. Abdounia beaugei (Arambourg, 1935). H. MUSA.P.VERT.1750a, posterolateral tooth in labial (H1) and lingual (H2) views. I. MUSA.P.VERT.1750b, lateral tooth in labial (I1), profile (I2), and lingual (I3) views. J. MUSA.P.VERT.1750c, lateral tooth in labial (J1), profile (J2), and lingual (J3) views. Scale bar 10 mm.
Remarks.—In the Austral (= Magallanes) Basin, Striatolamia teeth have been found in association to teeth of the genus Carcharias (Otero et al. 2012, 2013; Otero and Soto-Acuña 2015). However, the presence of Carcharias in higher latitudes of Antarctica during the Eocene remains unreported (see diversity summary in Kriwet 2005 and Reguero et al. 2012). Teeth of Striatolamia can be distinguished from those of Carcharias by the presence of strong folds in the lingual face and a comparatively less recurved profile, especially in anterior teeth (Cappetta 1987, 2012; Cunningham 2000). According to Cappetta (2012), the species Striatolamia macrota can be recognized by the marked reduction of the lateral cusplets in anterior teeth (a feature present in MUSA.P.VERT.1752) and by the pectinated aspect of the cusplets in lateral teeth (as the posterior cusplet present in MUSA.P.VERT.1756 and 1764). Following these traits and considering the widespread Weddellian occurrences of Striatolamia macrota (Welton and Zinsmeister 1980; Long 1992; Reguero et al. 2012; Otero et al. 2012, 2013; Otero and Soto-Acuña 2015; and likely in Rodríguez et al. 2023), the current records are here referred to this species.
Odontaspididae Müller & Henle, 1839
Genus Carcharias Rafinesque, 1810
Type species: Carcharias taurus Rafinesque, 1810; Recent, Mediterranean.
Carcharias sp.
Fig. 2D.
Material.—MUSA.P.VERT.1753, slightly worn lateral tooth from lowest part of the Algarrobo Beds basal conglomerate, briefly above the K/Pg erosive conformable contact, Ypresian.
Description.—The available lateral tooth possesses a slender, sigmoidal cusp with a single, triangular cusplet on each flank. Root has divergent lobes with one of them broken. The labial and lingual faces are both smooth, without striations.
Remarks.—Teeth of Carcharias are the most common vertebrate remains in the Upper Cretaceous deposits of central Chile (Suárez et al. 2003; Suárez 2015; Otero 2024; Cañete et al. 2025). During the late Paleocene, the genus was also reported in central Chile (Muñoz-Ramírez et al. 2007; Groz and Palma-Heldt 2013; Fernández-Jiménez et al. 2016; Rodríguez et al. 2023), proving its local continuous abundance throughout the K/Pg boundary and onwards. In the Austral Basin, Carcharias teeth have been recognized in several Eocene localities, being represented by a large number of samples (Otero et al. 2012, 2013; Otero and Soto-Acuña 2015). These are also abundant in the Paleocene of New Zealand (Mannering and Hiller 2008). On the contrary, Paleogene occurrences of the genus Carcharias in Antarctica remain dubious. Most of the previously referred material has been reassigned in later studies (e.g., “Carcharias” macrota in Long 1992, belongs to Striatolamia macrota).
The unique available specimen of this genus has a crown and root surfaces abraded. but mineralization and general preservation is much similar to most recovered samples, thus, suggesting transportation.
Xiphodolamiidae Glikman, 1964
Genus Xiphodolamia Leidy, 1877
Type species: Xiphodolamia ensis Leidy, 1877; Eocene, New Jersey, USA.
aff. Xiphodolamia sp.
Fig. 2E
Material.—MUSA.P.VERT.1754, lateral tooth from sandstone level overlying the basal conglomerate, Ypresian.
Description.—Tooth with distinctive low and broad triangular cusp, without lateral cusplets. Its cutting edge is smooth. The root is squared, without divergent lobes. The tooth is labiolingually compressed.
Remarks.—The previous features plus its size under 3 cm coincide with the diagnostic traits for the genus Xiphodolamia (see Cappetta 2012). However, this referral must be kept with precaution, considering the previous report of other lamnids in the Paleogene of central Chile, particularly, the species Macrorhizodus praecursor (Leriche, 1905) that was preliminary reported by Otero (2015). The posterior teeth of this taxon could acquire a remarkably similar morphology to that of MUSA.P.VERT.1754 during the growth of lateral teeth. On the contrary, mature lateral teeth of Macrorhizodus praecursor are noticeably larger. Moreover, the available tooth shows clear signs of abrasion, indicative of transportation. MUSA.P.VERT.1754 is conferred to the genus Xiphodolamia by the moment, awaiting additional specimens from Algarrobo which could eventually confirm its taxonomical affiliation. Previous known records of Xiphodolamia are restricted to the lower Eocene–upper Eocene of North America, Europe, north and west Africa, and India (Cappetta 2012).
Cretoxyrhinidae Glikman, 1958
Genus Cretolamna Glikman, 1958
Type species: Otodus appendiculatus Agassiz, 1838; Turonian, England, UK.
Cretolamna sp.
Fig. 2F.
Material.—MUSA.P.VERT.1757, one lateral tooth from basal conglomerate of the Algarrobo Beds unit, Ypresian.
Description.—Small tooth lacking its anterior cusplet and a fragment of the root, with triangular, posteriorly recurved crown having complete, smooth cutting edges. The posterior cusplet is broad, triangular and blunt, with complete, smooth cutting edges, having a basal constriction in its contact with the root, and clearly separated from the crown. The root is squared, lingually prominent, and lacking divergent lobes.
Remarks.—The presence of a squared (rectangular) root is a trait commonly present in several advanced lamnids (e.g., Lamna nasus, Carcharodon carcharias) but otherwise, indistinctly present among lateral teeth of few Paleogene lamnids (e.g., “Isurolamna spp.”, Lethenia spp., Macrorhizodus praecursor, Xiphodolamia spp., Cretolamna spp.). However, the presence of a single, broad and triangular cusplet in lateral teeth is informative. This trait is absent in the genera “Isurolamna”, Lethenia, Macrorhizodus, and Xiphodolamia, but indeed occurs in Cretolamna (= Cretalamna, following the opinion of Cappetta 2012), although, in the latter genus more than one cusplet is commonly present on lateral teeth.
Remarkably similar teeth referred to this genus have been reported in the southern hemisphere, particularly in the upper Campanian of Antarctica (“Cretalamna” in Otero et al. 2014: fig. 3). Material referred to “Cretalamna” appendiculata was also described from Maastrichtian units of Argentinean Patagonia (Bogan and Agnolín 2010). The current new record represents the first documented occurrence of Cretolamna in the southeastern Pacific and its youngest austral occurrence.
Carcharhiniformes Compagno, 1973
Carcharhinidae Jordan & Evermann, 1896
Genus Carcharhinus de Blainville, 1816
Type species: Carcharias melanopterus Quoy & Gaimard, 1824; Recent off Waigeo Island.
aff. Carcharhinus sp.
Fig. 2G.
Material.—MUSA.P.VERT.1765, lateral tooth from upper levels of the Algarrobo Beds, middle–upper Eocene.
Description.—Tooth with high and narrow cusp strongly recurved posteriorly, having a flat labial face and a convex lingual face. The cusp has smooth cutting edges and is separated from the mesial and distal heels. Both heels have a rough serration. The root lobes are divergent and medially separated by a nutritious groove. The medial part of the root extends occlusally beyond the heels level.
Remarks.—The lack of additional teeth of the same type precludes by the moment assuring a generic affiliation. However, the presence of smooth cutting edges in teeth of Carcharinus was noted by Cappetta (2012) to occur among pre-Neogene representatives. On the contrary, serrated cutting edges are known to occur in all Neogene species (Cappetta 2012: 301). The genus Carcharhinus can be tracked back to the Lutetian (Cappetta 2012). The current record could potentially represent the first occurrence of Carcharhinus in the Paleogene of the south Pacific Ocean.
Genus Abdounia Cappetta, 1980
Type species: Eugaleus beaugei Arambourg, 1935; Ypresian, Morocco.
Abdounia beaugei (Arambourg, 1935)
Fig. 2H–J.
Material.—MUSA.P.VERT.1750a, posterolateral tooth; MUSA.P.VERT.1750b; MUSA.P.VERT.1750c, d, two lateral teeth. All from basal conglomerate level of the Algarrobo Beds, Ypresian.
Description.—Small teeth characterized by the presence of a triangular and sharp crown moderately broad on its base, flanked by one or more triangular cusplets which are uninterrupted with the crown enameloid, having complete, smooth cutting edges. The crown has a flat labial face with basal folds, and a convex lingual surface, while the root has two divergent branches and a lingual medial nutritious groove.
Remarks.—All the available specimens from the Algarrobo Beds are indistinguishable from Abdounia beaugei, characterized by two or mostly three triangular, straight lateral cusplets (Cappetta 1980: fig. 4; Cappetta 2012: fig. 291). Austral records of the genus Abdounia were previously known in two upper Eocene localities of the Magallanes Basin on southernmost Chile (Otero et al. 2012; 2013) and in the Ypresian of Antarctica (Engelbrecht et al. 2017; Charnelli et al. 2024), being represented by A. mesetae and A. richteri (Engelbrecht et al. 2017). Moreover, the material from southernmost Chile likely belongs to these same latter species. On the contrary, the current record from the Algarrobo Beds represents the first Ypresian occurrence of Abdounia in the Weddellian Province and the first record of the species Abdounia beaugei in this realm.
Batomorphii Cappetta, 1980
Myliobatiformes Compagno, 1973
Genus Rhinoptera Cuvier, 1829
Type species: Myliobatis marginata Geoffroy-Saint-Hilaire, 1817; Recent, Mediterranean.
Rhinoptera sp.
Fig. 3A–C.
Material.—MUSA.P.VERT.1758a, abraded symphyseal tooth from sandstone level immediately above the basal conglomerate of the Algarrobo Beds; MUSA.P.VERT.1758b, c, two incomplete symphyseal tooth from 30 m (surface) from the top of the basal conglomerate, mid-levels of the Algarrobo Beds. All from Ypresian.
Description.—Symphyseal teeth with soft surface and hexagonal occlusal contour, much broader than axially long. Its root is characterized by an alternation of furrows and laminae of very similar breadth.

Fig. 3. Myliobatiformes from the Ypresian (lower Eocene) of Estratos de Algarrobo, Algarrobo, central Chile. A–C. Rhinoptera sp. A. MUSA.P.VERT.1758a, worn symphyseal tooth in occlusal (A1) and basal (A2) views. B. MUSA.P.VERT.1758c, symphyseal tooth in axial (B1), occlusal (B2), and profile (B3) views. C. MUSA.P.VERT.1758b, damaged symphyseal tooth in the matrix, in axial (C1), occlusal (C2), and profile (C3) views. D, E. Sulcidens sulcidens (Dartevelle & Casier, 1943). D. MUSA.P.VERT.1751a, symphyseal tooth in occlusal (D1), basal (D2), axial (D3), and profile (D4) views. E. MUSA.P.VERT.1751b, associated symphyseal tooth in occlusal (E1), basal (E2), axial (E3), and profile (E4) views. Scale bar 10 mm.
Remarks.—Dental plates of indeterminate Myliobatiformes from the Paleogene of Algarrobo have been reported previously, but these were not figured nor described (Suárez and Marquardt 2003; Suárez 2015). In addition, coeval records of Rhinoptera have been described in the Biobío Region of central Chile (Muñoz-Ramírez et al. 2007) with additional records mentioned by Groz and Palma-Heldt (2013). Further austral records of the genus Rhinoptera are known from Ypresian levels of Antarctica (Charnelli et al. 2024).
Incertae sedis
Genus Sulcidens Underwood et al., 2019
Type species: Myliobatis sulcidens Dartevelle & Casier, 1943; Paleocene?, Angola.
Sulcidens sulcidens (Dartevelle & Casier, 1943)
Fig. 3D, E.
Material.—MUSA.P.VERT.1751a, b, two associated symphyseal teeth from mid-levels of the Algarrobo Beds, Ypresian.
Description.—Symphyseal teeth having a slight concave occlusal surface. The latter also has axial parallel sulci and a very fine granulose surface. The root base has alternated furrows and laminae, the latter being almost twice broader than the furrows.
Remarks.—Symphyseal teeth of Sulcidens sulcidens can be recognized by their concave occlusal surface having axial parallel sulci and possessing a very fine granulose surface rarely removed by in life wear (Underwood et al. 2017). Ahead this research, this species has been exclusively recorded in the Paleocene of Angola (Dartevelle and Casier 1943) and in the Paleocene and Ypresian, lower Eocene of Morocco (Arambourg 1952; Underwood et al. 2017). The current material represents its first austral record.
Discussion
Age of the studied section.—The occurrence of Sulcidens sulcidens in the Algarrobo Beds suggests the presence of upper Paleocene–lower Eocene levels in the unit. Previous known records of this species were restricted to the Selandian–Ypresian of north Africa (Underwood et al. 2017 and historical references therein). In addition, the known biochron of Cretolamna is restricted to the Albian–Ypresian (Cappetta 2012), implying a fossil-bearing level age not younger than the early Eocene. The eventual occurrence of Xiphodolamia is also informative although this must be considered with precaution due to the possible transportation of the unique available specimen. This genus was restricted to the Ypresian–Priabonian of Europe, North America, west Indies, north and west Africa, Near East, and central Asia (Cappetta 2012). Adnet et al. (2009) commented the eventual presence of Xiphodolamia in the Danian of Kazakhstan (based on previous records referred to Eoxyphodolamia mangislakensis by Glikman 1980). Moreover, records of Abdounia beaugei are restricted to the Thanetian–lower Ypresian of north Africa and the Lutetian of southern France (Arambourg 1935; Cappetta 1980, 2012; Boulemia and Adnet 2023). On the contrary, known South American records of Abdounia are restricted to the upper Eocene of southernmost Chile (Otero et al. 2012, 2013; Cañete et al. 2025), from geologic units that are part of the former Austral (=Magallanes) Basin that opened into the Atlantic realm. Finally, the local presence of a single specimen referred to aff. Carcharhinus in upper levels of the Algarrobo Beds, complements the chronostratigraphic interpretation, considering that the latter genus is known since the Lutetian and onwards (Cappetta 2012), thus, reinforcing an expectable age younger than the Ypresian for the upper part of the Algarrobo Beds. The combined biochron of the recognized taxa supports a lower age closer to the Thanetian/Ypresian boundary for the studied section (Table 1). Based on the new studied chondrichthyan assemblage, the Algarrobo Beds sediments extend at least from the lower Eocene to the upper Eocene, while its lower section between the basal conglomerate until the Sulcidens sulcidens fossil-bearing level (ca. 35 m on surface), is no younger than the Ypresian. The upper boundary of the Algarrobo Beds belongs to the middle–upper Eocene, based on the biostratigraphy of the invertebrate assemblage (after Brüggen 1915; Tavera 1980; Gana et al. 1996).
Table 1. Biochrons of each identified taxon (in dark grey) recovered from the Algarrobo Beds and described here. Records with tentative biochron extension are indicated in light grey. The current age interpretation relies on well-preserved teeth without signs of transportation. Otherwise, unique taxon samples with signs of abrasion are marked with an asterisk (in the case of Rhinoptera sp., only a single tooth shows abrasion, while the remaining samples have good preservation).
|
Taxon |
Maastrichtian |
Danian |
Selandian |
Thanetian |
Ypresian |
Lutetian |
Bartonian |
Priabonian |
Rupelian |
Chattian |
References |
|
aff. Xiphodolamia sp.* |
|
|
|
|
|
|
|
|
|
|
|
|
Cretolamna sp. |
|
|
|
|
|
|
|
|
|
|
|
|
Abdounia beaugei |
|
|
|
|
|
|
|
|
|
|
|
|
Carcharias sp.* |
|
|
|
|
|
|
|
|
|
|
|
|
Striatolamia macrota |
|
|
|
|
|
|
|
|
|
|
|
|
Rhinoptera sp. |
|
|
|
|
|
|
|
|
|
|
|
|
Sulcidens sulcidens |
|
|
|
|
|
|
|
|
|
|
|
|
aff. Carcharhinus sp. |
|
|
|
|
|
|
|
|
|
|
Taphonomic remarks.—The lowest part of the Algarrobo Beds basal conglomerate has frequent in situ teeth of carcharhinids (i.e., Abdounia beaugei) associated with scarcer in situ material referred to Cretolamna sp. On the other hand, the upper part of this conglomerate includes in situ material referred to Striatolamia macrota, as well as likely transported teeth referred to aff. Xiphodolamia sp. and Rhinoptera sp. Extant carcharhinids are the dominant sharks in all warm and temperate seas, inhabiting tropical continental coastal and offshore waters, coral reefs and oceanic islands while a few are truly oceanic and range far into the great ocean basins ( ). On the contrary, extant species of Rhinoptera are characterized to inhabit tropical to warm temperate coastal waters up to 100 m depth, preferring shallow bays, lagoons and estuaries (Ebert and Stehmann 2013). The frequent presence of Rhinoptera (and secondarily, Sulcidens) in the overlying levels, adds to frequent driftwood remains, gastropods banks (“Turritella” landbecki), decapods, Ophiomorpha isp. and Thalassinoides isp. (Tavera 1980; Schweitzer et al. 2006) directly associated in some cases, reinforcing the evidence of shallow water environment with fluvial influence. All these taxa and their stratigraphic occurrences suggest a second or third order regressive local condition during the accumulation of the studied unit.
Paleogeography.—Before this study, the species Abdounia beaugei and Sulcidens sulcidens were both restricted to North Africa and southern Europe (Cappetta 2012). The new studied records are the first documented in the southern hemisphere and in the Pacific realm. In addition, the previously known occurrences of Xiphodolamia were restricted to several localities in the northern hemisphere, occurring now in the southeastern Pacific. Moreover, the Eocene occurrences of Carcharhinus were previously described in north Africa and North America (Cappetta 2012).
The studied material adds to an emerging picture for a direct interchange with the northern Tethys, as previously noted by the presence of blochiid fishes both in the Tethys and the southeastern Pacific (Fierstine and Monsch 2002; Otero 2019a). Previous records of Palaeogaleus, Physogaleus, and Premontreia in central Chile (Rodríguez et al. 2023) adds extra support for a direct connection with the North Atlantic, considering that these three genera were previously known in Europe, North America, north/west Africa, and the Near East (Cappetta 2012). In addition, the recent description of Lethenia carranzaensis in the Eocene–Oligocene boundary of central Chile (Otero 2025), complements this scenario, considering that the genus Lethenia was previously restricted to the lower Oligocene of Belgium (Bault and Génault 1999). This biogeographic connection likely began in the upper Paleocene (accounted by the records described by Rodríguez et al. 2023) and reached at least the middle Eocene or even the lower Oligocene.
This faunal distribution hypothesis is consistent with the proposed deflection of the circum-equatorial ocean circulation from the Tethys through the Pacific, as proposed by Lawver and Gahagan (2003: fig. 2). In this sense, the Paleogene of central Chile shows a display of taxa with widespread representation in the WBP, together with taxa previously considered as typical of the northern Atlantic.
Algarrobo can be considered as the northernmost available Late Cretaceous locality with faunal assemblages typical of the WBP. Its paleolatitude has remained stable during the Late Cretaceous and onwards, being mostly affected by W-E subduction that caused variations in the basin shape and a successive uplift trend (Gianni et al. 2018). Considering this, the marine vertebrate faunal turnover here disclosed occurred in a paleolatitude of ca. 33°S (almost the same as today). This implies that, at least during the late Paleocene, typical Tethyan chondrichthyan fauna reached mid-latitudes of the southeastern Pacific, a condition that remained at least until the middle–late Eocene. On the contrary, the chondrichthyan diversity known in austral high latitudes, show a clear different diversity with strong affinities between southernmost South America (Otero et al. 2012, 2013; Otero and Soto-Acuña 2015; Agnolín et al. 2021), Antarctica (Welton and Zinsmeister 1980; Long 1992; Kriwet 2005; Reguero et al. 2012; Otero et al. 2014; Engelbrecht et al. 2017; Charnelli et al. 2024) and even New Zealand (Mannering and Hiller 2008). These provincial-level affinities suggest that the Atlantic chondrichthyan diversity had low interchange through austral seaways. In this sense, a more plausible explanation for low-latitude Pacific localities such Algarrobo relies on a seaway through the northern part of South America, where the model of the Tethys Circumglobal Current (Bush 1997; Cousin-Rittemard et al. 2002) is consistent. The Tethyan Circumpolar Current was intermittent since the Jurassic, being functional during the Late Cretaceous and eventually reaching the late Paleogene (Bush 1997). Interestingly, during the Late Cretaceous, the functional Tethyan Circumpolar Current had no major influence over the Weddellian nature of the Algarrobo vertebrate assemblage. Thus, the drivers of the Paleogene turnover hypothesis along central Chile could be linked to other relevant austral paleogeographic changes occurred during that time span. Among these, a plausible scenario is the gradual appearance of the Antarctic Circumpolar Current (Lawver and Gahagan 2003) beginning during the early Eocene and initially linked to tectonics changes between Antarctica and Australia (in detriment of the former strong current from Australia/Antarctica, previously transiting the southern margin of the South Pacific and possibly heading northward along the west coast of South America; Lawver and Gahagan 2003: fig. 3), until a fully establishment of the Antarctic Circumpolar Current during the Middle Miocene linked to the expansion of the Antarctic Ice Sheet (Evangelinos et al. 2023).
Conclusions
This contribution describes the first Paleogene chondrichthyan assemblage from Algarrobo, in central Chile. The basal conglomerate of the Algarrobo Beds includes a diverse assemblage comprising in situ teeth of Striatolamia macrota, Cretolamna sp., Abdounia beaugei aff. Carcharhinus sp., and Rhinoptera sp., plus transported and likely redeposited teeth referred to Carcharias sp., and aff. Xiphodolamia sp. Additional specimens referable to Rhinoptera sp. were recovered in overlying levels, together with the first austral record of the ray Sulcidens sulcidens. The occurrence of the latter species is remarkable, because this was previously constrained to the Selandian–lower Ypresian of north Africa. The new record represents its first occurrence in the Pacific. The combined biochron of the recovered taxa indicates that the lower part of the Algarrobo Beds was deposited during the Ypresian (lower Eocene).
Previous records of Palaeogaleus, Physogaleus, and Premontreia in the upper Paleocene of central Chile already shown that typical Atlantic taxa present in the Pacific during that time span. The new diversity from the Ypresian of Algarrobo adds to this emerging picture, with the presence of former typical north African taxa represented by Sulcidens sulcidens, Abdounia beaugei, aff. Carcharhinus sp., and likely, the north Atlantic Xiphodolamia. The Atlantic-Pacific faunal interchange pattern is also suggested to occur during the Eocene–lower Oligocene, based on the Atlantic-Pacific known distribution of blochiid bilfishes.
The chondrichthyan assemblage accumulated to date in the Paleogene of central Chile differs from the endemic records that characterized the Weddellian Biogeographic Province since the Late Cretaceous until the Paleogene. The fauna from Algarrobo complements this scenario by providing a first Ypresian assemblage with marked north Atlantic influence, suggesting a temporal and geographical declination of the Weddellian Biogeographic Province began between the 33–37°S, at least from the late Paleocene until the early Oligocene, prior to the establishment of the paleogeographic and environmental conditions that molded the southeastern Pacific marine vertebrate fauna since the Neogene and onwards.
Acknowledgements
Israel Meyer-Navia and Juan Carlos Escalante (both MUSA) are thanked for managing the incorporation of the material studied to the paleontology collection of MUSA. The authors thank the Algarrobo inhabitants for their permanent concern on this remarkable fossil site. Two anonymous reviewers are thanked for all their comments and suggestions that improved the original draft. This research was supported by the Núcleo Milenio EVOTEM (ANID-MILENIO-NCN2023-025).
Editor: Camila Cupello
References
Adnet, S., Hosseinzadeh, R., Antunes, M.T., Balbino, A.C., Kozlov, V.A., and Cappetta, H. 2009. Eocene shark genus Xiphodolamia (Chondrichthyes, Lamniformes) and description of a new species recovered from Angola, Iran and Jordan. Journal of African Earth Sciences 55: 197–204. Crossref
Agassiz, L. 1843. (1833–44). Recherches sur les poissons fossiles. Vol. 3. 390+332 pp. Neuchâtel. Crossref
Agnolín, F., Bogan, S., and Casadío, S. 2021. First marine ichthyofauna from the Late Eocene of Santa Cruz province, Patagonia, Argentina. Journal of South American Earth Sciences 107: 103064. Crossref
Arambourg, C. 1935. Note préliminaire sur les vertébrés fossiles des phosphates du Maroc. Bulletin de la Société géologique de France 5: 413–439.
Arambourg, C. 1952. Les vertébrés fossiles des gisements de phosphates (Maroc-Algérie-Tunisie). Notes et Mémoires du Service Géologique du Maroc 92: 1–372.
Baut, J.P. and Génault, B. 1999. Les élasmobranches des Sables de Kerniel (Rupélien), à Gellik, Nord Est de la Belgique. Memoirs of the Geological Survey of Belgium 45: 1–61.
Berg, L.S. 1958. System der rezenten und fossilen Fischartigen und Fische. 310 pp. VEB Verlag der Wissenschaften, Berlin.
Blainville, de, H.M.D. 1816. Prodrome d’une nouvelle distribution systematique du regne animal. Bulletin de la Société Philomatique de Paris 8:105–124.
Bogan, S. and Agnolín, F.L. 2010. Primera ictiofauna marina del Cretácico Superior (Formación Jagüel, Maastrichtiano) de la Provincia de Río Negro, Argentina. Papéis Avulsos de Zoología 50: 175–188. Crossref
Bogan, S., Agnolín, F.L., and Novas, F.E. 2016. New selachian records from the Upper Cretaceous of southern Patagonia: paleobiogeographical implications and the description of a new taxon. Journal of Vertebrate Paleontology 36: e1105235.. Crossref
Bonaparte, C.L. 1838. Selachorum tabula analytica, Systema Ichthyologicum. Memoires de la Societe Neuchateloise des Sciences Naturelles 2: 1–16.
Boulemia, S. and Adnet, S. 2013. A new Palaeogene elasmobranch fauna (Tebessa region, eastern Algeria) and the importance of Algerian-Tunisian phosphates for the North African fossil record. Annales de Paléontologie 109: 102632. Crossref
Brüggen, J. 1915. El Cretáceo de Algarrobo. 15 pp. Sociedad Imprenta Litográfica Barcelona, Santiago-Valparaíso.
Bush, A.E.G. 1997. Numerical Simulation of the Cretaceous Tethys Circumglobal Current. Science 275: 807–810. Crossref
Cañete, N.G., Villafaña, J.A., Farias, L., Gayo, E.M., and Rivadeneira, M.M. 2025. The fossil record of chondrichthyans (Elasmobranchii, Holocephali) from the Cretaceous–Eocene in Chile: diversity and palaeobiogeographic implications. Historical Biology 38: 35–53. Crossref
Cappetta, H. 1980. Modification du statut générique de quelques espèces de sélaciens crétacés et tertiaires. Palaeovertebrata 10: 29–42. Crossref
Cappetta, H. 1987. Chondrichthyes II: Mesozoic and Cenozoic Elasmobranchii. In: H.P. Schultze (ed.), Handbook of Paleoichthyology, Vol. 3B, 1–193. Gustav Fischer Verlag, Stuttgart.
Cappetta, H. 2012. Handbook of Paleoichthyology, Vol. 3E: Chondrichthyes, Mesozoic and Cenozoic Elasmobranchii: Teeth, 512 pp. Verlag Dr. Friedrich Pfeil, München.
Carlsen, A.W. and Cuny, G. 2014. A study of the sharks and rays from the Lillebælt Clay (Early–Middle Eocene) of Denmark, and their palaeoecology. Bulletin of the Geological Society of Denmark 62: 39–88. Crossref
Charnelli, M., Gouiric-Cavalli, S., Reguero M.A., and Cione A.L. 2024. Middle Eocene chondrichthyan fauna from Antarctic Peninsula housed in the Museo de La Plata, Argentina. Advances in Polar Science 35: 14–47.
Compagno, L.J.V. 1973. Interrelationships of living elasmobranchs. In: P.H. Greenwood, R.S. Miles, and C. Patterson (eds.), Interrelationships of Fishes. Zoological Journal of the Linnean Society 53 (Supplement 1): 15–61.
Compagno, L.J.V. 1977. Phyletic relationships of living sharks and rays. American Zoologist 17: 303–322. Crossref
Compagno, L.J.V. 1984. Sharks of the world. An annotated and illustrated catalogue of shark species known to date. Part 1. Hexanchiformes to Lamniformes. FAO Species Catalogue Vol. 4, 125–249. Rome.
Cousin-Rittemard, N.M.M., Dijkstra, H.A., and Zwagers, T. 2002. Was there a wind-driven Tethys circumglobal current in the Late Cretaceous? Earth and Planetary Science Letters 203: 741–753. Crossref
Cunningham, S.B. 2000. A comparison of isolated teeth of early Eocene Striatolamia macrota (Chondrichthyes, Lamniformes), with those of a Recent sand shark, Carcharias taurus. Tertiary Research 20: 17–31.
Cuvier, G.L.C.F.D. 1829. Le règne animal, distribué d’après son organisation, pour servir de base à l’histoire naturelle des animaux et d’introduction à l’anatomie comparée. Edition 2. 566 pp. Chez Déterville, Libraire, Paris. Crossref
Dartevelle, E. and Casier, E. 1943. Les poissons fossiles du Bas-Congo et des régions voisines. Annales du Musée du Congo Belge, Sér. A (Minéralogie Géologie, Paléontologie) 3 2: 1–200.
Ebert, D.A. and Stehmann, M. 2013. Sharks, batoids, and chimaeras of the North Atlantic. FAO Species Catalogue for Fishery Purposes 7: 1–523.
Engelbrecht, A., Mörs, T., Reguero, M.A., and Kriwet, J. 2017. Revision of Eocene Antarctic carpet sharks (Elasmobranchii, Orectolobiformes) from Seymour Island, Antarctic Peninsula. Journal of Systematic Palaeontology 15: 969–990. Crossref
Evangelinos, D.,Etourneau, J., van de Flierdt, T., Crosta, X., Jeandel, C., Flores, J.-A., Harwood, D.M., Valero, L., Ducassou, E., Sauermilch, I., Klocker, A., Cacho, I., Pena, L. D., Kreissig, K., Benoit1, M., Belhadj, M., Paredes, E., Garcia-Solsona, E., López-Quirós, A., Salabarnada, A., and Escutia, C. 2023. Late Miocene onset of the modern Antarctic Circumpolar Current. Nature Geoscience 17: 165–270. Crossref
Fernández-Jiménez, R., Otero, R.A., Soto-Acuña, S., Flores, G., Bonilla, R., González, A., and Quinzio, L.A. 2016. New records of Paleogene Elasmobranchii from the coast of Coliumo, Biobío Province, Chile. In: C. Salazar, P. Zambrano, K. Cisterna, E. Bostelmann, F. Mourgues, A. Abarzúa, and T. Torres (eds.), V Simposio de Paleontología de Chile, Libro de Resúmenes, 85. Universidad Andrés Bello, Concepción.
Fernández-Jiménez, R., Groz, C., Otero, R.A., Manríquez, L., and Soto-Acuña, S. 2018. Nuevos Hallazgos de elasmobranquios de la Formación Quiriquina, Región del Biobío. In: M. Leppe, A. Abarzúa, and R. Villa (eds.), Primer Congreso de Paleontología de Chile, Abstracts, 340. Instituto Antártico Chileno, Punta Arenas.
Fierstine, H. and Monsch, K. 2002. Redescription and phylogenetic relationships of the family Blochiidae (Perciformes: Scombroidei), middle Eocene, Monte Bolca, Italy. Miscellanea Paleontologia, Studi e Richerche sui Giacimenti Terziari de Bolca, Museo Civico di Storia Naturale di Verona 9: 121–163.
Gana, P., Wall, R., and Gutiérrez, A. 1996. Mapa Geológico del área de Valparaíso-Curacaví, Region de Valparaíso y Región Metropolitana. Escala 1:100.000. Servicio Nacional de Geología y Minería (Chile), Mapas Geologicos N °1, Santiago.
Gasparini, Z. 1979. Comentarios críticos sobre los vertebrados mesozoicos de Chile. Actas del II Congreso Geológico Chileno, Arica 3: H15–H32.
Geoffroy Saint-Hilaire, E. 1817. Poissons du Nil, de la Mer Rouge et de la Méditerranée. In: E.F Jomard (ed.), Description de l’Egypte ou recueil des observations et des recherches qui ont été faites en Égypte pendant l’expedition de l’Armée française, publié par les ordres de sa Majesté-eL’Empereur Napoléon le Grand, 311–340. Imprimerie Impériale, Paris.
Gianni, G., Echaurren, A., Fennell, L., Navarrete, C., Quezada, P., Tobal, J., Jiménez, M., Dávila, F., and Folguera, A. 2018. Cretaceous orogeny and marine transgression in the Southern Central and Northern Patagonian Andes: Aftermath of a large-scale flat-subduction event? In: A. Folguera, E. Contreras-Reyes, N. Heredia, A. Encinas, S. Iannelli, V. Oliveros, F. Dávila, G. Collo, L. Giambiagi, A. Maksymowicz, M. Iglesia Llanos, M. Turienzo, M. Naipauer, D. Orts, V. Litvak, O. Álvarez, and C. Arriagada (eds.), The Evolution of the Chilean-Argentinean Andes, 291–328. Springer Earth System Sciences, Cham. Crossref
Glikman, L.S. 1958. Rates of evolution in lamnoid sharks [in Russian]. Doklady Akademii Nauk SSSR 123: 568–571.
Glikman, L. 1964. Akuly paleogena i ih stratigrafičeskoe značenie. 228 pp. Akademiâ Nauk Soûza Sovetskih Sotsialističeskih Respublik, Moskva.
Glikman, L. 1980. Evolûtsiâ melovyh i kainozoiskih lamnoidnyh aku. 247 pp. Akademiâ Nauka SSSR, Moskva.
Groz, C. and Palma-Heldt, S. 2013. Contribution of fossil record of elasmobranchs to the knowledge of the K/P limit in the Biobio Region, Chile. Geosur 2013, Abstracts, 234. Universidad Andrés Bello, Viña del Mar.
Huxley, T.H. 1880. On the application of the laws of evolution to the arrangement of the Vertebrata, and more particularly of the Mammalia. Proceedings of the Zoological Society of London 43: 649–661.
Jordan, D.S. and Evermann, B.W. 1896. The fishes of North and Middle America. Bulletin of the United States National Museum 47: 1–1240.
Jordan, D.S. 1898. Description of a species of fish (Mitsukurina owstoni) from Japan, the type of a distinct family of lamnoid sharks. Proceedings of the California Academy of Sciences 3: 199–204.
Kriwet, J. 2005. Additions to the Eocene Selachian fauna of Antarctica with comments on Antarctic Selachian diversity. Journal of Vertebrate Paleontology 25: 1–7. Crossref
Lawver, L.A. and Gahagan, L.M. 2003. Evolution of Cenozoic seaways in the circum-Antarctic region. Palaeogeography, Palaeoclimatology, Palaeoecology 198: 11–37. Crossref
Leidy, J. 1877. Description of vertebrate remains, chiefly from the phosphate beds of South Carolina. Journal of the Academy of Natural Sciences of Philadelphia 8: 209–261.
Leriche, M. 1905. Les poissons éocènes de la Belgique. Mémoires du Musée Royal d’Histoire Naturelle de Belgique 3: 49–228.
Long, D.J. 1992. Sharks from the La Meseta Formation (Eocene), Seymour Island, Antarctic Peninsula. Journal of Vertebrate Paleontology 12: 11–32. Crossref
Mannering, A.A. and Hiller, N. 2008. An Early Cenozoic neoselachian shark fauna from the Southwest Pacific. Palaeontology 51: 1341–1365. Crossref
Müller, J. and Henle, F. 1839. On the generic characters of cartilaginous fishes, with descriptions of new genera. Magazine of Natural History 2: 33–37.
Muñoz-Ramírez, C.P., Zambrano, P., Montoya, G., and Moyano, H. 2007. Dientes de tiburones y rayas (Chondrichthyes, Elasmobranchii) de la Formación Quiriquina aflorante en Talcahuano, Chile Central. Boletín de la Sociedad de Biología de Concepción 78: 7–22.
Oliver-Schneider, C. 1936. El Odontaspis elegans (Agassiz) en el Terciario Eoceno de Chile. Comunicaciones del Museo de Concepción 4: 77–78.
Otero, R.A. 2019a. Loancorhynchus catrillancai gen. et sp. nov., a new swordfish (Xiphioidei, Blochiidae) from the Middle Eocene of central Chile. PeerJ 7: e6671. Crossref
Otero, R.A. 2019b. Myledaphus araucanus sp. nov. (Batomorphi, Rajiformes incertae sedis), a new Late Cretaceous ray from the austral Pacific, and first occurrence of the genus in the Southern Hemisphere. Cretaceous Research 100: 82–90. Crossref
Otero, R.A. 2024. Review of two marine vertebrate assemblages from the Arauco Basin (central Chile) reveals diversity changes throughout the Maastrichtian. Cretaceous Research 166: 105996. Crossref
Otero, R.A. 2025. New Paleogene records of cartilaginous fishes (Chondrichthyes) from central Chile, including the oldest lamnid diversity from the southeastern Pacific. PeerJ 13: e19996. Crossref
Otero, R.A. and Soto-Acuña, S., 2015. New chondrichthyans from Bartonian–Priabonian levels of Río de Las Minas and Sierra Dorotea, Magallanes Basin, Chilean Patagonia. Andean Geology 42: 268–283. Crossref
Otero, R.A., Gutstein, C.S., Vargas, A.O., Rubilar-Rogers, D., Yury-Yáñez, R., Bastías, J., and Ramírez, C. 2014. New chondrichthyans from the Late Cretaceous (Campanian–Maastrichtian) of Seymour and James Ross islands, Antarctica. Journal of Paleontology 88: 411–420. Crossref
Otero, R.A., Oyarzún, J.L., Soto-Acuña, S., Yury-Yáñez, R., Gutiérrez, N., Le Roux, J., Torres, T., and Hervé, F. 2013. Neoselachians and Chimaeriformes (Chondrichthyes) from the Upper Cretaceous–Paleogene of Sierra Baguales, southernmost Chile. Chronostratigraphic, paleobiogeographic and paleoenvironmental implications. Journal of South American Earth Sciences 48: 13–30. Crossref
Otero, R.A., Torres, T., Le Roux, J.P., Hervé, F., Fanning, C.M., Yury-Yañez, R., and Rubilar-Rogers, D. 2012. A late Eocene age proposal for the Loreto Formation, (Brunswick Peninsula, southernmost Chile), based on fossil cartilaginous fishes, paleobotany and radiometric evidence. Paleontological Note. Andean Geology 39: 180–200. Crossref
Philippi, R.A. 1887. Los Fosiles Terciarios i Cuartarios de Chile. 236 pp. F.A. Brockhaus printing, Leipzig.
Quoy, J.R.C. and Gaimard, J.P. 1824. Description des Poissons. Chapître IX. In: L. de Freycinet (ed.), Voyage autour du monde, entrepris par ordre du roi. Exécuté sur les corvettes de S.M. l’Uranie et la Physicienne, pendant les années 1817, 1818, 1819 et 1820, 192–401 [1–328 in 1824; 329–616 in 1825], pls. 43–65. Chez Pillet aîné, Paris.
Rafinesque, C.S. 1810.
Caratteri di alcuni
nuovi generi e nuove specie di animali e piante della Sicilia, con
varie osservazioni sopra i medisimi.
[i–iv] 3–69. Sanfilippo, Palermo. Crossref
Reguero, M.A., Marenssi, S.A., and Santillana, S.N. 2012. Weddellian marine/coastal vertebrates diversity from a basal horizon (Ypresian, Eocene) of the Cucullaea I Allomember, La Meseta Formation, Seymour (Marambio) Island, Antarctica. Revista Peruana de Biología 19: 275–284. Crossref
Rodríguez, D., Ward, D.J., and Quezada, J.A. 2023. Paleontology and stratigraphic implications of a late Paleocene elasmobranch assemblage in Talcahuano, southcentral Chile. Andean Geology 50: 217–247. Crossref
Schweitzer, C.E., Feldmann, R.M., Encinas, A., and Suárez, M. 2006. New Cretaceous and Eocene Callianassoidea (Thalassinidea, Decapoda) from Algarrobo, Chile. Journal of Crustacean Biology 26: 73–81. Crossref
Solé, F., Noiret, C., Desmares, D., Adnet, S., Taverne, L., de Putter, T., Mees, F., Yans, J., Steeman, T., Louwye, S., Folie, A., Stevens, N.J., Gunnell, G.F., Baudet, D., Kitambal, N., Yaya, N.K., and Smith, T. 2019. Reassessment of historical sections from the Paleogene marine margin of the Congo Basin reveals an almost complete absence of Danian deposits. Geoscience Frontiers 10: 1039–1063. Crossref
Suárez, M.E. 2015. Tiburones, rayas y quimeras (Chondrichthyes) fósiles de Chile. Publicación Ocasional del Museo Nacional de Historia Natural (Chile) 63: 17–33. Crossref
Suárez, M.E. and Marquardt, C. 2003. Revisión preliminar de las faunas de peces elasmobranquios del Mesozoico y Cenozoico de Chile. Congreso Geológico Chileno n° 10, Sesion temática 3, Paleontología. Actas, CD-ROM, 9 pp. Universidad de Concepción, Concepción.
Suárez, M.E., Quinzio, L.A., Fritis, O., and Bonilla, R. 2003. Aportes al conocimiento de los vertebrados marinos de la Formación Quiriquina. Congreso Geológico Chileno n° 10, Sesión temática 3, Paleontología. Actas, 7 pp. Universidad de Concepción, Concepción.
Tavera, J. 1942. Contribución al estudio de la estratigrafía y paleontología del Terciario de Arauco. Congreso Panamericano de Ingeniería de Minas y Geología 1, Anales Parte 1, 2: 580–632. Universidad de Chile, Santiago.
Tavera, J. 1980. Cretáceo y Terciario de la localidad de Algarrobo. 45 pp. Imprentas Gráficas Conchalí, Santiago.
Underwood, C.J., Kolmann, M., and Ward, D. J. 2017. Paleogene origin of planktivory in the Batoidea. Journal of Vertebrate Paleontology 37 (1): e1293068. Crossref
Welton, B., and Zinsmeister, W. 1980. Eocene Neoselachians from the La Meseta Formation, Seymour Island, Antarctic Peninsula. Natural History Museum of Los Angeles County, Contributions in Science 329: 1–10. Crossref
Wetzel, W. 1930. Die Quiriquina-Schichten als sediment und paläontologisches archiv. Palaeontographica 73: 49‒106.
Wilckens, O. 1904. Revision der Fauna der Quiriquina Schichten. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie Beilage-Band 18: 181–284. Crossref
Zinsmeister, W.J. 1979. Biogeographic significance of the Late Mesozoic and early Tertiary molluscan faunas of Seymour Island (Antarctic Peninsula) to the final break-up of Gondwanaland. In: J. Gray and A.J. Boucot (eds.), Historical Biogeography, Plate Tectonics and the Changing Environment, 349–355. Oregon State University Press, Corvallis, OR.
Acta Palaeontol. Pol. 71 (2): 255–266, 2026
https://doi.org/10.4202/app.01299.2025