
Early Paleocene siluroid catfish from Bolivia and its evolutionary and paleogeographic importance
EVA CARON, DIOGO MAYRINCK, RICARDO CESPEDES, and OLGA OTERO
Caron, E., Mayrinck, D., Cespedes, R., and Otero, O. 2026. Early Paleocene siluroid catfish from Bolivia and its evolutionary and paleogeographic importance. Acta Palaeontologica Polonica 71 (1): 109–118.
The fossil genus Yuskaichthys, previously known only from Eocene deposits of the Maíz Gordo site in Argentina, is here identified based on three isolated parieto-supraoccipital bones collected in the Tiupampa locality, Cochabamba-Potosí Basin, Bolivia. This discovery notably extends both the paleogeographic and temporal distribution of the genus into the early Paleocene. Morphological traits including the bifid posterior margin of the supraoccipital process and the transverse pit lines closely resemble those described for Yuskaichthys. Futhermore, these fish share a longitudinal groove along the entire supraoccpital process, supporting a provisional assignment to Yuskaichthys. The occurrence of Yuskaichthys in both Tiupampa and Maíz Gordo is consistent with regional fluvial systems associated with the Andean foreland basin evolution. This finding supports an early diversification of modern catfish lineages, including extinct genera displaying long time range.
Key words: Teleostei, Siluriformes, Yuskaichthys, catfish, Paleocene, Bolivia.
Eva Caron [eva.caron44@gmail.com, ORCID: https://orcid.org/0009-0009-7551-9679], Programa de Pós-Graduação em Biociências, Instituto de Biologia Roberto Alcântara Gomes, Universidade do Estado do Rio de Janeiro.Boulevard 28 de setembro 87, PAPC, Maracanã, 20551-030, Rio de Janeiro, Brazil. LICTAE (Laboratório de Ictiologia Aplicada: anatomia e evolução), Departamento de Ensino de Ciências e Biologia, Instituto de Biologia Roberto Alcântara Gomes, Universidade do Estado do Rio de Janeiro. Rua São Francisco Xavier 524, PHLC sala 504, Maracanã, 20559-900, Rio de Janeiro, Brazil.
Diogo Mayrinck [mdiogobio@yahoo.com.br, ORCID: https://orcid.org/0000-0002-6899-1432]. LICTAE (Laboratório de Ictiologia Aplicada: anatomia e evolução), Departamento de Ensino de Ciências e Biologia, Instituto de Biologia Roberto Alcântara Gomes, Universidade do Estado do Rio de Janeiro. Rua São Francisco Xavier 524, PHLC sala 504, Maracanã, 20559-900, Rio de Janeiro, Brazil.
Ricardo Cespedes [ricardojcespedes@gmail.com, ORCID: https://orcid.org/ 0000-0003-1004-8246], Museo de Historia Natural Alcide d’Orbigny, avenida Potosí n°1458, Cochabamba, Bolivia.
Olga Otero [olga.otero@univ-poitiers.fr, ORCID: https://orcid.org/0000-0002-8129-2054], Université de Poitiers, laboratoire PALEVOPRIM (UMR 7262 CNRS-UP), 6 rue Michel Brunet, 86000 Poitiers, France.
Received 13 September 2025, accepted 29 December 2025, published online 26 March 2026.
Copyright © 2026 E. Caron et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License (for details please see http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Introduction
With over 4000 extant species assigned to 39 families, Siluriformes is a highly diverse order of teleosts found predominantly in freshwater environments worldwide, although only 2 families occur in marine waters (Fricke et al. 2021). The Paleocene represents a critical interval for ray-finned fish evolution, as it follows the K/Pg mass extinction and likely witnessed the initial diversification of many acanthomorph and ostariophysan lineages, including catfishes, yet the fossil record for this interval remains sparse. Fossils from the Paleocene therefore provide essential insights into early evolutionary radiations and patterns of biogeography in these groups. The catfish fossil record begins in the Upper Cretaceous and Paleocene in South America and continues during the Cenozoic across all continents (Arratia 2025; Otero et al. 2025). Although fragmentary, the Cretaceous and Paleocene South American catfish fossils belong to extinct taxa and provide key insights into the early diversification of catfishes (Arratia 2025: fig. 1; Otero et al. 2025; Pinna et al. 2025). To date, Cretaceous and Paleocene genera and species are known only from their type locality or the corresponding paleo-basin and they exhibit a relatively short temporal span (Fig. 1). This contrasts sharply with extant taxa, some of which exhibit subcontinental distribution and/or long-time span. For instance, in South America, the extant genera Brachyplatystoma and Phractocephalus display a fossil record since at least 9 Ma (Ferraris 2007) and some phylogenetic studies including several species of the same genus show that many extant genera exist for 10 Ma or over, as in the case of Pimelodus Lacépède, 1803 (Sullivan et al. 2013). This may be due to high endemicity in South America during the Late Cretaceous and Paleocene periods, a highly lacunose fossil record, or a combination of both.
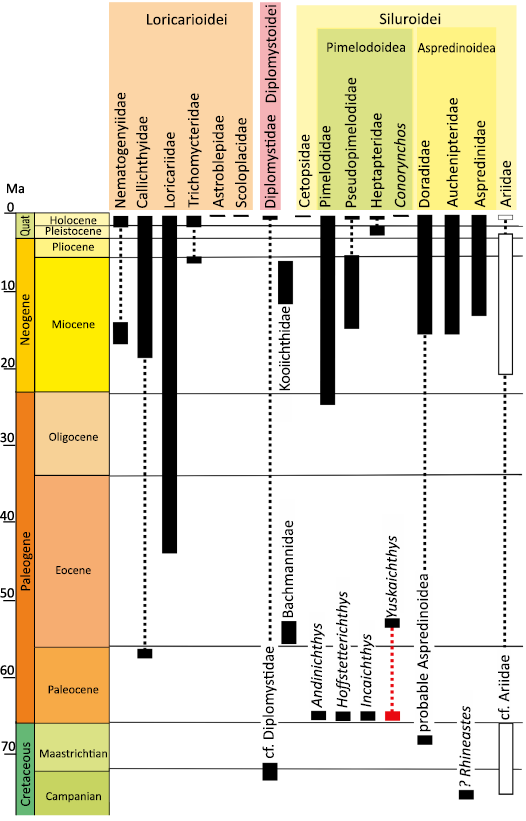
Fig. 1. Phylogeny of the Siluriformes and the fossil record of South American families, with details on extinct genera. Data from Arratia (2025), Otero et al. (2025), and Pinna et al. (2025). Rectangles, occurrences of taxa; red rectangles, studied taxon; white rectangles, marine taxa; dashed lines, absence of occurrences.
In the context of a review of paleontological material collected in Bolivia in the 1980s and 1990s from the Upper Cretaceous and Paleocene series of the Cochabamba-Potosí Basin (e.g., Otero et al. 2025), we rediscovered a parieto-supraoccipital bones collected in Tiupampa (Santa Lucía Formation) that had been mistakenly attributed to the catfish Incaichthys suarezi (Gayet 1991). Indeed, it does not match with the initial description and illustration of Incaichthys suarezi Gayet, 1990, but it does show a close resemblance to the taxon Yuskaichthys eocenicus (Bogan et al., 2018) from Argentina, a fossil species that the authors have proposed to link at the family level to an extinct genus from the lower Paleocene of Bolivia: Andinichthys sp. (Bogan et al. 2018; Otero et al. 2025). We also recovered two other parieto-supraoccipital bones that display the same morphology. Undoubtedly, these three bones resemble Yuskaichthys more than any other catfish, including those found in Bolivia. This paper aims to redescribe these fossils and discuss their attribution to the extinct genus Yuskaichthys, significantly extending its spatial and temporal range.
Institutional abbreviations.—MHNC, Museo de Historia Natural Alcide d’Orbigny de Cochabamba, Cochabamba, Bolivia; MNHN, Muséum national d’Histoire naturelle, Paris, France; MNRJ, Museu Nacional Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil.
Geological setting
The fossiliferous sites Estancia Blanco Rancho, Pajcha Pata, and Tiupampa are located in the Cochabamba-Potosí Basin. Department of Cochabamba, Bolivia. The mammalian levels in Tiupampa are dated as Danian (64–63 Ma) (Fig. 2; Muizon and Ladevèze 2020). Depending on the stratigraphic level studied, the Santa Lucia Formation’s aquatic paleoenvironments correspond to brackish or freshwater (Gayet et al. 1991). However, the Tiupampan mammalian levels are interpreted as lacustrine to meandering fluvial environments (Muizon et al. 1983; Gayet et al. 1991; Marshall et al. 1995).
The three parieto-supraoccipital fossils studied here were likely collected from the lower Paleocene mammalian levels at Tiupampa (Santa Lucía Formation), although slightly older levels at Estancia Blanco Rancho and Pajcha Pata cannot be excluded.
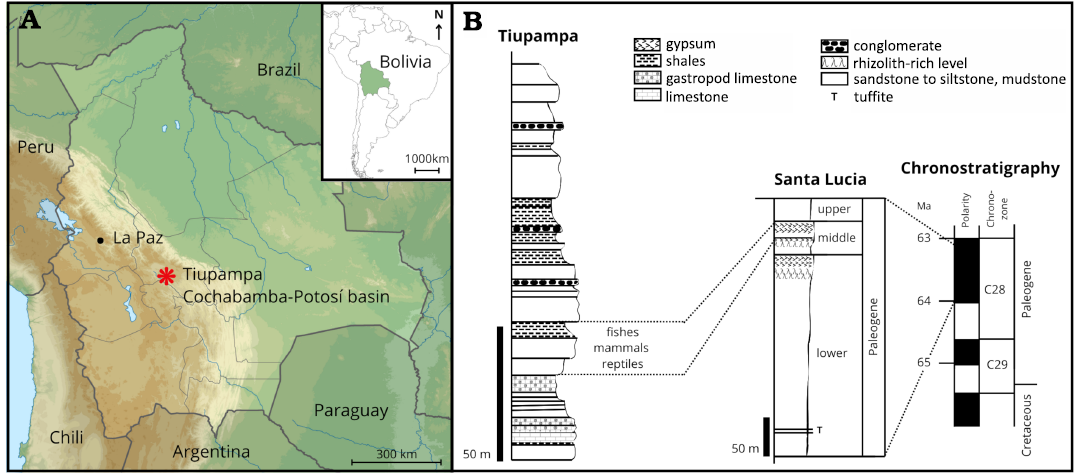
Fig. 2. Location (A), general stratigraphic column and age (B) of the fossiliferous levels at Tiupampa, Bolivia that probably yielded Yuskaichthys sp. fossils. Data from De Muizon et al. 1983; Gayet et al. 1991; Sempéré et al. 1997; Gelfo et al. 2009; de Muizon and Ladevèze 2020.
Material and methods
Study material.—The three fossils studied here belong to the paleontological collection of the MHNC (labelled by the Academia Nacional de Ciencias de Bolivia) and numbered MHNC 6291, 6294, 15350. Photographs and 3D data produced for this study are available from the authors upon request.
MHNC 6291 This specimen may have been treated with 5% acetic acid at the same time as certain other specimens from the formation, during the study of Arratia and Gayet (1995). If the specimen was indeed treated (although misidentification cannot be excluded; see below), its surface, internal structure, and density appear unaltered, based on observations from the CT scan reconstructions.
MHNC 6294 was figured and misnumbered as MHNC 6253 by Gayet (1991). Despite significant differences between the two specimens (Fig. 3), she presented the current MHNC 6294 as the holotype of Incaichthys suarezi, an extinct catfish species that had previously been described based on a different specimen (Gayet 1990). This misattribution likely resulted from changes in the fossil numbering. Specifically (Fig. 3), the holotype of Incaichthys suarezi is a fragment of the neurocranium, missing most of the anterior half and the posterior part of the parieto-supraoccipital bone, and was originally described under the number MHNC 6253 (Gayet 1990). In a subsequent publication, Gayet and Meunier (1998) provided an interpretative drawing of this holotype under the new number MHNC 8558, with the old number in brackets (Fig. 3B). The publication of MHNC 6294 with the erroneous identification (Gayet 1991) occurred between these two works, probably due to the renumbering of the Bolivian fossils, which caused a mix-up in the attribution of MHNC 6253. Later, the original numbers were restored: MHNC 8558 was reattributed to a Lepisosteidae vertebra (Gayet et al. 2001), the holotype of Incaichthys suarezi recovered its original number (MHNC 6253), and the parieto-supraoccipital misidentified by Gayet (1991) is now correctly known as MHNC 6294.
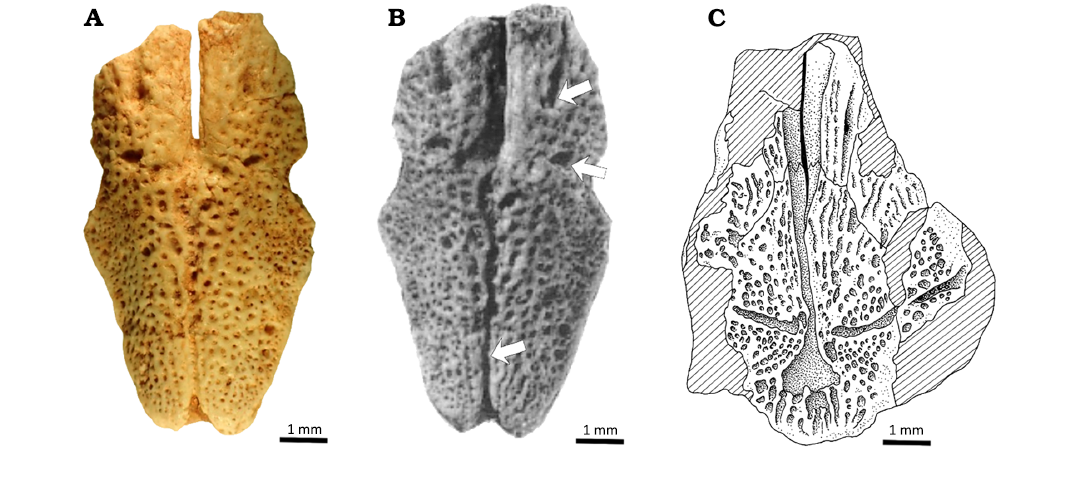
Fig. 3. Early Paleocene siluroid catfish Yuskaichthys sp. from Tiupampa, Bolivia. Parieto-supraoccipitals in dorsal view. A. MHNC 6294, former MHNC 6253 in Gayet (1991), B. MHNC 8558 (=6253), former holotype of Incaichthys suazeri, Gayet (1990) and re-numbered by Gayet and Meunier (1998). (modified from Gayet (1991), arrows indicate the differences from the valid Incaichthys suazeri; C. MHNC 8558 holotype modifed from Gayet and Meunier (1998).
In addition to these numbering issues, precise information on fossil sampling is scarce. Therefore, cross-checked published data and scattered notes must be used to reconstruct original collection information. All papers reporting on small catfish remains from Bolivia (variously classified as belonging to Andinichthyidae, Andinichthyoidea, or Andinichthyoidei) only report cranial remains in deposits of the Santa Lucia Formation in Tiupampa (see Otero et al. 2025, for details). Older levels (El Molino Formation) and other localities have only yielded fin spines, with the notable exceptions of Estancia Blanco Rancho (upper El Molino and Santa Lucía formations) and Pajcha Pata (upper El Molino and Santa Lucía), where neurocranial bones of Andinichthys sp. have been found (Gayet 1991; Otero et al. 2025). So, the exact collection levels of the three parieto-supraoccipital fossils studied here are unspecified, but they most probably correspond to the mammalian levels in Tiupampa (Muizon et al. 1983), or to slightly older deposits in Estancia Blanco Rancho, Pajcha Pata.
Comparative material.—Extant species: CT scan 3D rendering available from DigiMorph (2025): Conoryhnchos cornirostris (Cuvier, 1829), MNRJ23011; CT scan 3D renderings produced for this study: Batrochoglanis villosus (Eigenmann, 1912), MNHN-IC-2001-2054; Heptapterus sp., MNHN-IC-998-1782; Heptapterus tenuis (Mees, 1986), MNHN-IC-2001-1638; Megalonema platanum (Günther, 1880), MNHN-IC-1988-0834; Microglanis poecilus Eigenmann, 1912, MNHN-IC-2001-1425; Phractoce- phalus hemioliopterus Bloch & Schneider, 1801, MNHN-IC-A-1952; Pimelodella elongata (Günther, 1860), MNHN-IC-2004-3105; Propimelodus eigenmanni van der Stigchel, 1946, MNHN-IC-2002-0123; Pseudopimelodus bufonius Valenciennes, 1840, MNHN-IC-2000-5855; Pseudo-pimelodus nigricaudus Mees, 1974, MNHN-IC-2000-1533; Pseudopimelodus raninus (Valenciennes, 1840), MNHN- IC-2004-1301; Pseudopimelodus zungaro Bleeker, 1858, MNHN-IC-1981-0252; Sorubim lima (Bloch & Schneider, 1801), MNHN-IC-1988-0900; Rhamdia sp., MNHN-IC- 1988-0826.
Fossil species from Tiupampa, Santa Lucia Formation: Andinichthys boliviensis Gayet, 1988, MHNC6178 (holotype), MHNC6161; Hoffsteterichthys pucai Gayet, 1990, MHNC6252 (holotype), MHNC6181, MHNC6292; Incaichthys suarezi Gayet, 1990, MHNC6252 (holotype).
Imaging methods.—Microtomography of extant and fossil specimens was carried out at the Université de Poitiers (France) by Arnaud Mazurier (Université de Poitiers, IC2MP UMR 7285, Plateforme Caractérisation de Matériaux) on EasyTom XL Duo equipment developed by RX-solutions (Chavanod, France). Specifically, a Hamamatsu L10711 cooled X-ray source with nannofocus transmission was used with a LaB6 cathode coupled to a Varian PaxScan 2520DX detector (amorphous silicon flat panel and CsI conversion screen; 1920×1536 pixel matrix; 127 µm pixel pitch; 16-bit dynamic range). Segmentation of each specimen was carried out using Avizo version 8.1 software.
Systematic paleontology
Order Siluriformes Rafinesque, 1820
Suborder Siluroidei Frizzell & Dante, 1965
Family Andinichthyidae Gayet,1988
Genus Yuskaichthys Bogan et al., 2018
Type species: Yuskaichthys eocenicus Bogan et al., 2018, from Eocene of Argentina.
Yuskaichthys sp.
Fig. 4.
Material.—MHNC 6291, 6294, 15350, partial and complete isolated parieto-supraoccipitals. All from Tiupampa, Santa Lucía Formation (64–63 Ma). 3D data and photographs are available from the authors upon request.
Description.—The parieto-supraoccipitals display reticulated ornamentation that forms an anastomosing pattern on the outer face (Fig. 4). A transverse middle pitline groove is present on the anterior part of the bone and varies in shape among specimens. It appears as an irregular dotted line near the anterior constriction of the bone (Fig. 4A). Bones that would articulate with the parieto-supraoccipital on the skull roof and posterior wall of the braincase are not well preserved enough to ascertain their position. However, gently curved lateral notches at the base of the supraoccipital process may correspond to the articulation edge with an extrascapula (Fig. 4A). Moreover, the transverse pitline is usually placed at the pterotic level, which also supports this interpretation. The supraoccipital process spans the width of the bone and extends posteriorly, following an ovoid outline except for the deeply notched posterior tip, which ends in a bifid process. The anterior nuchal plate articulates with this notch, as evidenced by the fibrous aspect of the bone along the posterior margin of the process (Fig. 4A, B). A longitudinal groove runs along the parieto-supraoccipital bone, continuing along the entire length of the supraoccipital process (Fig. 4). Along the process, the groove widens slightly posteriorly and opens into the posterior notch (Fig. 4A, B). A shallow longitudinal lamina develops ventrally along the entire length of the supraoccipital process (Fig. 4).

Fig. 4. Early Paleocene siluroid catfish Yuskaichthys sp. from Tiupampa, Bolivia. A. MHNC 6294 partial parieto-supraoccipital in dorsal (A1, A3) and ventral (A2) views. B. MHNC 6291 posterior part of a partial parieto-supraoccipital in dorsal (B1, B2) and ventral (B3) views. C. MHNC 15350 fragment of a parieto-supraoccipital in dorsal (C1, C2) and ventral (C3) views. Photographs (A1, A3, B1, C1) and 3D virtual models (A2, B2, B3, C2, C3)
Discussion
The morphological variation observed in the parieto-supraoccipital among extant and fossil Siluriformes provides insights not only into taxonomic affinities but also into evolutionary trends in skull roof morphology within the group.
Common features of the
parieto-supraoccipital of Siluriformes.—The fusion
of the parietal with the supraoccipital is a synapomorphy
of Siluriformes (Arratia and Gayet 1995; Wiley
and Johnson 2010). Only a few exceptions have been
reported so far, typically concerning early developmental stages and
juveniles (Arratia and Gayet 1995) and most catfishes
do not retain a distinct parietal
ossification centre (Arratia 2003). Consequently, the
robust parieto-
supraoccipital bone largely covers the catfish braincase and the dorsal
portion of its posterior wall. The organisation of bones articulated
with the parieto-supraoccipital on the skull roof remains relatively
stable at the family level. In all catfishes, the parieto-supraoccipital
is bordered anteriorly by the frontal, laterally by
the sphenotic and pterotic, and depending on the family, also by
an extrascapula and/or
the epiotic (Fig. 5). The suture of the parieto-supraoccipital with the
sphenotic is a catfish apomorphy, and its absence in
certain Pseudopimelodidae represents a reversal (Arratia
and Gayet 1995). In Loricarioidei, the pterotic bone is compound
due to fusion with elements of the pectoral girdle (Fig.
5; Aquino and Schaefer 2002; Arratia
2003). These features confirm that the parieto-supraoccipital
morphology observed in the Tiupampa fossils is consistent with
Siluroidei, supporting their phylogenetic placement.
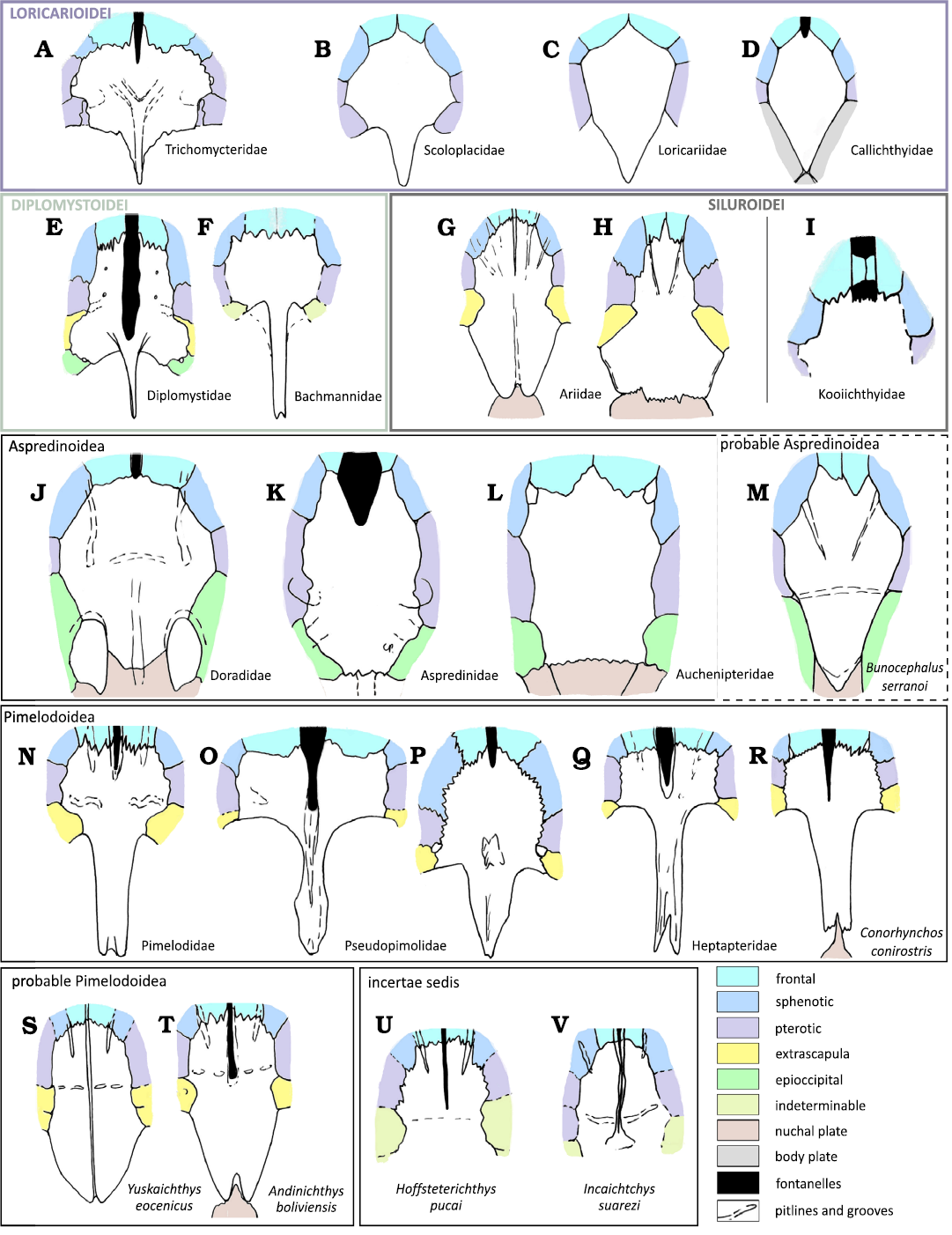
Fig. 5. Outline, bone connection, longitudinal
fontanelles and groove extent, anterior and transversal groove or
pit line position in the parieto-supraoccipital of members of extant
families and extinct genera from South America (when known), in
dorsal views. Loricarioidei: Nematogenys
inermis (A), Scoloplax dicra (B)
and Otocinclus vittatus (C), Corydoras
aeneus (D).
Diplomystoidei: Diplomystes camposensis
(E), Bachmannia
chubutensis (F).
Siluroidei: Ariidae: Sciades dowii (G), Ariopsis
felis (H),
Incertae sedis: Kooichthys jono
(I). Aspredinoidea: Tenellus ternetzi (J),
Bunocephalus hartti (K), Entomocorus
gameroi (L), Bunocephalus serranoi (M).
Pimelodoidea: Brachyplatystoma
vaillantii (N),
Microglanis poecilus
(MNHN-IC-2001-1425 (O), Pseudopimelodus raninus
(MNHN-IC-2004-1301) (P), Pimelodella elongata (MNHN-IC-2004-3105)
(Q), Conorhynchos
conirostris (MNRJ 23011) (R).
Paleocene fossils from Argentina and Bolivia: Yuskaichthys
eaocenicus (S),
Andinichthys bolivianenses (T), Hoffstetterichthys
pucai (U), Incaichthys suazeri (V).
Not to scale. Data modified from Arratia
2003 (A–C, E); Huysentruyt and Adriaens 2005
(D); Azpelicueta and Cione 2011 (F); Aguilera
and Marceniuk 2018 (G); Marceniuk et al. 2017
(H); de las Mercedes Azpelicueta et al. 2015 (I);
Birindelli 2014 (J); Carvalho et al.
2015 (K); Calegari et al. 2019 (L); Bogan
and Agnolin 2024 (M); Aguilera et
al. 2013 (N); DigiMorph (O–R); Bogan et al. 2018
(S); Otero et al. 2025 (T–V).
Features of the parieto-supraoccipital variable among Siluriformes.—The supraoccipital process shows considerable variation among Siluriformes. It may extend posteriorly in a dorsal direction and sometimes reach the nuchal plates. In some cases, the process is completely sutured to form a nuchal shield, as observed in Aspredinioidae and Loricariidae (Fig. 5; Birindelli 2014). In other South American families, such as Pimelodidae and Ariidae, only the posterior tip or border may suture with the anterior-most nuchal plate (e.g., Pimelodidae, Fig. 5; Marceniuk and Menezes 2007; Ariidae, Marceniuk et al. 2012), a condition also seen in the fossils described here (see Fig. 4). More specifically, in these fossils, the supraoccipital process displays a bifid tip with connecting bony tissue in a notch of the nuchal plate, a specific condition also seen in many Pimelodoidae and Ariidae. Within many families, and even genera, the supraoccipital process varies in width, shape, and length (see e.g., Pimelodidae in Fig. 5), while in some others it remains reduced, such as the Diplomystidae (Arratia 1987). When fully developed, a well-defined longitudinal ventral lamina is present, as observed in the fossils described here (Fig. 4).
The arrangement and ornamentation of grooves also vary across and within catfish families. Grooves or aligned pits often develop on the parietal-supraoccipital roof, forming anterior grooves (or pit lines) shared with the frontals and sometimes the sphenotic edge. These structures correspond to the parietal branch of the sensory canal and to a transverse groove or pit line. Both features are present in the fossils described here (Fig. 4). As noted by Arratia and Gayet (1995), these grooves and pit lines are often associated to neuromast lines, although bony relief is not always present. Anterior grooves are common in Siluroidei, whereas transverse pitline grooves are less common but present in certain Pimelodoidea. Occasionally, the posterior fontanel groove extends to the anterior edge of the parieto-supraoccipital bone and sometimes bisects it. In certain species, the fontanel, or at least the groove associated, extends to the back of the braincase (e.g., Diplomystidae and certain Pimelodidae, see Fig. 5). Finally, to our knowledge, a longitudinal groove continuing onto the supraoccipital process has been observed only in Yuskaichthys eocenicus (Bogan et al. 2018) and in the fossils described here (Fig. 4).
Ornamentation.—In the fossils described here, reticulated ornamentation is consistently observed on the outer face, both to the naked eye and in 3D reconstructions. In Yuskaichthys eocenicus (Bogan et al. 2018), ornamentation appears to range from granulated to reticulated, depending on the specimen (Bogan et al. 2018: fig. 2A, F). However, the resolution of the figures limits a detailed assessment of this variation. The perception of ornamentation may vary depending on the orientation of the light and may additionally be influenced by the state of preservation of the specimens. Ornamentation is also highly variable among catfish families and sometimes even within the same species (e.g., Yuskaichthys eocenicus, Bogan et al. 2018), as reported in comparative studies of cranial morphology (Murray and Holmes 2021). These observations highlight that ornamentation alone is an unreliable character for taxonomic attribution in this group. Therefore, while informative for morphological description, this feature is not considered a determining factor in assigning the three Bolivian parieto-supraoccipitals to a particular genus, underscoring the need to integrate multiple cranial features in systematic analysis.
Features of the fossil parieto-supraoccipitals and their distribution in extant and extinct South American catfishes.—We interpret the three fossil parieto-supraoccipitals bones described here (Fig. 4) as probable Yuskaichthys, since they share all the preserved features of the Argentinian specimen, including:
(i) a probable dorsal connection with four cranial bones, including an extrascapula (also present in some Pimeloidoidea, Ariidae and Bolivian fossils attributed to Andinichthyidae and Yuskaichthys among others);
(ii) a large, elongated supraoccipital process extending along the entire parieto-supraoccipital edge of the braincase, with a bifid end that testifies to its articulation with a pointed anterior nuchal plate (also present in certain Pimelodoidea and Ariidae, and in Bolivian fossils attributed to Andinichthyidae and Yuskaichthys among South American catfishes);
(iii) anterior grooves, frequent in South American catfishes (notably Pimelodoidea, Ariidae, Bolivian fossils attributed to Andinichthyidae and Yuskaichthys);
(iv) transverse pit lines with two pits on each side at the level of the pterotic-extrascapular suture (also observed in some Pimelodoidea, in Bolivian fossils attributed to Andinichthyidae and Yuskaichthys);
(v) a longitudinal fontanelle extending over half of the skull roof, with a surrounding groove reaching the posterior part of the braincase (also seen in Diplomystidae, some Pimelodidae, some Ariidae, and Yuskaichthys);
(vi) a narrow longitudinal groove continuing along the entire length of the supraoccipital process (a feature unique to Yuskaichthys).
The fossils described here show several affinities with both Pimelodoidea and Ariidae. However, they differ notably from Ariidae by exhibiting a posterior fontanel that extends onto the parieto-supraoccipital, combined with a broad and elongated supraoccipital that is posteriorly ended by a bifid process. As well as a markedly developed transverse pitline groove, the latter being a more consistent trait only known with some Pimelodoidea. Notwithstanding, the combination of features (i–v) on the three Tiupampan specimens is consistent with those seen in certain extant pimelodids, and also in the extinct genera Yuskaichthys and Andinichthys (Fig. 5). However, feature (vi), which is unique to Yuskaichthys, supports their probable assignment to this extinct genus. This is the most parsimonious hypothesis.
Parieto-supraoccipital preservation.—Most of the skull material recovered from Tiupampa and attributed to small catfishes consists of articulated neurocranial fragments (Otero et al. 2025), making the disarticulated parieto-supraoccipital bones described here particularly notable. This condition may reflect either weaker suturing of the neurocranial bones in this fossil species or the fact that these specimens represent juvenile individuals. Such observations are significant because they provide insights into the developmental biology of early catfishes and may influence the interpretation of morphological variation in both fossil and extant taxa.
Paleobiogeography and paleoenvironment.—The ancient hydrographical connections in Central South America likely reflect the evolving drainage systems and foreland basin dynamics of the region (Fig. 6; Horton and DeCelles 1997; Lundberg et al. 1998). During the late Maastrichtian–early Paleocene period, a major fluvial system developed along the underfilled axis of the Andean foreland basin, flowing northwards (Sempéré et al. 1997). This system was the precursor to the modern Amazon and Orinoco basins. Both localities were situated in the upper reaches of this system (Fig. 6). Although a marine transgression reached the region around 61–60 Ma, sedimentary evidence indicates a return to freshwater conditions by ~59 Ma (Sempéré et al. 1997; Lundberg et al. 1998). The Santa Lucía and Maíz Gordo formations were deposited in lacustrine to palustrine environments within the back-bulge of the foreland system (Gayet et al. 1991; Sempéré et al. 1997), which is possibly comparable to the modern Pantanal (Horton and DeCelles 1997). Continental continuity, combined with tectonic quiescence and sedimentary stability, may have facilitated the dispersal of freshwater taxa between basins. The shared ichthyofauna between the Santa Lucía and Maíz Gordo formations is therefore not surprising and reflects the geographic proximity and hydrographic coherence of the central Andean foreland during the Paleocene (Marshall et al. 1997; Lundberg et al. 1998).
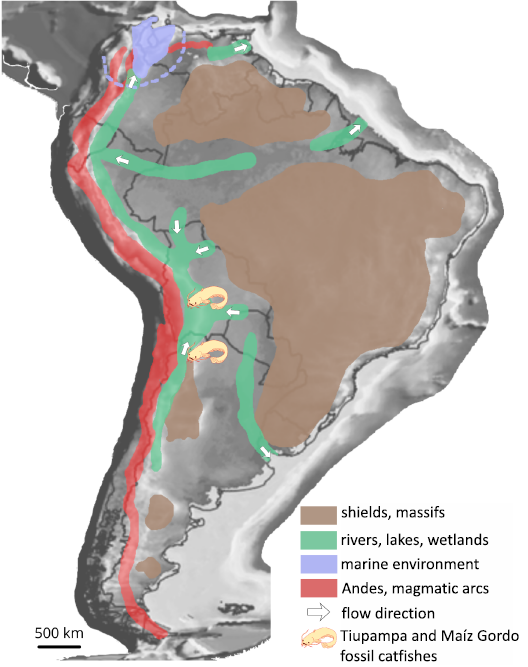
Fig. 6. Paleoenvironments in South America around
60–45 Ma (modified from Lundberg et al.
1998; Bogan et al. 2018) on a paleogeographical
map (modified from Scottese 2001), with location of
the Yuskaichthys fossil
localities.
Extending the temporal and geographical range of an extinct genus.—The geographical and hydrographical proximity of the Yuskaichthys from Tiupampa in Bolivia and Maiz Gordo in Argentina (around 550 km), as well as the long time elapsed between their respective occurrences (between 9 and 14 million years), do not demonstrate remarkable dispersal capacity (only a minimum of 55 km per million years, which corresponds to 5.5 cm per year). However, for the first time in South America, an extinct genus of catfish shows an extension of time comparable to that known in many extant genera, such as Corydoras since 58 Ma (Cockerell 1925), Nematogenys since ~13 Ma (Azpelicueta and Rubilar 1998) or Brachyplatystoma and Phractocephalus since ~9 Ma (Lundberg and Aguilera 2003; Lundberg 2005). In this case, despite the relatively important quantity and quality of the catfish fossils in the Campanian–Maastrichtian and Paleocene of Bolivia (Otero et al. 2025), only three isolated bones can be attributed to Yuskaichthys sp., which, in our opinion, suggests a very incomplete paleontological record of the early stages of catfish evolution. In any case, this demonstrates the importance of collecting efforts in these key levels to understand the evolution of catfishes, in Bolivia and elsewhere in South America.
Concluding remarks
The discovery of provisional Yuskaichthys fossils in Paleocene deposits from the Potosí-Cochabamba basin, Bolivia, dated 64–63 Ma, extends the temporal and geographical range of this genus of early catfish, which was previously known only from Eocene deposits in north-west Argentina, dated 54–50 Ma (Bogan et al. 2018). This discovery indicates ancient hydrographical connections between these regions and suggests a long temporal and a wide geographical span for a Paleogene South American catfish genus for the first time. Future work should focus on continued collection and study of Paleocene catfish material to further clarify the early diversity and evolutionary history of South American Siluriformes.
Authors’ contributions
OO designed the study; EC and OO prepared and analysed the data, designed the figures and wrote initial version of the paper; RC cures the material; EC, DM, RC and OO wrote the final version of the paper.
Acknowledgements
We thank Arnaud Mazurier (Université de Poitiers, France) for CT scanning and 3D volume reconstruction, and also the reviewers of the paper. We also thank Donald Davesne (Sorbonne Université, Paris, France) and the anonymous reviewer for their constructive comments, which helped improve this manuscript. We are grateful to the editor, Camila Cupello (Departamento de Arqueologia, Universidade do Estado do Rio de Janeiro) for her helpful comments and for handling the manuscript. This work was supported by PALEVOPRIM (CT scan facilities), Université de Poitiers, Faperj – Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro funding DM and OO (Professor International Mobility grants: E-26/203.635/2025 SEl-260003/008356/2025; E-26/203.636/2025 SEl-260003/012121/2025), and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES - financing code 001), PhD fellowship grant for EC.
Editor: Camila Cupello
References
Aguilera, O. and Marceniuk, A.P. 2018. Neogene tropical sea catfish (Siluriformes: Ariidae), with insights into paleo and modern diversity within northeastern South America. Journal of South American Earth Sciences 82: 108–121. Crossref
Aguilera, O., Lundberg, J., Birindelli, J., Sabaj Pérez, M., Jaramillo, C., and Sanchez-Villagra, M.R. 2013. Palaeontological evidence for the last temporal occurrence of the ancient Western Amazonian river outflow into the Caribbean. PLoS ONE 8(9): e76202. Crossref
Aquino, A.E. and Schaefer, S.A. 2002. The temporal region of the cranium of loricarioid catfishes (Teleostei: Siluriformes): morphological diversity and phylogenetic significance. Zoologischer Anzeiger 241: 223–244. Crossref
Arratia, G. 1987. Description of the primitive family Diplomystidae (Siluriformes, Teleostei, Pisces): morphology, taxonomy and phylogenetic implications. Bonner Zoologische Monographien 24: 1–120.
Arratia, G. 2003. Catfish head skeleton, an overview. In: G. Arratia, B.G. Kapoor, M. Chardon, and R. Diogo (eds.). Catfishes, 3–46. Science Publishers, Enfield.
Arratia, G. 2025. Siluriforms and the South American fossil record: their significance and the origin and evolutionary radiation of the group. In: G. Arratia and R.E. Reis (eds.). Catfishes, a Highly Diversified Group, Volume 2: Evolution and Phylogeny, 70–94. CRC Press, Boca Raton. Crossref
Arratia, G. and Gayet, M. 1995. Sensory canals and related bones of Tertiary siluriform crania from Bolivia and North America and comparison with recent forms. Journal of Vertebrate Paleontology 15: 482–505. Crossref
Azpelicueta, M.M. and Cione, A.L. 2011. Redescription of the Eocene catfish Bachmannia chubutensis (Teleostei: Bachmanniidae) of southern South America. Journal of Vertebrate Paleontology 31 258–269. Crossref
Azpelicueta, M.M. and Rubilar, A. 1998. A Miocene Nematogenys (Teleostei: Siluriformes: Nematogenyidae) from South-Central Chile. Journal of Vertebrate Paleontology 18: 475–483. Crossref
Azpelicueta, M.M., Cione, A.L., Cozzuol, M.A., and Mirande, J.M. 2015. Kooichthys jono n. gen. n. sp., a primitive catfish (Teleostei: Siluriformes) from the marine Miocene of southern South America. Journal of Paleontology 89: 791–801. Crossref
Birindelli, J.L.O. 2014. Phylogenetic relationships of the South American Doradoidea (Ostariophysi: Siluriformes). Neotropical Ichthyology 12: 451–564. Crossref
Bogan, S. and Agnolin, F.L. 2024. First fossil record of Aspredinidae: a new species from the late Miocene of northeastern Argentina. Zootaxa 5493: 392–400. Crossref
Bogan, S., Agnolin, F.L., and Scanferla, A. 2018. A new Andinichthyidae catfish (Ostariophysi: Siluriformes) from the Paleogene of northwestern Argentina. Journal of Vertebrate Paleontology 38: e1449117. Crossref
Calegari, B.B., Vari, R.P., and Reis, R.E. 2019. Phylogenetic systematics of the driftwood catfishes (Siluriformes: Auchenipteridae): a combined morphological and molecular analysis. Zoological Journal of the Linnean Society 187: 661–773. Crossref
Carvalho, T.P., Cardoso, A.R., Friel, J.P., and Reis, R.E. 2015. Two new species of the banjo catfish Bunocephalus Kner (Siluriformes: Aspredinidae) from the upper and middle Rio São Francisco basins, Brazil. Neotropical Ichthyology 13: 499–512. Crossref
Cockerell, T. 1925. A fossil fish of the family Callichthyidae. Science 62: 397–398. Crossref
DigiMorph 2025. Conorhynchos conirostris. Available at https://www.digimorph.org/specimens/Conorhynchos_conirostris/
Ferraris, C.J. 2007. Checklist of catfishes, recent and fossil (Osteichthyes: Siluriformes), and catalogue of siluriform primary types. Zootaxa 1418: 1–628. Crossref
Fricke, R., Eschmeyer, W.N., and Fong, J.D. 2021. Eschmeyer’s Catalog of Fishes: Genera/species by Family/subfamily. Available at http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp [accessed 30 May 2025].
Gayet, M. 1988. Le plus ancien crâne de Siluriforme: Andinichthys bolivianensis nov. gen., nov. sp. (Andinichthyidae nov. fam.) du Maastrichtien de Tiupampa (Bolivie). Comptes Rendus de l’Académie des Sciences, série 2 307: 833–836.
Gayet, M. 1990. Nouveaux Siluriformes du Maastrichtien de Tiupampa (Bolivie). Comptes Rendus de l’Académie des Sciences, série 2 310: 867–872.
Gayet, M. 1991. Holostean and teleostean fishes of Bolivia. Revista Técnica YPFB 12: 453–494.
Gayet, M. and Meunier, F. 1998. Maastrichtian to early late Paleocene freshwater Osteichthyes of Bolivia: additions and comments. In: L.R. Malabarba, R.E. Reis, R.P. Vari, Z.M. Lucena, and C.A.S. Lucena (eds.), Phylogeny and Classification of Neotropical Fishes, Part 1: Fossils and Geological Evidence, 85–110. Edipucrs, Porto Alegre.
Gayet, M., Marshall, and L.G., Sempéré, T. 1991. The Mesozoic and Paleocene vertebrates of Bolivia and their stratigraphic context: a review. Revista Técnica YPFB 12 (3–4): 393–433.
Gayet, M., Marshall, L.G., Sempere, T., Meunier, F.J., Cappetta, H., and Rage, J.C. 2001. Middle Maastrichtian vertebrates (fishes, amphibians, dinosaurs and other reptiles, mammals) from Pajcha Pata (Bolivia). Biostratigraphic, palaeoecologic and palaeobiogeographic implications. Palaeogeography, Palaeoclimatology, Palaeoecology 169: 39–68. Crossref
Gelfo, J.N., Goin, F.J., Woodburne, M.O., and Muizon, C. de 2009. Biochronological relationships of the earliest South American Paleogene mammalian faunas. Palaeontology 52: 251–269. Crossref
Horton, B.K. and DeCelles, P.G. 1997. The modern foreland basin system adjacent to the Central Andes. Geology 25: 895–898. Crossref
Huysentruyt, F. and Adriaens, D. 2005. Descriptive osteology of Corydoras aeneus (Siluriformes: Callichthyidae). Cybium 29 (3): 261–273.
Lundberg, J.G. 2005. Brachyplatystoma promagdalena n. sp., a fossil goliath catfish (Siluriformes: Pimelodidae) from the Miocene of Colombia, South America. Neotropical Ichthyology 3: 597–605. Crossref
Lundberg, J.G. and Aguilera, O. 2003. The late Miocene Phractocephalus catfish (Siluriformes: Pimelodidae) from Urumaco, Venezuela: additional specimens and reinterpretation as a distinct species. Neotropical Ichthyology 1: 97–109. Crossref
Lundberg, J.G., Marshall, L.G., Guerrero, J., Horton, B., Malabarba, M.C.S.L., Wesselingh, F. 1998. The stage for Neotropical fish diversification: a history of tropical South American rivers. In: L.R. Malabarba, R.E. Reis, R.P. Vari, Z.M. Lucena, and C.A.S. Lucena (eds.), Phylogeny and Classification of Neotropical Fishes, Part 1: Fossils and Geological Evidence, 14–48. Edipucrs, Porto Alegre.
Marceniuk, A.P. and Menezes, N.A. 2007. Systematics of the family Ariidae (Ostariophysi, Siluriformes), with a redefinition of the genera. Zootaxa 1416: 1–126. Crossref
Marceniuk, A.P., Acero, A.P., Cooke, R., and Betancur-R, R. 2017. Taxonomic revision of the New World genus Ariopsis Gill (Siluriformes: Ariidae), with description of two new species. Zootaxa 4290: 1–42. Crossref
Marceniuk, A.P., Menezes, N.A., and Britto, M.R. 2012. Phylogenetic analysis of the family Ariidae (Ostariophysi: Siluriformes), with a hypothesis on the monophyly and relationships of the genera. Zoological Journal of the Linnean Society 165: 534–669. Crossref
Marshall, L.G., Muizon, C. de, and Sigogneau-Russell, D. 1995. Pucadelphys andinus (Marsupialia, Mammalia) from the early Paleocene of Bolivia. Part I. The locality of Tiupampa: age, taphonomy and mammalian fauna. Mémoires du Muséum national d’histoire naturelle 165: 11–20.
Marshall, L.G., Sempéré, T., and Butler, R.F. 1997. Chronostratigraphy of the mammal-bearing Paleocene of South America. Journal of South American Earth Sciences 10: 49–70. Crossref
Muizon, C. de and Ladevèze, S. 2020. Cranial anatomy of Andinodelphys cochabambensis, a stem metatherian from the early Palaeocene of Bolivia. Geodiversitas 42 (30): 597–739. Crossref
Muizon, C. de, Gayet, M., Lavenu, A., Marshall, L.G., Sigé, B., and Villarroel, C. 1983. Late Cretaceous vertebrates, including mammals, from Tiupampa, South-central Bolivia. Geobios 16: 747–753. Crossref
Murray, A.M. and Holmes, R. 2021. Osteology of the cranium and Weberian apparatus of African catfish families (Teleostei: Ostariophysi: Siluriformes) with an assessment of Palaeogene genera. Vertebrate Anatomy Morphology Palaeontology 9 (1): 156–191. Crossref
Otero, O., Cavin, L., and Cespedes, R. 2025. Siluriformes from the Late Cretaceous–Paleocene of Bolivia. In: G. Arratia and R.E. Reis (eds.), Catfishes, a Highly Diversified Group, Volume 2: Evolution and Phylogeny, 52–69. CRC Press, Boca Raton. Crossref
Pinna, M. de, Peixoto, L.,
Tagliacollo, V., Britto, M. 2025. Phylogenetic relationships and
evolution of major groups of Siluriformes. In:
G. Arratia and R.E. Reis (eds.), Catfishes, a Highly Diversified Group,
Volume 2: Evolution and Phylogeny, 97–127. CRC
Press, Boca
Raton. Crossref
Scottese, C.R. 2001. Paleomap Project. Available at: http://www.scotese.com/.
Sempéré, T., Butler, R.F., Richards, D.R., Marshall, L.G., Sharp, W., and Swisher, C.C. III. 1997. Stratigraphy and chronology of Upper Cretaceous–lower Paleogene strata in Bolivia and northwest Argentina. GSA Bulletin 109: 709–727. Crossref
Sullivan, J.P., Muriel-Cunha, J., and Lundberg, J.G. 2013. Phylogenetic relationships and molecular dating of the major groups of catfishes of the Neotropical superfamily Pimelodoidea (Teleostei, Siluriformes). Proceedings of the Academy of Natural Sciences of Philadelphia 162 (1): 89–110. Crossref
Wiley, E.O., Johnson, G.D. 2010.
A teleost classification based on monophyletic groups. In:
J.S. Nelson, H.-P. Schultze, and M.V.H. Wilson (eds.), Origin
and Phylogenetic Interrelationships of Teleosts,
123–182. Verlag Dr. Friedrich Pfeil, München.
Acta Palaeontol. Pol. 71 (1): 109–118, 2026
http://doi.org/10.4202/app.01300.2025